
Development of a Quadruplex RT-qPCR Assay for Rapid Detection and Differentiation of PRRSV-2 and Its Predominant Genetic Sublineages in China


Abstract
1. Introduction
2. Materials and Methods
2.1. Virus
2.2. Major Software Used
2.3. Primer and Probe Development
2.4. Synthesis of Standard Plasmids
2.5. Reaction Condition Optimization
2.6. Establishment of the Standard Curve
2.7. Sensitivity Test
2.8. Specificity Test
2.9. Repeatability Test
2.10. Concordance Analysis Between RT-qPCR Results and Sequencing Results
2.11. Clinical Application
2.12. Statistical Analysis
3. Results
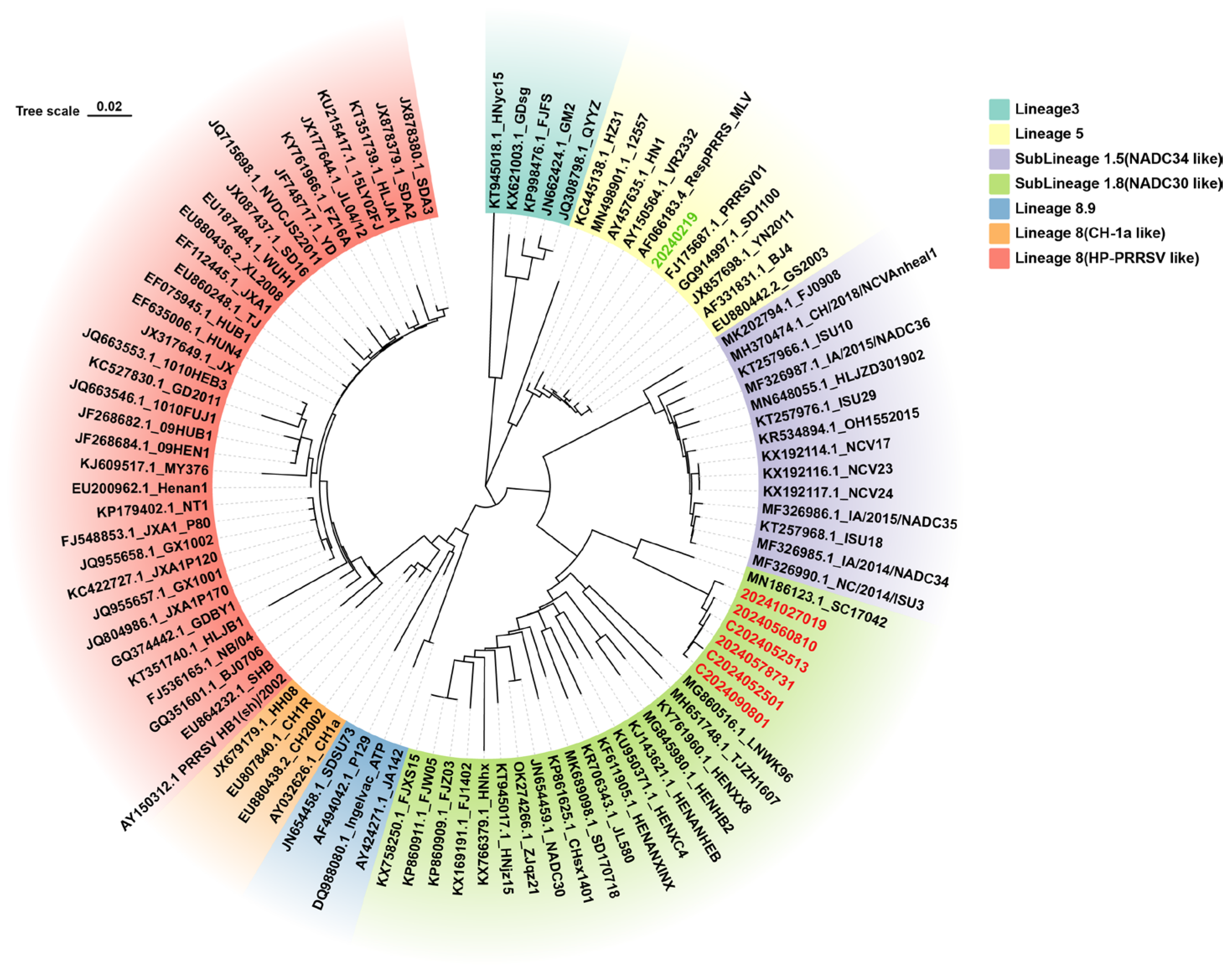
3.1. Primer and Probe Design
3.2. Optimization of Annealing Temperature
3.3. Optimization of Primer and Probe Concentration
3.4. Standard Curve Construction
3.5. Sensitivity Test
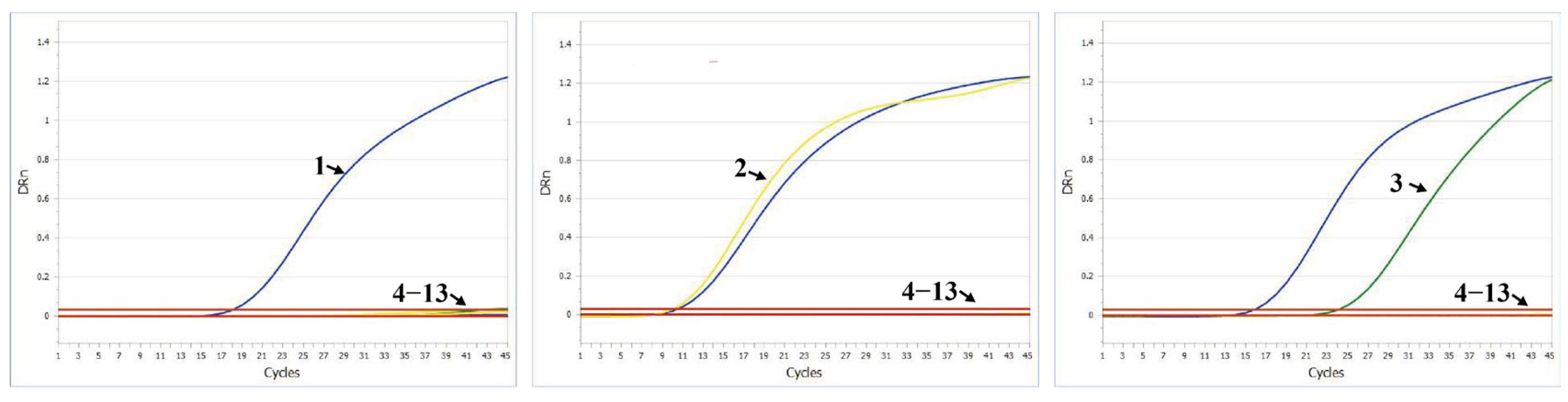
3.6. Specificity Test
3.7. Repeatability Test
3.8. Concordance Validation Study
3.8.1. Clinical Sample Testing from Farm A in Guangxi Zhuang Autonomous Region, China
3.8.2. Clinical Sample Testing from Farm B in Guangxi Zhuang Autonomous Region, China
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| PRRSV | Porcine reproductive and respiratory syndrome virus |
| C-PRRSV | Classic strains—Porcine reproductive and respiratory syndrome virus |
| HP-PRRSV | Highly pathogenic strains—Porcine reproductive and respiratory syndrome virus |
| PEDV | Porcine epidemic diarrhea virus |
| PoRV A | Group A porcine rotavirus |
| PDCoV | Porcine deltacoronavirus |
| PRV | Pseudorabies virus |
| CSFV | Classical swine fever virus |
| PPV | Porcine parvovirus |
| LOD | The limit of detection |
| CV | Coefficient of variation |
| aa | Amino acids |
References
- Collins, J.E.; Benfield, D.A.; Christianson, W.T.; Harris, L.; Hennings, J.C.; Shaw, D.P.; Goyal, S.M.; McCullough, S.; Morrison, R.B.; Joo, H.S. Isolation of Swine Infertility and Respiratory Syndrome Virus (Isolate ATCC VR-2332) in North America and Experimental Reproduction of the Disease in Gnotobiotic Pigs. J. Vet. Diagn. Investig. 1992, 4, 117–126. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, M.N.; Corzo, C.A.; Zimmerman, J.J.; Linhares, D.C.L. Longitudinal Piglet Sampling in Commercial Sow Farms Highlights the Challenge of PRRSV Detection. Porc. Health Manag. 2021, 7, 31. [Google Scholar] [CrossRef]
