Roles of ITPA and IL28B Genotypes in Chronic Hepatitis C Patients Treated with Peginterferon Plus Ribavirin


 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Patient Characteristics According to IL28B and ITPA Genotypes

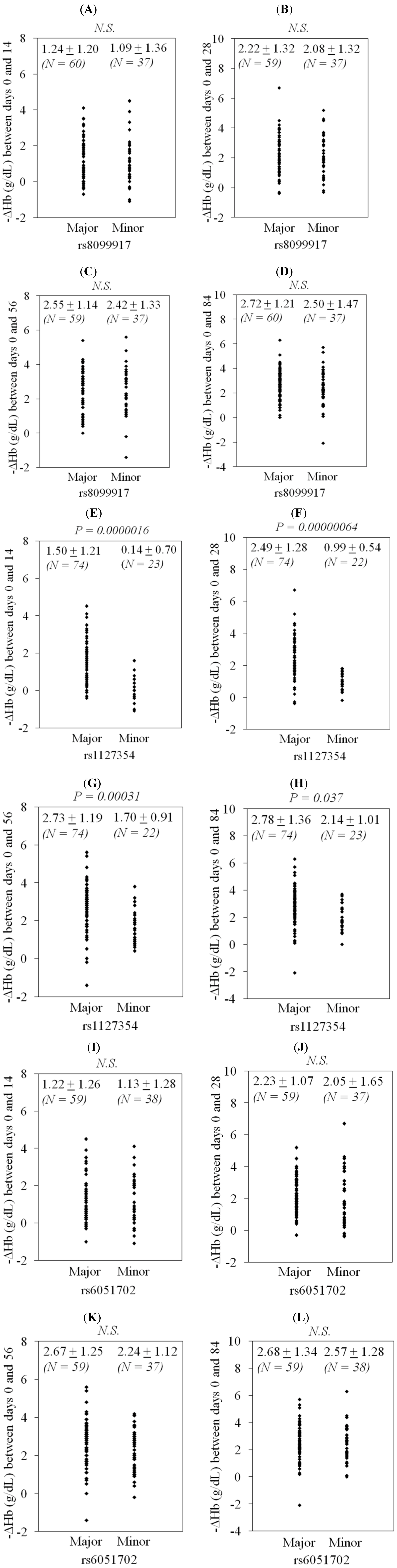{kind=link}
| Study variables | Total (n = 97) |
|---|---|
| Age (years) | 55.1 ± 10.8 |
| Gender (male/female) | 44/53 |
| SNP genotype | |
| IL28B rs8099917 TT/TG/GG | 60/35/2 |
| ITPA rs1127354 CC/CA/AA | 74/21/2 |
| ITPA rs6051702 AA/AC/CC | 59/32/6 |
| Response to previous therapy | |
| Naïve/relapse/null response | 67/17/13 |
| HCV RNA (H/L) | 95/2 |
| HCV genotype (G1/G2) | 81/16 |
| AST (IU/L) | 56.0 ± 49.4 |
| ALT (IU/L) | 67.9 ± 62.4 |
| γGTP (IU/L) | 53.5 ± 73.2 |
| WBC (/mm3) | 5,410 ± 1,640 |
| Hemoglobin (g/dL) | 14.0 ± 1.1 |
| Platelets (×104/mm3) | 17.5 ± 5.1 |
| History of diabetes mellitus (+/−) | 15/82 |
| US (CLD/cirrhosis/unknown) | 83/12/2 |
| Treatment Response | |
| RVR (+/−/unknown) | 14/82/1 |
| EVR (+/−) | 52/45 |
| SVR (+/relapser/null/unknown) | 40/27/22/8 |
2.2. Treatment Response According to IL28B and ITPA Genotypes
| Study variables | IL28B rs8099917 | ITPA rs1127354 | ITPA rs6051702 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| TT | TG/GG | P-value | CC | CA/AA | P-value | AA | AC/CC | P-value | |
| No. of patients | 60 | 37 | 74 | 23 | 59 | 38 | |||
| Age (years) | 55.7 ± 11.2 | 54.7 ± 10.1 | N.S. | 56.8 ± 9.7 | 49.6 ± 12.2 | 0.0043 | 55.6 ± 11.3 | 54.4 ± 9.9 | N.S. |
| Gender (male/female) | 25/35 | 19/18 | N.S. | 29/45 | 15/8 | 0.0511 | 29/30 | 15/23 | N.S. |
| Response to previous therapy (naïve/relapse/null response) | 46/10/4 | 21/7/9 | 0.029 | 48/17/9 | 19/0/4 | N.S. | 40/12/7 | 27/5/6 | N.S. |
| HCV RNA (H/L) | 58/2 | 37/0 | N.S. | 73/1 | 22/1 | N.S. | 58/1 | 37/1 | N.S. |
| HCV genotype (G1/G2) | 49/11 | 32/5 | N.S. | 63/11 | 18/5 | N.S. | 48/11 | 33/5 | N.S. |
| AST (IU/L) | 53.3 ± 56.2 | 60.3 ± 36.0 | N.S. | 52.8 ± 31.9 | 66.2 ± 84.4 | N.S. | 51.6 ± 30.1 | 62.8 ± 69.5 | N.S. |
| ALT (IU/L) | 62.4 ± 65.3 | 76.9 ± 57.0 | N.S. | 62.3 ± 48.5 | 85.7 ± 93.5 | N.S. | 62.4 ± 47.5 | 76.4 ± 80.2 | N.S. |
| γGTP (IU/L) | 35.5 ± 34.5 | 82.8 ± 104 | 0.0016 | 55.1 ± 80.9 | 48.7 ± 40.1 | N.S. | 51.6 ± 72.1 | 56.5 ± 75.6 | N.S. |
| WBC (/mm3) | 5580 ± 1820 | 5140 ± 1260 | N.S. | 5390 ± 1630 | 5470 ± 1680 | N.S. | 5570 ± 1690 | 5160 ± 1540 | N.S. |
| Hb (g/dL) | 13.9 ± 1.1 | 14.3 ± 1.1 | N.S. | 13.9 ± 1.0 | 14.3 ± 1.2 | N.S. | 14.0 ± 1.1 | 14.0 ± 1.1 | N.S. |
| Platelets (×104/mm3) | 17.9 ± 5.3 | 16.8 ± 5.0 | N.S. | 17.4 ± 5.4 | 17.7 ± 4.3 | N.S. | 17.8 ± 5.4 | 17.0 ± 4.7 | N.S. |
| History of diabetes mellitus (+/−) | 9/51 | 7/30 | N.S. | 11/63 | 4/19 | N.S. | 8/51 | 7/31 | N.S. |
| US (CLD/cirrhosis/unknown) | 51/8/1 | 32/4/1 | N.S. | 62/10/2 | 21/2 | N.S. | 50/8/1 | 33/4/1 | N.S. |
