
Porcine Astrovirus Infection in Brains of Pigs in Korea
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. RNA Extraction and cDNA Synthesis
2.3. PCR Screening and Sequencing
2.4. Sequence Analysis
2.5. Other Laboratory Diagnostics
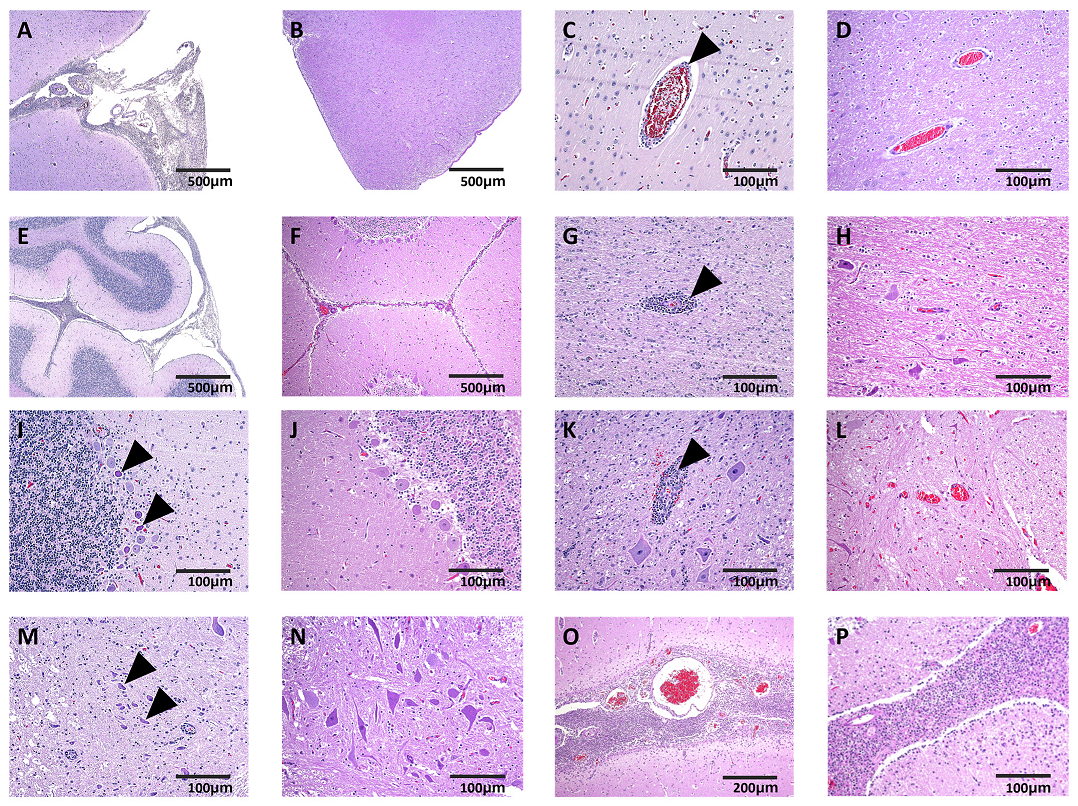
2.6. Histopathology
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guix, S.; Bosch, A.; Pintó, R.M. Astrovirus taxonomy. In Astrovirus Research; Springer: Berlin/Heidelberg, Germany, 2012; pp. 97–118. [Google Scholar]
- Bosch, A.; Pintó, R.M.; Guix, S. Human astroviruses. Clin. Microbiol. Rev. 2014, 27, 1048–1074. [Google Scholar] [CrossRef]
- Xiao, C.-T.; Gimenez-Lirola, L.G.; Gerber, P.F.; Jiang, Y.-H.; Halbur, P.G.; Opriessnig, T. Identification and characterization of novel porcine astroviruses (PAstVs) with high prevalence and frequent co-infection of individual pigs with multiple PAstV types. J. Gen. Virol. 2013, 94, 570–582. [Google Scholar] [CrossRef] [PubMed]
- Laurin, M.-A.; Dastor, M.; L’Homme, Y. Detection and genetic characterization of a novel pig astrovirus: Relationship to other astroviruses. Arch. Virol. 2011, 156, 2095. [Google Scholar] [CrossRef]
- Luo, Z.; Roi, S.; Dastor, M.; Gallice, E.; Laurin, M.-A.; L’Homme, Y. Multiple novel and prevalent astroviruses in pigs. Vet. Microbiol. 2011, 149, 316–323. [Google Scholar] [CrossRef]
- Donato, C.; Vijaykrishna, D. The broad host range and genetic diversity of mammalian and avian astroviruses. Viruses 2017, 9, 102. [Google Scholar] [CrossRef] [PubMed]
- Madeley, C.; Cosgrove, B. 28 nm particles in faeces in infantile gastroenteritis. Lancet 1975, 306, 451–452. [Google Scholar] [CrossRef]
- Bon, F.; Fascia, P.; Dauvergne, M.; Tenenbaum, D.; Planson, H.; Petion, A.; Pothier, P.; Kohli, E. Prevalence of group A rotavirus, human calicivirus, astrovirus, and adenovirus type 40 and 41 infections among children with acute gastroenteritis in Dijon, France. J. Clin. Microbiol. 1999, 37, 3055–3058. [Google Scholar] [CrossRef] [PubMed]
- Aktaş, O.; Aydin, H.; Timurkan, M.O. A molecular study on the prevalence and coinfections of Rotavirus, Norovirus, Astrovirus and Adenovirus in children with gastroenteritis. Minerva Pediatr. 2019, 71, 431–437. [Google Scholar] [CrossRef]
- Zhirakovskaia, E.; Tikunov, A.; Tymentsev, A.; Sokolov, S.; Sedelnikova, D.; Tikunova, N. Changing pattern of prevalence and genetic diversity of rotavirus, norovirus, astrovirus, and bocavirus associated with childhood diarrhea in Asian Russia, 2009–2012. Infect. Genet. Evol. 2019, 67, 167–182. [Google Scholar] [CrossRef]