- Karniychuk, U.U.; Nauwynck, H.J. Quantitative Changes of Sialoadhesin and CD163 Positive Macrophages in the Implantation Sites and Organs of Porcine Embryos/Fetuses During Gestation. Placenta 2009, 30, 497–500. [Google Scholar] [CrossRef]
- Huang, B.; Li, F.; You, D.; Deng, L.; Xu, T.; Lai, S.; Ai, Y.; Huang, J.; Zhou, Y.; Ge, L.; et al. Porcine Reproductive and Respiratory Syndrome Virus Infects the Reproductive System of Male Piglets and Impairs Development of the Blood–Testis Barrier. Virulence 2024, 15, 2384564. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Zhang, H.; Cheng, H.; Liu, M.; Wen, S.; Ren, J. Progress in PRRSV Infection and Adaptive Immune Response Mechanisms. Viruses 2023, 15, 1442. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, P.; Xie, C.; Ha, Z.; Shi, N.; Zhang, H.; Li, Z.; Han, J.; Xie, Y.; Qiu, X.; et al. Synergistic Pathogenicity by Coinfection and Sequential Infection with NADC30-like PRRSV and PCV2 in Post-Weaned Pigs. Viruses 2022, 14, 193. [Google Scholar] [CrossRef] [PubMed]
- Holtkamp, D.J.; Kliebenstein, J.B.; Zimmerman, J.J.; Neumann, E.; Rotto, H.; Yoder, T.K.; Wang, C.; Yeske, P.; Mowrer, C.L.; Haley, C. Economic Impact of Porcine Reproductive and Respiratory Syndrome Virus on U.S. Pork Producers; Iowa State University: Ames, IA, USA, 2012. [Google Scholar]
- Zhang, Z.; Li, Z.; Li, H.; Yang, S.; Ren, F.; Bian, T.; Sun, L.; Zhou, B.; Zhou, L.; Qu, X. The Economic Impact of Porcine Reproductive and Respiratory Syndrome Outbreak in Four Chinese Farms: Based on Cost and Revenue Analysis. Front. Vet. Sci. 2022, 9, 1024720. [Google Scholar] [CrossRef]
- Tian, K.; Yu, X.; Zhao, T.; Feng, Y.; Cao, Z.; Wang, C.; Hu, Y.; Chen, X.; Hu, D.; Tian, X.; et al. Emergence of Fatal PRRSV Variants: Unparalleled Outbreaks of Atypical PRRS in China and Molecular Dissection of the Unique Hallmark. PLoS ONE 2007, 2, e526. [Google Scholar] [CrossRef]