| Study variables | IL28B rs8099917 | ITPA rs1127354 | ITPA rs6051702 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| TT | TG/GG | P-value | CC | CA/AA | P-value | AA | AC/CC | P-value | |
| No. of patients | 60 | 37 | 74 | 23 | 59 | 38 | |||
| RVR (+/−/unknown) | 12/47/1 | 2/35/0 | 0.085 | 10/63/1 | 4/19/0 | N.S. | 10/49/0 | 4/33/1 | N.S. |
| EVR (+/−) | 43/17 | 9/28 | 0.000014 | 36/38 | 16/17 | N.S. | 31/28 | 21/17 | N.S. |
| SVR (+/Relapser/Null/unknown) | 29/6/18/7 | 11/16/9/1 | 0.042 | 28/17/22/7 | 12/5/5/1 | N.S. | 27/13/16/3 | 13/9/11/5 | N.S. |
2.3. Ribavirin-Induced Anemia According to IL28B and ITPA Genotypes

2.4. Association Between ITPA rs1127354 Genotype and Dose Reduction of Drugs During Treatment
| Study variables | ITPA rs1127354 major type | ITPA rs1127354 minor type |
|---|---|---|
| A | ||
| Dose reduction (+) | 17 | 4 |
| Dose reduction (−) | 57 | 19 |
| B | ||
| Dose reduction (+) | 22 | 0 |
| Dose reduction (−) | 52 | 23 |
2.5. Effects of IL28B and ITPA Genotypes on the Reduction of White Blood Cell/Neutrophil Count
2.6. Effects of IL28B and ITPA Genotypes on the Reduction of Platelet Count
3. Experimental Section
3.1. Patients
3.2. Treatment
3.3. HCV RNA Quantification
3.4. HCV Genotyping
3.5. Classification of Treatment Outcome
3.6. DNA Extraction and TaqMan SNP Assay
3.7. Statistical Analysis
4. Discussion and Conclusion
Acknowledgments
Conflict of Interest
References and Notes
- Di Bisceglie, A.M. Hepatitis C and hepatocellular carcinoma. Hepatology 1997, 26, 34S–38S. [Google Scholar] [CrossRef]
- Kanda, T.; Imazeki, F.; Yokosuka, O. New antiviral therapies for chronic hepatitis C. Hepatol. Int. 2010, 4, 548–561. [Google Scholar] [CrossRef]
- Jensen, D. A new era of hepatitis C therapy begins. N. Engl. J. Med. 2011, 364, 1272–1274. [Google Scholar] [CrossRef]
- Suppiah, V.; Moldovan, M.; Ahlenstiel, G.; Berg, T.; Weltman, M.; Abate, M.L.; Bassendine, M.; Spengler, U.; Dore, G.J.; Powell, E.; et al. IL28B is associated with response to chronic hepatitis C interferon-alpha and ribavirin therapy. Nat. Genet. 2009, 41, 1100–1104. [Google Scholar] [CrossRef]
- Tanaka, Y.; Nishida, N.; Sugiyama, M.; Kurosaki, M.; Matsuura, K.; Sakamoto, N.; Nakagawa, M.; Korenaga, M.; Hino, K.; Hige, S.; et al. Genome-wide association of IL28B with response to pegylated interferon-alpha and ribavirin therapy for chronic hepatitis C. Nat. Genet. 2009, 41, 1105–1109. [Google Scholar] [CrossRef]
- Ge, D.; Fellay, J.; Thompson, A.J.; Simon, J.S.; Shianna, K.V.; Urban, T.J.; Heinzen, E.L.; Qiu, P.; Bertelsen, A.H.; Muir, A.J.; et al. Genetic variation in IL28B predicts hepatitis C treatment-induced viral clearance. Nature 2009, 461, 399–401. [Google Scholar]
- Nakamoto, S.; Kanda, T.; Imazeki, F.; Wu, S.; Arai, M.; Fujiwara, K.; Yokosuka, O. Simple assay based on restriction fragment length polymorphism associated with IL28B in chronic hepatitis C patients. Scand. J. Gastroenterol. 2011, 46, 955–961. [Google Scholar] [CrossRef]