- Xiao, C.T.; Halbur, P.G.; Opriessnig, T. Complete genome sequence of a newly identified porcine astrovirus genotype 3 strain US-MO123. J. Virol. 2012, 86, 13126. [Google Scholar] [CrossRef] [PubMed]
- De Benedictis, P.; Schultz-Cherry, S.; Burnham, A.; Cattoli, G. Astrovirus infections in humans and animals–molecular biology, genetic diversity, and interspecies transmissions. Infect. Genet. Evol. 2011, 11, 1529–1544. [Google Scholar] [CrossRef] [PubMed]
- Janowski, A.B. Beyond the Gastrointestinal Tract: The Emerging and Diverse Tissue Tropisms of Astroviruses. Viruses 2021, 13, 732. [Google Scholar] [CrossRef] [PubMed]
- Gough, R.; Collins, M.; Borland, E.; Keymer, L. Astrovirus-like particles associated with hepatitis in ducklings. Vet. Rec. 1984, 114, 279. [Google Scholar] [CrossRef]
- Imada, T.; Yamaguchi, S.; Mase, M.; Tsukamoto, K.; Kubo, M.; Morooka, A. Avian nephritis virus (ANV) as a new member of the family Astroviridae and construction of infectious ANV cDNA. J. Virol. 2000, 74, 8487–8493. [Google Scholar] [CrossRef] [PubMed]
- Quan, P.-L.; Wagner, T.A.; Briese, T.; Torgerson, T.R.; Hornig, M.; Tashmukhamedova, A.; Firth, C.; Palacios, G.; Baisre-De-Leon, A.; Paddock, C.D. Astrovirus encephalitis in boy with X-linked agammaglobulinemia. Emerg. Infect. Dis. 2010, 16, 918. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.R.; Morfopoulou, S.; Hubb, J.; Emmett, W.A.; Ip, W.; Shah, D.; Brooks, T.; Paine, S.M.; Anderson, G.; Virasami, A. Astrovirus VA1/HMO-C: An increasingly recognized neurotropic pathogen in immunocompromised patients. Clin. Infect. Dis. 2015, 60, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Ullman, K.; Chowdry, V.; Reining, M.; Benyeda, Z.; Baule, C.; Juremalm, M.; Wallgren, P.; Schwarz, L.; Zhou, E. Molecular investigations on the prevalence and viral load of enteric viruses in pigs from five European countries. Vet. Microbiol. 2016, 182, 75–81. [Google Scholar] [CrossRef]
- Blomström, A.-L.; Widén, F.; Hammer, A.-S.; Belák, S.; Berg, M. Detection of a novel astrovirus in brain tissue of mink suffering from shaking mink syndrome by use of viral metagenomics. J. Clin. Microbiol. 2010, 48, 4392–4396. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Diab, S.; McGraw, S.; Barr, B.; Traslavina, R.; Higgins, R.; Talbot, T.; Blanchard, P.; Rimoldi, G.; Fahsbender, E. Divergent astrovirus associated with neurologic disease in cattle. Emerg. Infect. Dis. 2013, 19, 1385. [Google Scholar] [CrossRef]
- Pfaff, F.; Schlottau, K.; Scholes, S.; Courtenay, A.; Hoffmann, B.; Höper, D.; Beer, M. A novel astrovirus associated with encephalitis and ganglionitis in domestic sheep. Transbound. Emerg. Dis. 2017, 64, 677–682. [Google Scholar] [CrossRef]