- Brockmeier, S.L.; Loving, C.L.; Vorwald, A.C.; Kehrli, M.E.; Baker, R.B.; Nicholson, T.L.; Lager, K.M.; Miller, L.C.; Faaberg, K.S. Genomic Sequence and Virulence Comparison of Four Type 2 Porcine Reproductive and Respiratory Syndrome Virus Strains. Virus Res. 2012, 169, 212–221. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, Z.; Ding, Y.; Ge, X.; Guo, X.; Yang, H. NADC30-like Strain of Porcine Reproductive and Respiratory Syndrome Virus, China. Emerg. Infect. Dis. J. CDC 2015, 21, 12. [Google Scholar] [CrossRef]
- Zhou, L.; Yang, B.; Xu, L.; Jin, H.; Ge, X.; Guo, X.; Han, J.; Yang, H. Efficacy Evaluation of Three Modified-Live Virus Vaccines against a Strain of Porcine Reproductive and Respiratory Syndrome Virus NADC30-Like. Vet. Microbiol. 2017, 207, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Van Geelen, A.G.M.; Anderson, T.K.; Lager, K.M.; Das, P.B.; Otis, N.J.; Montiel, N.A.; Miller, L.C.; Kulshreshtha, V.; Buckley, A.C.; Brockmeier, S.L.; et al. Porcine Reproductive and Respiratory Disease Virus: Evolution and Recombination Yields Distinct ORF5 RFLP 1-7-4 Viruses with Individual Pathogenicity. Virology 2018, 513, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-L.; Zhang, W.-L.; Xiang, L.-R.; Leng, C.-L.; Tian, Z.-J.; Tang, Y.-D.; Cai, X.-H. Emergence of Novel Porcine Reproductive and Respiratory Syndrome Viruses (ORF5 RFLP 1-7-4 Viruses) in China. Vet. Microbiol. 2018, 222, 105–108. [Google Scholar] [CrossRef]
- Xie, C.-Z.; Ha, Z.; Zhang, H.; Zhang, Y.; Xie, Y.-B.; Zhang, H.; Nan, F.-L.; Wang, Z.; Zhang, P.; Xu, W.; et al. Pathogenicity of Porcine Reproductive and Respiratory Syndrome Virus (ORF5 RFLP 1-7-4 Viruses) in China. Transbound. Emerg. Dis. 2020, 67, 2065–2072. [Google Scholar] [CrossRef] [PubMed]
- Gong, B.; Xu, H.; Sun, Q.; Li, C.; Xiang, L.; Zhao, J.; Li, W.; Guo, Z.; Li, J.; Wang, Q.; et al. Dissecting Genetic Diversity and Evolutionary Trends of Chinese PRRSV-1 Based on Whole-Genome Analysis. Transbound. Emerg. Dis. 2024, 2024, 9705539. [Google Scholar] [CrossRef]