- Miyamura, T.; Kanda, T.; Nakamoto, S.; Wu, S.; Fujiwara, K.; Imazeki, F.; Yokosuka, O. Hepatic STAT1-nuclear translocation and interleukin 28B polymorphisms predict treatment outcomes in hepatitis C virus genotype 1-infected patients. PLoS One 2011, 6, e28617. [Google Scholar]
- Lagging, M.; Askarieh, G.; Negro, F.; Bibert, S.; Soderholm, J.; Westin, J.; Lindh, M.; Romero, A.; Missale, G.; Ferrari, C.; et al. Response prediction in chronic hepatitis C by assessment of IP-10 and IL28B-related single nucleotide polymorphisms. PLoS One. 2011, 6, e17232. [Google Scholar]
- Honda, M.; Sakai, A.; Yamashita, T.; Nakamoto, Y.; Mizukoshi, E.; Sakai, Y.; Yamashita, T.; Nakamura, M.; Shirasaki, T.; Horimoto, K.; et al. Hepatic ISG expression is associated with genetic variation in interleukin 28B and the outcome of IFN therapy for chronic hepatitis C. Gastroenterology 2010, 139, 499–509. [Google Scholar] [CrossRef]
- Fellay, J.; Thompson, A.J.; Ge, D.; Gumbs, C.E.; Urban, T.J.; Shianna, K.V.; Little, L.D.; Qui, P.; Bertelsen, A.H.; Watson, M.; et al. ITPA gene variants protect against anemia in patients treated for chronic hepatitis C. Nature 2010, 464, 40540–40548. [Google Scholar]
- Magg, D.; Castro, C.; Hong, Z.; Cameron, C.E. Hepatitis C virus RNA-dependent RNA polymerase (NS5B) as a mediator of the antiviral activity of ribavirin. J. Biol. Chem. 2001, 276, 46094–46098. [Google Scholar] [CrossRef]
- Contreras, A.M.; Hiasa, Y.; He, W.; Terella, A.; Schmidt, E.V.; Chung, R.T. Viral RNA mutations are region specific and increased by ribavirin in a full-length hepatitis C virus replicon system. J. Virol. 2002, 76, 8505–8517. [Google Scholar] [CrossRef]
- Kanda, T.; Yokosuka, O.; Imazeki, F.; Tanaka, M.; Shino, Y.; Shimada, H.; Tomonaga, T.; Nomura, F.; Nagao, K.; Ochiai, T.; Saisho, H. Inhibition of subgenomic hepatitis C virus RNA in Huh-7 cells: Ribavirin induces mutagenesis in HCV RNA. J. Viral Hepat. 2004, 11, 479–487. [Google Scholar] [CrossRef]
- Zhou, S.; Liu, R.; Baroudy, B.M.; Malcolm, B.A.; Reyes, G.R. The effect of ribavirin and IMPDH inhibitors on hepatitis C virus subgenomic replicon RNA. Virology 2003, 310, 333–342. [Google Scholar] [CrossRef]
- Tam, R.C.; Pai, B.; Bard, J.; Lim, C.; Averett, D.R.; Phan, U.T.; Milovanovic, T. Ribavirin polarizes human T cell responses towards a Type 1 cytokine profile. J. Hepatol. 1999, 30, 376–382. [Google Scholar] [CrossRef]
- Thomas, E.; Feld, J.J.; Li, Q.; Hu, Z.; Fried, M.W.; Liang, T.J. Ribavirin potentiates interferon action by augmenting interferon-stimulated gene induction in hepatitis C virus cell culture models. Hepatology 2011, 53, 32–41. [Google Scholar] [CrossRef]
- Clark, V.; Nelson, D.R. The role of ribavirin in direct acting antiviral drug regimens for chronic hepatitis C. Liver Int. 2012, 32, 103–107. [Google Scholar] [CrossRef]