- Bailey Arruda, P.A.; Hensch, M.; Chen, Q.; Zheng, Y.; Yang, C.; Gatto, I.R.H.; Ferreyra, F.M.; Gauger, P.; Schwartz, K.; Bradner, L. Porcine astrovirus type 3 in central nervous system of swine with polioencephalomyelitis. Emerg. Infect. Dis. 2017, 23, 2097. [Google Scholar] [CrossRef] [PubMed]
- Boros, Á.; Albert, M.; Pankovics, P.; Bíró, H.; Pesavento, P.A.; Phan, T.G.; Delwart, E.; Reuter, G. Outbreaks of neuroinvasive astrovirus associated with encephalomyelitis, weakness, and paralysis among weaned pigs, Hungary. Emerg. Infect. Dis. 2017, 23, 1982. [Google Scholar] [CrossRef] [PubMed]
- Boujon, C.L.; Koch, M.C.; Kauer, R.V.; Keller-Gautschi, E.; Hierweger, M.M.; Hoby, S.; Seuberlich, T. Novel encephalomyelitis-associated astrovirus in a muskox (Ovibos moschatus): A surprise from the archives. Acta Vet. Scand. 2019, 61, 31. [Google Scholar] [CrossRef]
- Küchler, L.; Koch, M.C.; Seuberlich, T.; Boujon, C.L. Archive mining brings to light a 25-year old astrovirus encephalitis case in a sheep. Front. Vet. Sci. 2019, 6, 51. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-Y.; Kim, J.-H.; Kim, Y.-J.; Kim, Y.-S.; Roh, S.-G.; Lee, K.-H.; Kim, H.-J.; Shin, J.-H.; Oem, J.-K. Astrovirus infection in cattle with nonsuppurative meningoencephalitis in South Korea. Viruses 2021, 13, 1941. [Google Scholar] [CrossRef] [PubMed]
- Chae, S.B.; Jeong, C.G.; Park, J.S.; Na, E.J.; Oem, J.K. Detection and Genetic Characterization of Astroviruses in Brain Tissues of Wild Raccoon Dogs. Viruses 2023, 15, 1488. [Google Scholar] [CrossRef]
- Wildi, N.; Seuberlich, T. Neurotropic Astroviruses in Animals. Viruses 2021, 13, 1201. [Google Scholar] [CrossRef] [PubMed]
- Wohlgemuth, N.; Honce, R.; Schultz-Cherry, S. Astrovirus evolution and emergence. Infect. Genet. Evol. 2019, 69, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Blomström, A.-L.; Ley, C.; Jacobson, M. Astrovirus as a possible cause of congenital tremor type AII in piglets? Acta Vet. Scand. 2014, 56, 82. [Google Scholar] [CrossRef]
- Indik, S.; Valícek, L.; Smíd, B.; Dvoráková, H.; Rodák, L. Isolation and partial characterization of a novel porcine astrovirus. Vet. Microbiol. 2006, 117, 276–283. [Google Scholar] [CrossRef]
- Padmanabhan, A.; Hause, B.M. Detection and characterization of a novel genotype of porcine astrovirus 4 from nasal swabs from pigs with acute respiratory disease. Arch. Virol. 2016, 161, 2575–2579. [Google Scholar] [CrossRef]