- Ming, S.; Yongying, M.; Bohua, L.; Huiying, L.; Xiaoyu, D.; Qiaorong, L.; Mingming, Q.; Xi, C.; Xinyan, Y.; Xizhao, C. Pathogenic Characterization of European Genotype Porcine Reproductive and Respiratory Syndrome Virus Recently Isolated in Mainland China. Open Virol. J. 2017, 11, 83–89. [Google Scholar] [CrossRef]
- Hsueh, F.-C.; Kuo, K.-L.; Hsu, F.-Y.; Wang, S.-Y.; Chiu, H.-J.; Wu, M.-T.; Lin, C.-F.; Huang, Y.-H.; Chiou, M.-T.; Lin, C.-N. Molecular Characteristics and Pathogenicity of Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) 1 in Taiwan during 2019–2020. Life 2023, 13, 843. [Google Scholar] [CrossRef]
- Wang, X.; Bai, X.; Wang, Y.; Wang, L.; Wei, L.; Tan, F.; Zhou, Z.; Tian, K. Pathogenicity Characterization of PRRSV-1 181187-2 Isolated in China. Microb. Pathog. 2023, 180, 106158. [Google Scholar] [CrossRef]
- Xu, H.; Gong, B.; Sun, Q.; Li, C.; Zhao, J.; Xiang, L.; Li, W.; Guo, Z.; Tang, Y.; Leng, C.; et al. Genomic Characterization and Pathogenicity of BJEU06-1-Like PRRSV-1 ZD-1 Isolated in China. Transbound. Emerg. Dis. 2023, 2023, 6793604. [Google Scholar] [CrossRef]
- Yang, S.; Cui, M.; Li, C.; Qiu, M.; Zhu, X.; Lin, Y.; Meng, Y.; Qiu, Y.; Qi, W.; Lin, H.; et al. Isolation and Genomic Characterization of a Novel Porcine Reproductive and Respiratory Syndrome Virus 1 from Severely Diseased Piglets in China in 2024. Vet. Sci. 2025, 12, 61. [Google Scholar] [CrossRef]
- Li, Y.; Ji, G.; Xu, X.; Wang, J.; Li, Y.; Tan, F.; Li, X. Development and Application of an RT-PCR to Differentiate the Prevalent NA-PRRSV Strains in China. Open Virol. J. 2017, 11, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Ruan, S.; Ren, W.; Yu, B.; Yu, X.; Wu, H.; Li, W.; Jiang, Y.; He, Q. Development and Implementation of a Quadruple RT-qPCR Method for the Identification of Porcine Reproductive and Respiratory Syndrome Virus Strains. Viruses 2023, 15, 1946. [Google Scholar] [CrossRef]