- Kanda, T.; Imazeki, F.; Yonemitsu, Y.; Mikami, S.; Takada, N.; Nishino, T.; Takashi, M.; Tsubota, A.; Kato, K.; Sugiura, N.; et al. Quantification of hepatitis C virus in patients treated with peginterfron-alfa 2a plus ribavirin treatment by COBAS TaqMan HCV test. J. Viral Hepat. 2011, 18, e292–e297. [Google Scholar] [CrossRef]
- Kanda, T.; Imazeki, F.; Azemoto, R.; Yonemitsu, Y.; Mikami, S.; Kita, K.; Takashi, M.; Sunaga, M.; Wu, S.; Nakamoto, S.; et al. Response to peginterferon-alfa 2b and ribavirin in Japanese patients with chronic hepatitis C genotype 2. Dig. Dis. Sci. 2011, 56, 3335–3342. [Google Scholar] [CrossRef]
- Miyauchi, T.; Kanda, T.; Imazeki, F.; Mikata, R.; Tawada, A.; Arai, M.; Fujiwara, K.; Nakamoto, S.; Wu, S.; Tanaka, T.; et al. Response to peginterferon-alpha 2b and ribavirin in Japanese patients with chronic hepatitis C genotype 1. Hepatology Int. 2012, in press.. [Google Scholar]
- Tsukiyama-Kohara, K.; Yamaguchi, K.; Maki, N.; Ohta, Y.; Miki, K.; Mizokami, M.; Ohba, K.; Tanaka, S.; Hattori, N.; Nomoto, A.; Kohara, M. Antigenicities of group I and II hepatitis C virus polypeptides—Molecular basis of diagnosis. Virology 1993, 192, 430–437. [Google Scholar] [CrossRef]
- Simmonds, P.; Holmes, E.C.; Cha, T.A.; Chan, S.W.; McOmish, F.; Irvine, B.; Beall, E.; Yap, P.L.; Kolberg, J.; Urdea, M.S. Classification of hepatitis C virus into six major genotypes and a series of subtypes by phylogenetic analysis of the NS-5 region. J. Gen. Virol. 1993, 74, 2391–2399. [Google Scholar] [CrossRef]
- McHutchison, J.G.; Manns, M.; Patel, K.; Poynard, T.; Lindsay, K.L.; Trepo, C.; Dienstag, J.; Lee, W.M.; Mak, C.; Garaud, J.J.; Albrecht, J.K. International Hepatitis Interventional Therapy Group. Adherence to combination therapy enhances sustained response in genotype-1-infected patients with chronic hepatitis C. Gastroenterology 2002, 123, 1061–1069. [Google Scholar] [CrossRef]
- Lo Re, V., 3rd.; Teal, V.; Localio, A.R.; Amorosa, V.K.; Kaplan, D.E.; Gross, R. Relationship between adherence to hepatitis C virus therapy and virologic outcomes: A cohort study. Ann. Intern. Med. 2011, 156, 353–360. [Google Scholar]
- Reau, N.; Hadziyannis, S.J.; Messinger, D.; Fried, M.W.; Jensen, D.M. Early predictors of anemia in patients with hepatitis C genotype 1 treated with peginterferon alfa-2a (40KD) plus ribavirin. Am. J. Gastroenterol. 2008, 103, 1981–1988. [Google Scholar] [CrossRef]
- Sulkowski, M.S. Anemia in the treatment of hepatitis C virus infection. Clin. Infect. Dis. 2003, 37, S315–S322. [Google Scholar] [CrossRef]
- Ochi, H.; Maekawa, T.; Abe, H.; Hayashida, Y.; Nakano, R.; Kubo, M.; Tsunoda, T.; Hayes, C.N.; Kumada, H.; Nakamura, Y.; Chayama, K. ITPA polymorphism affects ribavirin-induced anemia and outcomes of therapy—A genome-wide study of Japanese HCV virus patients. Gastroenterology 2010, 139, 1190–1197. [Google Scholar] [CrossRef]
