Effects and Effectiveness of Two RNAi Constructs for Resistance to Pepper golden mosaic virus in Nicotiana benthamiana Plants


Abstract
:1. Introduction
2. Results
2.1. Development of CIRP (Construct of the Intergenic Region of PepGMV) and CIRT (Construct of the Intergenic Region of ToChLPV) Constructs
2.2. Resistance Assays and Determination of Severity, Incidence and Viral Load


{kind=link}
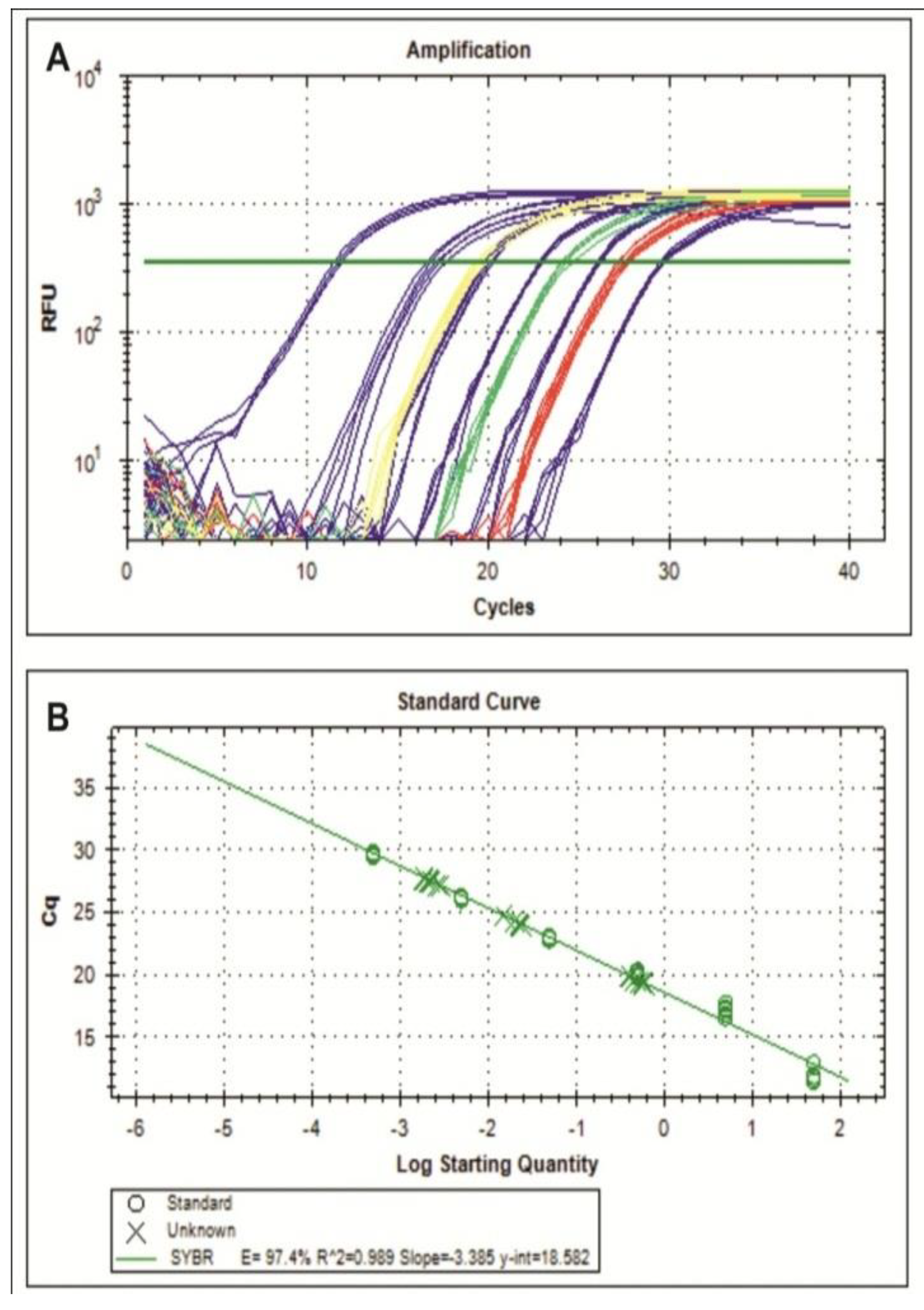{kind=link}
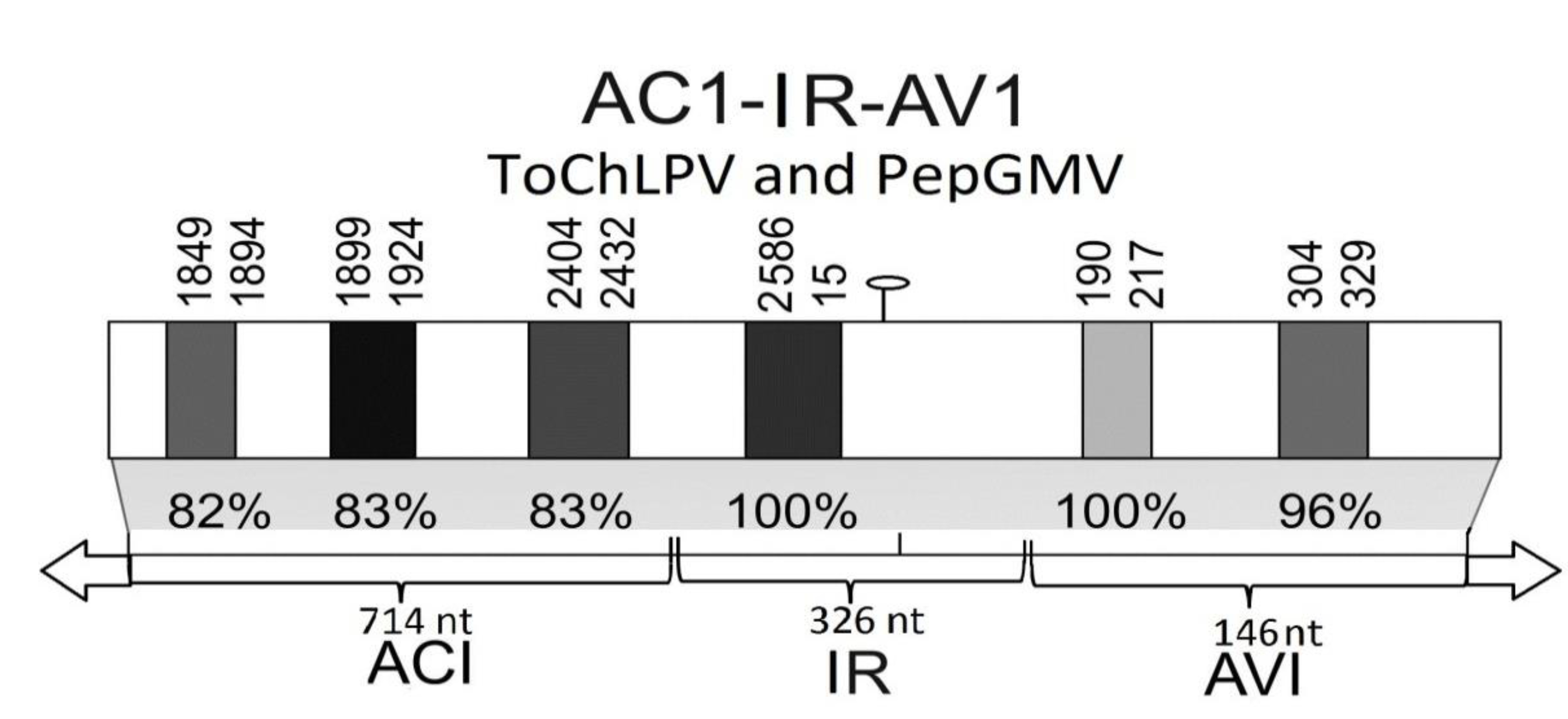{kind=link}
| Severity | Incidence | Viral load | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Constructs | Foliar severity range 1 | Percentage of foliar damage (%) 2 | Efficacy (%) 3 | PCR (+/-) | Adjusted data (%) | Efficacy (%) | Copies | Folds | Efficacy (%) | |||
| CIRT | 1.375 | 55 a | 45 | 4/4 | 57.4 c | 42.6 | 2.27 × 10-2 c | 19.3 | 95.6 | |||
| CIRP | 0.50 | 20 b | 80 | 3/5 | 42.8 b | 57.2 | 2.20 × 10-3 b | 237.3 | 99.56 | |||
| Positive control | 2.5 | 100 a | 0 | 8/0 | 100 a | 0 | 5.22 × 10-1 a | 1 | 0 | |||

2.3. Gene Expression Related to RNAi
2.4. Discussion

3. Experimental Section
3.1. Development of RNAi Constructs
3.2. Design of the Experimental Bioassay
3.2.1. Inoculation Assays
3.2.2. Evaluation of variables
A. Severity
B. Incidence
C. Viral Load
3.2.3. Determination of the Efficacy of RNAi Constructs
3.3. Microarray Data
4. Conclusions
Acknowledgments
Conflicts of Interest
References and Notes
- Brown, J.; Fauquet, C.; Briddon, R.; Zerbini, F.; Moriones, E.; Navas-Castillo, J. Family Geminiviridae . In Virus Taxonomy 9th Report of the International Committee on Taxonomy of Viruses; Adams, M., Carstens, E., Lefkowitz, E., Eds.; Elsevier Academic Press: London, UK, 2011; pp. 351–373. [Google Scholar]
- Rojas, M.; Hagen, C.; Lucas, W.; Gilbertson, R. Exploiting chinks in the plant's armor: evolution and emergence of geminiviruses. Annu. Rev. Phytopathol. 2005, 43, 361–394. [Google Scholar]
- Navas-Castillo, J.; Fiallo-Olivé, E.; Sánchez-Campos, S. Emerging virus diseases transmitted by whiteflies. Annu. Rev. Phytopathol. 2011, 49, 219–248. [Google Scholar]
- Morales, F. Distribution and dissemination of begomoviruses in Latin America and the Caribbean. In Bemisia: Bionomics and Management of a Global Pest; Springer, Netherlands, 2010; pp. 283–318. [Google Scholar]