- Opriessnig, T.; Xiao, C.-T.; Halbur, P.G. Porcine astrovirus type 5-associated enteritis in pigs. J. Comp. Pathol. 2020, 181, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Wang, C.; Liu, H.; Wu, Q.; Liang, S.; Cen, M.; Dong, Q.; Wei, Y.; Chen, Y.; Ouyang, K. Pathogenic characteristics of a porcine astrovirus strain isolated in China. Viruses 2019, 11, 1156. [Google Scholar] [CrossRef] [PubMed]
- Rawal, G.; Ferreyra, F.M.; Macedo, N.R.; Bradner, L.K.; Harmon, K.M.; Mueller, A.; Allison, G.; Linhares, D.C.; Arruda, B.L. Detection and cellular tropism of porcine astrovirus type 3 on breeding farms. Viruses 2019, 11, 1051. [Google Scholar] [CrossRef] [PubMed]
- Matias Ferreyra, F.S.; Bradner, L.K.; Burrough, E.R.; Cooper, V.L.; Derscheid, R.J.; Gauger, P.C.; Harmon, K.M.; Madson, D.; Piñeyro, P.E.; Schwartz, K.J. Polioencephalomyelitis in domestic swine associated with porcine astrovirus type 3. Vet. Pathol. 2020, 57, 82–89. [Google Scholar] [CrossRef]
- Ferreyra, F.M.; Arruda, P.E.; Bradner, L.K.; Harmon, K.M.; Zhang, M.; Giménez-Lirola, L.; Arruda, B.L. Experimental porcine astrovirus type 3-associated polioencephalomyelitis in swine. Vet. Pathol. 2021, 58, 1064–1074. [Google Scholar] [CrossRef]
- Lee, M.H.; Jeoung, H.Y.; Park, H.R.; Lim, J.A.; Song, J.Y.; An, D.J. Phylogenetic analysis of porcine astrovirus in domestic pigs and wild boars in South Korea. Virus Genes 2013, 46, 175–181. [Google Scholar] [CrossRef]
- Lee, S.; Jang, G.; Lee, C. Complete genome sequence of a porcine astrovirus from South Korea. Arch. Virol. 2015, 160, 1819–1821. [Google Scholar] [CrossRef]
- Shin, J.; Choe, S.; Hyun, B.H.; An, D.J. Phylogenetic analysis of kobuviruses and astroviruses from Korean wild boars: 2016–2018. Arch. Virol. 2021, 166, 2591–2596. [Google Scholar] [CrossRef]
- Chu, D.; Poon, L.; Guan, Y.; Peiris, J. Novel astroviruses in insectivorous bats. J. Virol. 2008, 82, 9107–9114. [Google Scholar] [CrossRef]
- Lee, S.-Y.; Son, K.-D.; Yong-Sik, K.; Wang, S.-J.; Kim, Y.-K.; Jheong, W.-H.; Oem, J.-K. Genetic diversity and phylogenetic analysis of newly discovered bat astroviruses in Korea. Arch. Virol. 2018, 163, 3065–3072. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.; Biosciences, I.; Carlsbad, C. BioEdit: An important software for molecular biology. GERF Bull Biosci 2011, 2, 60–61. [Google Scholar]
- Sievers, F.; Higgins, D.G. Clustal omega. Curr. Protoc. Bioinform. 2014, 48, 3.13.1–3.13.16. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef] [PubMed]
- Shiau, A.-L.; Lin, Y.-P.; Shieh, G.-S.; Su, C.-H.; Wu, W.-L.; Tsai, Y.-S.; Cheng, C.-W.; Lai, M.-D.; Wu, C.-L. Development of a conditionally replicating pseudorabies virus for HER-2/neu-overexpressing bladder cancer therapy. Mol. Ther. 2007, 15, 131–138. [Google Scholar] [CrossRef]