- Sakamoto, N.; Tanaka, Y.; Nakagawa, M.; Yatsuhashi, H.; Nishiguchi, S.; Enomoto, N.; Azuma, S.; Nishimura-Sakurai, Y.; Kakinuma, S.; Nishida, N.; et al. ITPA gene variant protects against anemia induced by pegylated interferon-α and ribavirin therapy for Japanese patients with chronic hepatitis C. Hepatol. Res. 2010, 40, 1063–1071. [Google Scholar] [CrossRef]
- Hitomi, Y.; Cirulli, E.T.; Fellay, J.; McHutchison, J.G.; Thompson, A.J.; Gumbs, C.E.; Shianna, K.V.; Urban, T.J.; Goldstein, D.B. Inosine triphosphate protects against ribavirin-induced adenosine triphosphate loss by adenylosuccinate synthase function. Gastroenterology 2011, 140, 1314–1321. [Google Scholar] [CrossRef]
- Thompson, A.J.; Clark, P.J.; Singh, A.; Ge, D.; Fellay, J.; Zhu, M.; Zhu, Q.; Urban, T.J.; Patel, K.; Tillmann, H.L.; et al. Genome-wide association study of interferon-related cytopenia in chronic hepatitis C patients. J. Hepatol. 2012, 56, 313–319. [Google Scholar]
- Tanaka, Y.; Kurosaki, M.; Nishida, N.; Sugiyama, M.; Matsuura, K.; Sakamoto, N.; Enomoto, N.; Yatsuhashi, H.; Nishiguchi, S.; Hino, K.; et al. Genome-wide association study identified ITPA/DDRGK1 variants reflecting thrombocytopenia in pegylated interferon and ribavirin therapy for chronic hepatitis C. Hum. Mol. Genet. 2011, 20, 3507–3516. [Google Scholar] [CrossRef]
- Kurosaki, M.; Tanaka, Y.; Tanaka, K.; Suzuki, Y.; Hoshioka, Y.; Tamaki, N.; Kato, T.; Yasui, Y.; Hosokawa, T.; Ueda, K.; et al. Relationship between polymorphisms of the inosine triphosphatase gene and anemia or outcome after treatment with pegylated interferon and ribavirin. Antivir. Ther. 2011, 16, 689–694. [Google Scholar]
- Jensen, D.; Pol, S. IL28B genetic polymorphism testing in the era of direct acting antivirals therapy for chronic hepatitis C: Ten years too late? Liver Int. 2012, 32, 74–78. [Google Scholar] [CrossRef]
- Lok, A.S.; Gardiner, D.F.; Lawitz, E.; Martorell, C.; Everson, G.T.; Ghalib, R.; Reindollar, R.; Rustgi, V.; McPhee, F.; Wind-Rotolo, M.; et al. Preliminary study of two antiviral agents for hepatitis C genotype 1. N. Engl. J. Med. 2012, 366, 216–224. [Google Scholar] [CrossRef]
- Omata, M.; Kanda, T.; Yu, M.L.; Yokosuka, O.; Lim, S.G.; Jafri, W.; Tateishi, R.; Hamid, S.S.; Chuang, W.L.; Chutaputti, A.; et al. APASL consensus statements and management algorithms for hepatitis C virus infection. Hepatol. Int. 2012, 6, 409–435. [Google Scholar] [CrossRef]
- Miyaaki, H.; Ichikawa, T.; Yatsuhashi, H.; Taura, N.; Miura, S.; Usui, T.; Mori, S.; Kamihira, S.; Tanaka, Y.; Mizokami, M.; Nakao, K. Suppressor of cytokine signal 3 and IL28 genetic variation predict the viral response to peginterferon and ribavirin. Hepatol. Res. 2011, 41, 1216–1222. [Google Scholar] [CrossRef]