- Méndez-Lozano, J.; Torres-Pacheco, I.; Fauquet, C.; Rivera-Bustamante, R. Interactions between geminiviruses in a naturally occurring mixture: Pepper huasteco virus and Pepper golden mosaic virus. Phytopathology. 2003, 93, 270–277. [Google Scholar]
- Rentería-Canett, I.; Xoconostle-Cázares, B.; Ruiz-Medrano, R.; Rivera-Bustamante, R. Geminivirus mixed infection on pepper plants: synergistic interaction between PHYVV and PepGMV. Virol. J. 2011, 8, 104–117. [Google Scholar]
- Lebsky, V.; Hernandez-Gonzalez, J.; Arguello-Astorga, G.; Cardenas-Conejo, Y.; Poghosyan, A.; Bertaccini, A.; Maini, S. Detection of phytoplasmas in mixed infection with begomoviruses: a case study of tomato and pepper in Mexico. Bulletin of Insectology. 2011, 64, 55–56. [Google Scholar]
- Holguín-Peña, R.; Arguello-Astorga, G.; Brown, J.; Rivera-Bustamante, R. A new strain of Tomato chino La Paz virus associated with a leaf curl disease of tomato in Baja California Sur, Mexico. Plant Dis. 2006, 90, 973. [Google Scholar]
- Gámez-Jiménez, C. Virus del enrollamiento de la hoja amarilla del tomate (TYLCV): Distribución en el norte de México, identificación de hospedantes alternos y variabilidad genética. PhD thesis, Centro de Investigacion para el Desarrollo Integral Regional CIIDIR-Sinaloa, México, 2007. [Google Scholar]
- Mauricio-Castillo, J. Métodos moleculares que potencian el descubrimiento de nuevas especies de begomovirus y la detección de infecciones mixtas. PhD thesis, Instituto Potosino de Investigación Científica y Tecnológica, A.C., San Luis Potosi; México, 2006. [Google Scholar]
- Cardenas-Conejo, Y.; Arguello-Astorga, G.; Poghosyan, A.; Hernandez-Gonzalez, J.; Lebsky, V.; Holguin-Peña, J.; Medina-Hernández, D.; Vega-Peña, S. First report of Tomato yellow leaf curl virus co-infecting pepper with Tomato chino La Paz virus in Baja California Sur, México. Plant Dis. 2010, 94, 1266–1267. [Google Scholar]
- Seal, S.; Vanden Bosch, F.; Jeger, M. Factors influencing begomovirus evolution and their increasing global significance: implications for sustainable control. Crit Rev Plant Sci. 2006, 25, 23–46. [Google Scholar]