- Park, J.-Y.; Park, S.; Park, Y.-R.; Kang, D.-Y.; Kim, E.-M.; Jeon, H.-S.; Kim, J.-J.; Kim, W.-I.; Lee, K.-T.; Kim, S.-H.; et al. Reverse-Transcription Loop-Mediated Isothermal Amplification (RT-LAMP) Assay for the Visual Detection of European and North American Porcine Reproductive and Respiratory Syndrome Viruses. J. Virol. Methods 2016, 237, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Qin, X.; Sun, Y.; Chen, T.; Zhang, Z. Rapid Detection of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus by a Fluorescent Probe-Based Isothermal Recombinase Polymerase Amplification Assay. Virus Genes 2016, 52, 883–886. [Google Scholar] [CrossRef]
- Long, F.; Chen, Y.; Shi, K.; Yin, Y.; Feng, S.; Si, H. Development of a Multiplex Crystal Digital RT-PCR for Differential Detection of Classical, Highly Pathogenic, and NADC30-like Porcine Reproductive and Respiratory Syndrome Virus. Animals 2023, 13, 594. [Google Scholar] [CrossRef]
- Liu, S.; Tao, D.; Liao, Y.; Yang, Y.; Sun, S.; Zhao, Y.; Yang, P.; Tang, Y.; Chen, B.; Liu, Y.; et al. Highly Sensitive CRISPR/Cas12a-Based Fluorescence Detection of Porcine Reproductive and Respiratory Syndrome Virus. ACS Synth. Biol. 2021, 10, 2499–2507. [Google Scholar] [CrossRef]
- Wang, X.; Wang, G.; Wang, N.; Liu, J.; Cai, Y.; Ren, M.; Li, Z. A Simple and Efficient Method for the Generation of a Porcine Alveolar Macrophage Cell Line for High-Efficiency Porcine Reproductive and Respiratory Syndrome Virus 2 Infection. J. Virol. Methods 2019, 274, 113727. [Google Scholar] [CrossRef] [PubMed]
- Singh, C.; Roy-Chowdhuri, S. Quantitative Real-Time PCR: Recent Advances. In Clinical Applications of PCR; Luthra, R., Singh, R.R., Patel, K.P., Eds.; Springer: New York, NY, USA, 2016; pp. 161–176. ISBN 978-1-4939-3360-0. [Google Scholar]
- Xie, J.; Chen, Y.; Cai, G.; Cai, R.; Hu, Z.; Wang, H. Tree Visualization By One Table (tvBOT): A Web Application for Visualizing, Modifying and Annotating Phylogenetic Trees. Nucleic Acids Res. 2023, 51, W587–W592. [Google Scholar] [CrossRef]
- GB/T 18090-2023; Diagnostic Techniques for Porcine Reproductive and Respiratory Syndrome. Standards Press of China: Beijing, China, 2023.