- Tillmann, H.L.; Patel, K.; Muir, A.J.; Guy, C.D.; Li, J.H.; Lao, X.Q.; Thompson, A.; Clark, P.J.; Gardner, S.D.; McHutchison, J.G.; McCarthy, J.J. Beneficial IL28B genotype associated with lower frequency of hepatic steatosis in patients with chronic hepatitis C. J. Hepatol. 2011, 55, 1195–1200. [Google Scholar] [CrossRef]
- Toyoda, H.; Kumada, H. Favorable association between genetic polymorphisms near the IL28B gene and hepatic steatosis: Direct or indirect? J. Hepatol. 2012, 56, 738–739. [Google Scholar] [CrossRef]
- Clark, P.J.; Thompson, A.J.; Zhu, M.; Vock, D.M.; Zhu, Q.; Ge, D.; Patel, K.; Harrison, S.A.; Urban, T.J.; Naggie, S.; et al. Interleukin 28B polymorphisms are the only common genetic variants associated with low-density lipoprotein cholesterol (LDL-C) in genotype-1 chronic hepatitis C and determine the association between LDL-C and treatment response. J. Viral Hepat. 2012, 19, 332–340. [Google Scholar] [CrossRef]
- Rembeck, K.; Alsio, A.; Christensen, P.B.; Farkkila, M.; Langeland, N.; Buhl, M.R.; Pedersen, C.; Morch, K.; Westin, J.; Lindh, M.; et al. Impact of IL28B-related single nucleotide polymorphisms on liver histopathology in chronic hepatitis C genotype 2 and 3. PLoS One 2012, 7, e29370. [Google Scholar]
- Stattermayer, A.F.; Rutter, K.; Beinhardt, S.; Scherzer, T.M.; Stadlmayr, A.; Hofer, H.; Wrba, F.; Steindl-Munda, P.; Krebs, M.; Datz, C.; et al. Association of the IL28B genotype with insulin resistance in patients with chronic hepatitis C. J. Hepatol. 2012, in press.. [Google Scholar]
- Veldt, B.J.; Duarte-Rojo, A.; Thompson, A.J.; Watt, K.D.; Heimbach, J.K.; Tillmann, H.L.; Goldstein, D.D.; McHutchison, J.G.; Charlton, M.R. Recipient IL28B polymorphism is an important independent predictor of posttransplant diabetes mellitus in liver transplant patients with chronic hepatitis C. Am. J. Transplant. 2012, 12, 737–744. [Google Scholar]
- Marabita, F.; Aghemo, A.; Nicola, S.D.; Rumi, M.G.; Cheroni, C.; Scavelli, R.; Crimi, M.; Soffredini, R.; Abrignani, S.; De Francesco, R.; Colombo, M. Genetic variation in the interleukin-28B gene is not associated with fibrosis progression in patients with chronic hepatitis C and known date of infection. Hepatology 2011, 54, 1127–1134. [Google Scholar] [CrossRef]
- Joshita, S.; Umemura, T.; Katsuyama, Y.; Ichikawa, Y.; Kimura, T.; Morita, S.; Kamijo, A.; Komatsu, M.; Ichijo, T.; Matsumoto, A.; et al. Association of IL28B gene polymorphism with development of hepatocellular carcinoma in Japanese patients with chronic hepatitis C virus infection. Hum. Immunol. 2012, 73, 298–300. [Google Scholar] [CrossRef]
- Bochud, P.Y.; Bibert, S.; Kutalik, Z.; Patin, E.; Guergnon, J.; Nalpas, B.; Goossens, N.; Kuske, L.; Mullhaupt, B.; Gerlach, T.; et al. IL28B alleles associated with poor hepatitis C virus (HCV) clearance protect against inflammation and fibrosis in patients infected with non-1 HCV genotypes. Hepatology 2012, 55, 384–394. [Google Scholar] [CrossRef]