- Desbiez, C.; Moury, B.; Lecoq, H. The hallmarks of “green” viruses: Do plant viruses evolve differently from the others? Infect Genet Evol. 2011, 11, 812–824. [Google Scholar] [CrossRef]
- Vanderschuren, H.; Stupak, M.; Fütterer, J.; Gruissem, W.; Zhang, P. Engineering resistance to geminiviruses–review and perspectives. Plant Biotechnol J. 2007, 5, 207–220. [Google Scholar]
- Shepherd, D.; Martin, D.; Thomson, J. Transgenic strategies for developing crops resistant to geminiviruses. Plant Sci. 2009, 176, 1–11. [Google Scholar] [CrossRef]
- Raja, P.; Wolf, J.; Bisaro, D. RNA silencing directed against geminiviruses: post-transcriptional and epigenetic components. BBA Gene Regul. Mech. 2010, 1799, 337–351. [Google Scholar]
- Vanitharani, R.; Chellappan, P.; Fauquet, C. Short interfering RNA-mediated interference of gene expression and viral DNA accumulation in cultured plant cells. ProcNatlAcad Sci. 2003, 100, 9632–9636. [Google Scholar]
- Lucioli, A.; Noris, E.; Brunetti, A.; Tavazza, R.; Ruzza, V.; Castillo, A.; Tavazza, M. Tomato yellow leaf curl Sardinia virus rep-derived resistance to homologous and heterologous geminiviruses occurs by different mechanisms and is overcome if virus-mediated transgene silencing is activated. J. Virol. 2003, 77, 6785–6798. [Google Scholar]
- Vanderschuren, H.; Alder, A.; Zhang, P.; Gruissem, W. Dose-dependent RNAi-mediated geminivirus resistance in the tropical root crop cassava. Plant Mol. Biol. 2009, 70, 265–272. [Google Scholar] [CrossRef]
- Ramesh, S.; Mishra, A.; Praveen, S. Hairpin RNA-mediated strategies for silencing of tomato leaf curl virus AC1 and AC4 genes for effective resistance in plants. Oligonucleotides. 2007, 17, 251–257. [Google Scholar] [CrossRef]
- Frizzi, A.; Huang, S. Tapping RNA silencing pathways for plant biotechnology. Plant Biotechnol. J. 2010, 8, 655–677. [Google Scholar] [CrossRef]
- Lin, C.; Ku, H.; Tsai, W.; Green, S.; Jan, F. Resistance to a DNA and a RNA virus in transgenic plants by using a single chimeric transgene construct. Transgenic Res. 2011, 20, 261–270. [Google Scholar]
- Prins, M.; Laimer, M.; Noris, E.; Schubert, J.; Wassenegger, M.; Tepfer, M. Strategies for antiviral resistance in transgenic plants. Mol. Plant Pathol. 2008, 9, 73–83. [Google Scholar]