- Paton, D.J.; McGoldrick, A.; Greiser-Wilke, I.; Parchariyanon, S.; Song, J.-Y.; Liou, P.; Stadejek, T.; Lowings, J.; Björklund, H.; Belak, S. Genetic typing of classical swine fever virus. Vet. Microbiol. 2000, 73, 137–157. [Google Scholar]
- Xu, X.-G.; Chen, G.-D.; Huang, Y.; Ding, L.; Li, Z.-C.; Chang, C.-D.; Wang, C.-Y.; Tong, D.-W.; Liu, H.-J. Development of multiplex PCR for simultaneous detection of six swine DNA and RNA viruses. J. Virol. Methods 2012, 183, 69–74. [Google Scholar] [CrossRef]
- Henritzi, D.; Zhao, N.; Starick, E.; Simon, G.; Krog, J.S.; Larsen, L.E.; Reid, S.M.; Brown, I.H.; Chiapponi, C.; Foni, E. Rapid detection and subtyping of European swine influenza viruses in porcine clinical samples by haemagglutinin-and neuraminidase-specific tetra-and triplex real-time RT-PCR s. Influenza Other Respir. Viruses 2016, 10, 504–517. [Google Scholar]
- La, T.; Phillips, N.D.; Hampson, D.J. Development of a duplex PCR assay for detection of Brachyspira hyodysenteriae and Brachyspira pilosicoli in pig feces. J. Clin. Microbiol. 2003, 41, 3372–3375. [Google Scholar] [CrossRef]
- La, T.; Collins, A.; Phillips, N.; Oksa, A.; Hampson, D. Development of a multiplex-PCR for rapid detection of the enteric pathogens Lawsonia intracellularis, Brachyspira hyodysenteriae, and Brachyspira pilosicoli in porcine faeces. Lett. Appl. Microbiol. 2006, 42, 284–288. [Google Scholar] [CrossRef]
- Seo, B.; Moon, J.; Gi Jeong, W.; Chai Kim, S. Won-Il Kim and Hur J. Virulence-associated Genes and Antimicrobial Resistance of Escherichia coli Isolated From Post-weaning Piglets with Diarrhea in Korea. J. Bacteriol. Mycol. 2018, 5, 1090. [Google Scholar]
- Zhang, W.; Zhao, M.; Ruesch, L.; Omot, A.; Francis, D. Prevalence of virulence genes in Escherichia coli strains recently isolated from young pigs with diarrhea in the US. Vet. Microbiol. 2007, 123, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Angen, Ø.; Oliveira, S.; Ahrens, P.; Svensmark, B.; Leser, T.D. Development of an improved species specific PCR test for detection of Haemophilus parasuis. Vet. Microbiol. 2007, 119, 266–276. [Google Scholar] [CrossRef]
- Suh, D.K.; Song, J.C. Simultaneous detection of Lawsonia intracellularis, Brachyspira hyodysenteriae and Salmonella spp. in swine intestinal specimens by multiplex polymerase chain reaction. J. Vet. Sci. 2005, 6, 231–237. [Google Scholar] [CrossRef]
- Burall, L.S.; Simpson, A.C.; Datta, A.R. Evaluation of a serotyping scheme using a combination of an antibody-based serogrouping method and a multiplex PCR assay for identifying the major serotypes of Listeria monocytogenes. J. Food Prot. 2011, 74, 403–409. [Google Scholar] [CrossRef]