- Di Marco, V.; Bronte, F.; Calvaruso, V.; Capra, M.; Borsellino, Z.; Maggio, A.; Renda, M.C.; Pitrolo, L.; Pinto, M.C.L.; Rizzo, M.; et al. IL28B polymorphisms influence stage of the liver fibrosis and spontaneous or interferon-induced viral clearance in thalassemia patients with hepatitis C virus infection. Haematologica 2012, in press.. [Google Scholar]
- Eurich, D.; Boas-Knoop, S.; Bahra, M.; Neuhaus, R.; Somasundaram, R.; Neuhaus, P.; Neumann, U.; Seehofer, D. Role of IL28B polymorphism in the development of hepatitis C virus-induced hepatocellular carcinoma, graft fibrosis, and posttransplant antiviral therapy. Transplantation 2012, 93, 644–649. [Google Scholar]
- Masuzaki, R.; Tateishi, R.; Yoshida, H.; Arano, T.; Uchino, K.; Enooku, K.; Goto, E.; Nakagawa, H.; Asaoka, Y.; Kondo, Y.; et al. Assessment of disease progression in patients with transfusion-associated chronic hepatitis C using transient elastography. World J. Gastroenterol. 2012, 18, 1385–1390. [Google Scholar] [CrossRef]
- Osada, M.; Kaneko, M.; Sakamoto, M.; Endoh, M.; Takigawa, K.; Suzuki-Inoue, O.; Satoh, K.; Enomoto, N.; Yatomi, Y.; Ozaki, Y. Causes of thrombocytopenia in chronic hepatitis C viral infection. Clin. Appl. Thromb. Hemost. 2012, 18, 272–280. [Google Scholar] [CrossRef]
- Ohira, M.; Ishifuro, M.; Ide, K.; Irei, T.; Tashiro, H.; Itamoto, T.; Ito, K.; Chayama, K.; Asahara, T.; Ohdan, H. Significant correlation between spleen volume and thrombocytopenia in liver transplant patients: A concept for predicting persistent thrombocytopenia. Liver Transpl. 2009, 15, 208–215. [Google Scholar] [CrossRef]
- Olariu, M.; Olariu, C.; Olteanu, D. Thrombocytopenia in chronic hepatitis C. J. Gastrointestin. Liver Dis. 2010, 19, 381–385. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Miyamura, T.; Kanda, T.; Nakamoto, S.; Wu, S.; Jiang, X.; Arai, M.; Fujiwara, K.; Imazeki, F.; Yokosuka, O. Roles of ITPA and IL28B Genotypes in Chronic Hepatitis C Patients Treated with Peginterferon Plus Ribavirin. Viruses 2012, 4, 1264-1278. https://doi.org/10.3390/v4081264
Miyamura T, Kanda T, Nakamoto S, Wu S, Jiang X, Arai M, Fujiwara K, Imazeki F, Yokosuka O. Roles of ITPA and IL28B Genotypes in Chronic Hepatitis C Patients Treated with Peginterferon Plus Ribavirin. Viruses. 2012; 4(8):1264-1278. https://doi.org/10.3390/v4081264
Chicago/Turabian StyleMiyamura, Tatsuo, Tatsuo Kanda, Shingo Nakamoto, Shuang Wu, Xia Jiang, Makoto Arai, Keiichi Fujiwara, Fumio Imazeki, and Osamu Yokosuka. 2012. "Roles of ITPA and IL28B Genotypes in Chronic Hepatitis C Patients Treated with Peginterferon Plus Ribavirin" Viruses 4, no. 8: 1264-1278. https://doi.org/10.3390/v4081264
APA StyleMiyamura, T., Kanda, T., Nakamoto, S., Wu, S., Jiang, X., Arai, M., Fujiwara, K., Imazeki, F., & Yokosuka, O. (2012). Roles of ITPA and IL28B Genotypes in Chronic Hepatitis C Patients Treated with Peginterferon Plus Ribavirin. Viruses, 4(8), 1264-1278. https://doi.org/10.3390/v4081264