- Duan, C.; Wang, C.; Guo, H. Application of RNA silencing to plant disease resistance. Silence. 2012, 3, 5. [Google Scholar] [CrossRef]
- Abhinav, K. RNAi: A Promising Approach to Develop Transgenic Plants Against Geminiviruses and Insects. J. Plant Physiol. Pathol. 2013, 1, 1. [Google Scholar]
- Chellappan, P.; Masona, M.; Vanitharani, R.; Taylor, N.; Fauquet, C. Broad spectrum resistance to ssDNA viruses associated with transgene-induced gene silencing in cassava. Plant Mol. Biol. 2004, 56, 601–611. [Google Scholar]
- Chellappan, P.; Vanitharani, R.; Fauquet, C. Short interfering RNA accumulation correlates with host recovery in DNA virus-infected hosts, and gene silencing targets specific viral sequences. J. Virol. 2004, 78, 7465–7477. [Google Scholar] [CrossRef]
- Abhary, M.; Anfoka, G.; Nakhla, M.; Maxwell, D. Post-transcriptional gene silencing in controlling viruses of the Tomato yellow leaf curl virus complex. Arch. Virol. 2006, 151, 2349–2363. [Google Scholar]
- Ribeiro, S.; Lohuis, H.; Goldbach, R.; Prins, M. Tomato chlorotic mottle virus is a target of RNA silencing but the presence of specific short interfering RNAs does not guarantee resistance in transgenic plants. J. Virol. 2007, 81, 1563–1573. [Google Scholar]
- González-Pérez, J.; Espino-Gudiño, M.; Torres-Pacheco, I.; Guevara-González, R.; Herrera-Ruiz, G.; Rodríguez-Hernández, V. Quantification of virus syndrome in chili peppers. Afr. J. Biotechnol. 2011, 10, 5236–5250. [Google Scholar]
- Reyes, M.; Nash, T.; Dallas, M.; Ascencio-Ibáñez, J.; Hanley-Bowdoin, L. Peptide Aptamers That Bind to Geminivirus Replication Proteins Confer a Resistance Phenotype to Tomato Yellow Leaf Curl Virus and Tomato Mottle Virus Infection in Tomato. J. Virol. 2013, 87, 9691–9706. [Google Scholar]
- Mason, G.; Caciagli, P.; Accotto, G.; Noris, E. Real-time PCR for the quantitation of Tomato yellow leaf curl Sardinia virus in tomato plants and in Bemisiatabaci. J. Virol. Methods. 2008, 147, 282–289. [Google Scholar] [CrossRef]
- Lapidot, M.; Friedmann, M. Breeding for resistance to whitefly̵transmitted geminiviruses. Ann. Appl. Biol. 2002, 140, 109–127. [Google Scholar] [CrossRef]