- Moore, M.; Feist, M. Real-time PCR method for Salmonella spp. targeting the stn gene. J. Appl. Microbiol. 2007, 102, 516–530. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.H.; Jacobson, K.A.; Rose, J.; Zeller, R. Hematoxylin and eosin staining of tissue and cell sections. Cold Spring Harb. Protoc. 2008, 2008, pdb. prot4986. [Google Scholar] [CrossRef] [PubMed]
- Reuter, G.; Pankovics, P.; Boros, Á. Nonsuppurative (aseptic) meningoencephalomyelitis associated with neurovirulent astrovirus infections in humans and animals. Clin. Microbiol. Rev. 2018, 31, e00040-18. [Google Scholar] [CrossRef]
- Ulloa, J.C.; Gutiérrez, M.F. Genomic analysis of two ORF2 segments of new porcine astrovirus isolates and their close relationship with human astroviruses. Can. J. Microbiol. 2010, 56, 569–577. [Google Scholar] [CrossRef]
- King, A.M. Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Dolores, G.-W.; Caroline, B.; Hans, H.D.; Englund, L.; Anne, S.H.; Hedlund, K.-O.; Carl Hård af, S.; Nilsson, K.; Nowotny, N.; Puurula, V. Investigations into shaking mink syndrome: An encephalomyelitis of unknown cause in farmed mink (Mustela vison) kits in Scandinavia. J. Vet. Diagn. Investig. 2004, 16, 305–312. [Google Scholar] [CrossRef]
- Bouzalas, I.G.; Wüthrich, D.; Walland, J.; Drögemüller, C.; Zurbriggen, A.; Vandevelde, M.; Oevermann, A.; Bruggmann, R.; Seuberlich, T. Neurotropic astrovirus in cattle with nonsuppurative encephalitis in Europe. J. Clin. Microbiol. 2014, 52, 3318–3324. [Google Scholar] [CrossRef] [PubMed]
- Küchler, L.; Rüfli, I.; Koch, M.C.; Hierweger, M.M.; Kauer, R.V.; Boujon, C.L.; Hilbe, M.; Oevermann, A.; Zanolari, P.; Seuberlich, T. Astrovirus-Associated Polioencephalomyelitis in an Alpaca. Viruses 2021, 13, 50. [Google Scholar] [CrossRef] [PubMed]
- Staats, J.; Feder, I.; Okwumabua, O.; Chengappa, M. Streptococcus suis: Past and present. Vet. Res. Commun. 1997, 21, 381–407. [Google Scholar] [CrossRef] [PubMed]
 ) KOR/0983/2020, (
) KOR/0983/2020, ( ) KOR/1006-2/2020 and (
) KOR/1006-2/2020 and ( ) KOR/1295/2020, while Ni-PAstV3 sequences of USA/IA/7023/2017 and NI-Brain/HUN strains are marked with (▶). (A) PAstVs with Ni-AstVs and AastVs; (B) PAstV1; (C) PAstV3; (D) PAstV4; (E) PAstV5.
) KOR/0983/2020, () KOR/1006-2/2020 and () KOR/1295/2020, while Ni-PAstV3 sequences of USA/IA/7023/2017 and NI-Brain/HUN strains are marked with (▶). (A) PAstVs with Ni-AstVs and AastVs; (B) PAstV1; (C) PAstV3; (D) PAstV4; (E) PAstV5.