- Hu, R.; Zhang, T.; Lai, R.; Ding, Z.; Zhuang, Y.; Liu, H.; Cao, H.; Gao, X.; Luo, J.; Chen, Z.; et al. PRRSV Elimination in a Farrow-to-Finish Pig Herd Using Herd Closure and Rollover Approach. Viruses 2023, 15, 1239. [Google Scholar] [CrossRef]
- Yim-im, W.; Anderson, T.K.; Paploski, I.A.D.; VanderWaal, K.; Gauger, P.; Krueger, K.; Shi, M.; Main, R.; Zhang, J. Refining PRRSV-2 Genetic Classification Based on Global ORF5 Sequences and Investigation of Their Geographic Distributions and Temporal Changes. Microbiol. Spectr. 2023, 11, e02916-23. [Google Scholar] [CrossRef]
- Zhao, Y.-Y.; Ma, X.; Chen, X.-M.; Song, Y.-P.; Zheng, L.-L.; Ma, S.-J.; Chen, H.-Y. Molecular Detection and Genetic Characteristics of Porcine Reproductive and Respiratory Syndrome Virus in Central China. Microb. Pathog. 2024, 197, 107024. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Ye, M.; Xiao, Y.; Li, S.; Huang, Y.; Li, X.; Tian, K.; Zhu, J. Development of Universal and Quadruplex Real-time RT-PCR Assays for Simultaneous Detection and Differentiation of Porcine Reproductive and Respiratory Syndrome Viruses. Transbound Emerg. Dis. 2019, 66, 2271–2278. [Google Scholar] [CrossRef] [PubMed]
- Tu, T.; Pang, M.; Jiang, D.; Zhou, Y.; Wu, X.; Yao, X.; Luo, Y.; Yang, Z.; Ren, M.; Lu, A.; et al. Development of a Real-Time TaqMan RT-PCR Assay for the Detection of NADC34-like Porcine Reproductive and Respiratory Syndrome Virus. Vet. Sci. 2023, 10, 279. [Google Scholar] [CrossRef]
- Fang, K.; Liu, S.; Li, X.; Chen, H.; Qian, P. Epidemiological and Genetic Characteristics of Porcine Reproductive and Respiratory Syndrome Virus in South China Between 2017 and 2021. Front. Vet. Sci. 2022, 9, 853044. [Google Scholar] [CrossRef]
- Zhang, R.; Li, H.; Xie, H.; Hou, X.; Zhou, L.; Cao, A.; Zeshan, B.; Zhou, Y.; Wang, X. Comparing the Molecular Evolution and Recombination Patterns of Predominant PRRSV-2 Lineages Co-Circulating in China. Front. Microbiol. 2024, 15, 1398470. [Google Scholar] [CrossRef]
- Zhang, H.; Luo, Q.; Zheng, Y.; Sha, H.; Li, G.; Kong, W.; Huang, L.; Zhao, M. Genetic Variability and Recombination of the NSP2 Gene of PRRSV-2 Strains in China from 1996 to 2021. Vet. Sci. 2023, 10, 325. [Google Scholar] [CrossRef]
- Liu, B.; Luo, L.; Shi, Z.; Ju, H.; Yu, L.; Li, G.; Cui, J. Research Progress of Porcine Reproductive and Respiratory Syndrome Virus NSP2 Protein. Viruses 2023, 15, 2310. [Google Scholar] [CrossRef] [PubMed]
- Otake, S.; Yoshida, M.; Dee, S. A Review of Swine Breeding Herd Biosecurity in the United States to Prevent Virus Entry Using Porcine Reproductive and Respiratory Syndrome Virus as a Model Pathogen. Animals 2024, 14, 2694. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Wu, H.; Wang, X. Effects of Herd Closure and Medication Programs on the Infection of NADC30-like PRRSV in Pig Farms. J. Vet. Sci. 2024, 25, e82. [Google Scholar] [CrossRef]