- Ascencio-Ibáñez, J.; Sozzani, R.; Lee, T.; Chu, T.; Wolfinger, R.; Cella, R.; Hanley-Bowdoin, L. Global analysis of Arabidopsis gene expression uncovers a complex array of changes impacting pathogen response and cell cycle during geminivirus infection. Plant Physiol. 2008, 148, 436–454. [Google Scholar]
- Rowe, J.; Dunigan, D.; Blanc, G.; Gurnon, J.; Xia, Y.; Van Etten, J. Evaluation of higher plant virus resistance genes in the green alga, Chlorellavariabilis NC64A, during the early phase of infection with Paramecium bursaria chlorella virus-1. Virology. 2013, 442, 101–113. [Google Scholar]
- Wyatt, S.; Brown, J. Detection of subgroup III geminivirus isolates in leaf extracts by degenerate primers and polymerase chain reaction. Phytopathology. 1996, 86, 1288–1293. [Google Scholar] [CrossRef]
- Tenllado, F.; Llave, C.; Díaz-Ruíz, J. RNA interference as a new biotechnological tool for the control of virus diseases in plants. Virus Res. 2004, 102, 85–96. [Google Scholar] [CrossRef]
- Benedito, V.; Visser, P.; Angenent, G.; Krens, F. The potential of virus-induced gene silencing for speeding up functional characterization of plant genes. Genet. Mol. Res. 2004, 3, 323–341. [Google Scholar]
- Vanitharani, R.; Chellappan, P.; Fauquet, C. Geminiviruses and RNA silencing. Trends Plant Sci. 2005, 10, 144–151. [Google Scholar] [CrossRef]
- Raja, P.; Sanville, B.; Buchmann, R.; Bisaro, D. Viral genome methylation as an epigenetic defense against geminiviruses. J. Virol. 2008, 82, 8997–9007. [Google Scholar]
- Pallas, V.; García, J. How do plant viruses induce disease? Interactions and interference with host components. J. Gen. Virol. 2011, 92, 2691–2705. [Google Scholar] [CrossRef]
- Lipp, M.; Shillito, R.; Giroux, R.; Spiegelhalter, F.; Charlton, S.; Pinero, D.; Song, P. Polymerase chain reaction technology as analytical tool in agricultural biotechnology. J. AOAC Int. 2005, 88, 136–155. [Google Scholar]
- Chellappan, P.; Vanitharani, R.; Ogbe, F.; Fauquet, C. Effect of temperature on geminivirus-induced RNA silencing in plants. Plant Physiol. 2005, 138, 1828–1841. [Google Scholar]
- Rojas, M.; Gilbertson, R.; Maxwell, D. Use of degenerate primers in the polymerase chain reaction to detect whitefly-transmitted geminiviruses. Plant Dis. 1993, 77, 340–347. [Google Scholar]
- Karimi, M.; Inzé, D.; Depicker, A. GATEWAY™ vectors for Agrobacterium mediated plant transformation. Trends Plant Sci. 2002, 7, 193–195. [Google Scholar] [CrossRef]