) KOR/1295/2020, while Ni-PAstV3 sequences of USA/IA/7023/2017 and NI-Brain/HUN strains are marked with (▶). (A) PAstVs with Ni-AstVs and AastVs; (B) PAstV1; (C) PAstV3; (D) PAstV4; (E) PAstV5.
) KOR/0983/2020, () KOR/1006-2/2020 and () KOR/1295/2020, while Ni-PAstV3 sequences of USA/IA/7023/2017 and NI-Brain/HUN strains are marked with (▶). (A) PAstVs with Ni-AstVs and AastVs; (B) PAstV1; (C) PAstV3; (D) PAstV4; (E) PAstV5.


{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′→3′) | Target Region | Position | Position Reference (GenBank No.) | |
|---|---|---|---|---|---|
| AstV Pol [41] | F1: GARTTYGATTGGRCKCGKTAYGA | ORF1b (RdRp) | 3498–3916 | KF787112 | |
| F2: GARTTYGATTGGRCKAGGTAYGA | |||||
| F3: CGKTAYGATGGKACKATHCC | 3513–3916 | ||||
| F4: AGGTAYGATGGKACKATHCC | |||||
| R: GGYTTKACCCACATNCCRAA | |||||
| ORF2 | ORF2 F | F: CTSYATGGGAAACTCCT | ORF2 | 4065–6597 | KF787112 |
| s2m | R: CCCTCGATCCTACTCGG | ||||
| AP-dT17 | R: GGCCACGCGTCGACTAGTAC-Oligo(dT)17 | ||||
| AP | R: GCCACGCGTCGACTAGTAC | ||||
| Samples | Target | Sequence (Accession No.) | Length (CDS) | |||
|---|---|---|---|---|---|---|
| No. | Location | Organ | ORF1b (RdRp) | ORF2 (Capsid) | ||
| 20-0983 | Jeollabuk-do, jeongeup † | Brain | + | 5 | PAstV5 KOR/0983/2020 Brain1 (OP643779) | 2247 (2226) |
| PAstV5 KOR/0983/2020 Brain2 (OP643780) | 2265 (2208) | |||||
| 20-1006-2 | Gyeongsangnam-do, hamyang | Brain | + | 1, 4, 5 | PAstV1 KOR/1006-2/2020 Brain (OP643769) | 2496 (2313) |
| PAstV4 KOR/1006-2/2020 Brain (OP643774) | 3104 (2465) | |||||
| PAstV5 KOR/1006-2/2020 Brain (OP643781) | 2937 (2208) | |||||
| 20-1295 | Jeollabuk-do, jeongeup † | Brain | + | 3, 5 | PAstV3_KOR/1295/2020 Brain (OP643772) | 2788 (2298) |
| PAstV5_KOR/1295/2020 Brain (OP643782) | 2437 (2238) | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.-S.; Jeong, C.-G.; Chae, S.-B.; Yang, M.-S.; Oh, B.; Lee, S.-Y.; Oem, J.-K. Porcine Astrovirus Infection in Brains of Pigs in Korea. Viruses 2024, 16, 1372. https://doi.org/10.3390/v16091372
Park J-S, Jeong C-G, Chae S-B, Yang M-S, Oh B, Lee S-Y, Oem J-K. Porcine Astrovirus Infection in Brains of Pigs in Korea. Viruses. 2024; 16(9):1372. https://doi.org/10.3390/v16091372
Chicago/Turabian StylePark, Jun-Soo, Chang-Gi Jeong, Su-Beom Chae, Myeon-Sik Yang, Byungkwan Oh, Sook-Young Lee, and Jae-Ku Oem. 2024. "Porcine Astrovirus Infection in Brains of Pigs in Korea" Viruses 16, no. 9: 1372. https://doi.org/10.3390/v16091372