- Magalhães, E.S.; Zimmerman, J.J.; Holtkamp, D.J.; Classen, D.M.; Groth, D.D.; Glowzenski, L.; Philips, R.; Silva, G.S.; Linhares, D.C.L. Next Generation of Voluntary PRRS Virus Regional Control Programs. Front. Vet. Sci. 2021, 8, 769312. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Primer/Probe Sets | Sequences (5′-3′) | Locations (nt) | Amplicon Size |
|---|---|---|---|---|
| PRRSV 2 (ORF7/3′UTR) | PRRSV-2-F | GCACTGATTGACAYTGTGCC | 15,317–15,336 | 85 bp |
| PRRSV-2-R | CGCATGGTTCTCGCCAAT | 15,384–15,401 | ||
| PRRSV-2-P | AGTCACCTATTCAATTAGGGCGACCG | 15,341–15,366 | ||
| NADC30-like PRRSV (NSP2) | NADC30-like-F | AGCGTRYTGGAYACCTCCTTTG | 2178–2199 | 212 bp |
| NADC30-like-R | CTGATYTTYCTGCGYGGRG | 2371–2389 | ||
| NADC30-like-P | TTCTCTGGRAGGCTCCTGACAGACYC | 2331–2356 | ||
| HP-like PRRSV (NSP2) | HP-PRRSV-F | TGGGTCGGCRCCAKTTCCT | 2892–2910 | 101 bp |
| HP-PRRSV-R | YATATTCCGTYTGTGAGGACRC | 2916–2941 | ||
| HP-PRRSV-P | TCAGCGTTGTTGTYACAGTYCTRCGC | 2971–2992 | ||
| NADC34-like PRRSV (NSP2) | NADC34-like-F | AGTYCACCTAACYGAGTTRCC | 2231–2251 | 169 bp |
| 1-4-4-F | AGTTCACYYAACCAAGCTGCC | 2189–2209 | ||
| NADC34-like-R | AGGGRYGGGCTTGCAATCT | 2381–2399 | ||
| NADC34-like-P | CGGCCYCATCGCCRCCTCT | 2314–2332 |
| Target | Testicular Fluid | Serum | Tissue | Oral and Nasal Swabs | Total | Positive Rate (%) |
|---|---|---|---|---|---|---|
| PRRSV-2 | 28 | 47 | 4 | 1 | 80 | 21.50 |
| NADC30-like PRRSV | 17 | 29 | 4 | 0 | 50 | 13.44 |
| HP-like PRRSV | 0 | 0 | 0 | 0 | 0 | 0 |
| NADC34-like PRRSV | 0 | 0 | 0 | 0 | 0 | 0 |
| Total | 59 | 240 | 45 | 28 | 372 | - |
| Target | Testicular Fluid | Serum | Tissue | Oral and Nasal Swabs | Total | Positive Rate(%) |
|---|---|---|---|---|---|---|
| PRRSV-2 | 26 | 68 | 21 | 9 | 124 | 21.90 |
| NADC30-like PRRSV | 0 | 20 | 0 | 0 | 20 | 3.53 |
| HP-like PRRSV | 0 | 0 | 0 | 0 | 0 | 0 |
| NADC34-like PRRSV | 0 | 0 | 0 | 0 | 0 | 0 |
| Total | 109 | 352 | 45 | 60 | 566 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, G.; Xiong, S.; Su, Z.; Chen, G.; Liu, S.; Wang, Z.; Chen, H.; Zhang, A. Development of a Quadruplex RT-qPCR Assay for Rapid Detection and Differentiation of PRRSV-2 and Its Predominant Genetic Sublineages in China. Viruses 2025, 17, 853. https://doi.org/10.3390/v17060853
Ye G, Xiong S, Su Z, Chen G, Liu S, Wang Z, Chen H, Zhang A. Development of a Quadruplex RT-qPCR Assay for Rapid Detection and Differentiation of PRRSV-2 and Its Predominant Genetic Sublineages in China. Viruses. 2025; 17(6):853. https://doi.org/10.3390/v17060853
Chicago/Turabian StyleYe, Guishan, Siyu Xiong, Zhipeng Su, Guosheng Chen, Siyuan Liu, Zixuan Wang, Huanchun Chen, and Anding Zhang. 2025. "Development of a Quadruplex RT-qPCR Assay for Rapid Detection and Differentiation of PRRSV-2 and Its Predominant Genetic Sublineages in China" Viruses 17, no. 6: 853. https://doi.org/10.3390/v17060853
APA StyleYe, G., Xiong, S., Su, Z., Chen, G., Liu, S., Wang, Z., Chen, H., & Zhang, A. (2025). Development of a Quadruplex RT-qPCR Assay for Rapid Detection and Differentiation of PRRSV-2 and Its Predominant Genetic Sublineages in China. Viruses, 17(6), 853. https://doi.org/10.3390/v17060853