- Tenllado, F.; Dıaz-Ruız, J. Double-stranded RNA-mediated interference with plant virus infection. J. Virol. 2001, 75, 12288–12297. [Google Scholar] [CrossRef]
- Carrillo-Tripp, J.; Lozoya-Gloria, E.; Rivera-Bustamante, R. Symptom remission and specific resistance of pepper plants after infection by Pepper golden mosaic virus. Phytopathology. 2007, 97, 51–59. [Google Scholar]
- Vander Plank, E. Plants diseases, Epidemiology and Control. Academia press: New York, 1963; 69. [Google Scholar]
- Nicot, N.; Hausman, J.; Hoffmann, L.; Evers, D. Housekeeping gene selection for real-time RT-PCR normalization in potato during biotic and abiotic stress. J. Exp. Bot. 2005, 56, 2907–2914. [Google Scholar]
- Péréfarres, F.; Hoareau, M.; Chiroleu, F.; Reynaud, B.; Dintinger, J.; Lett, J. A novel synthetic quantification standard including virus and internal report targets: application for the detection and quantification of emerging begomoviruses on tomato. Virol. J. 2011, 8, 1–9. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Medina-Hernández, D.; Rivera-Bustamante, R.F.; Tenllado, F.; Holguín-Peña, R.J. Effects and Effectiveness of Two RNAi Constructs for Resistance to Pepper golden mosaic virus in Nicotiana benthamiana Plants. Viruses 2013, 5, 2931-2945. https://doi.org/10.3390/v5122931
Medina-Hernández D, Rivera-Bustamante RF, Tenllado F, Holguín-Peña RJ. Effects and Effectiveness of Two RNAi Constructs for Resistance to Pepper golden mosaic virus in Nicotiana benthamiana Plants. Viruses. 2013; 5(12):2931-2945. https://doi.org/10.3390/v5122931
Chicago/Turabian StyleMedina-Hernández, Diana, Rafael Francisco Rivera-Bustamante, Francisco Tenllado, and Ramón Jaime Holguín-Peña. 2013. "Effects and Effectiveness of Two RNAi Constructs for Resistance to Pepper golden mosaic virus in Nicotiana benthamiana Plants" Viruses 5, no. 12: 2931-2945. https://doi.org/10.3390/v5122931
APA StyleMedina-Hernández, D., Rivera-Bustamante, R. F., Tenllado, F., & Holguín-Peña, R. J. (2013). Effects and Effectiveness of Two RNAi Constructs for Resistance to Pepper golden mosaic virus in Nicotiana benthamiana Plants. Viruses, 5(12), 2931-2945. https://doi.org/10.3390/v5122931