Characterization of Clade 2.3.2.1 H5N1 Highly Pathogenic Avian Influenza Viruses Isolated from Wild Birds (Mandarin Duck and Eurasian Eagle Owl) in 2010 in Korea

Abstract
:1. Introduction
2. Results
2.1. Virus Isolation
2.2. The Disease Situation on Duck Farms during 2010/2011 H5N1 HPAI Outbreak in Korea
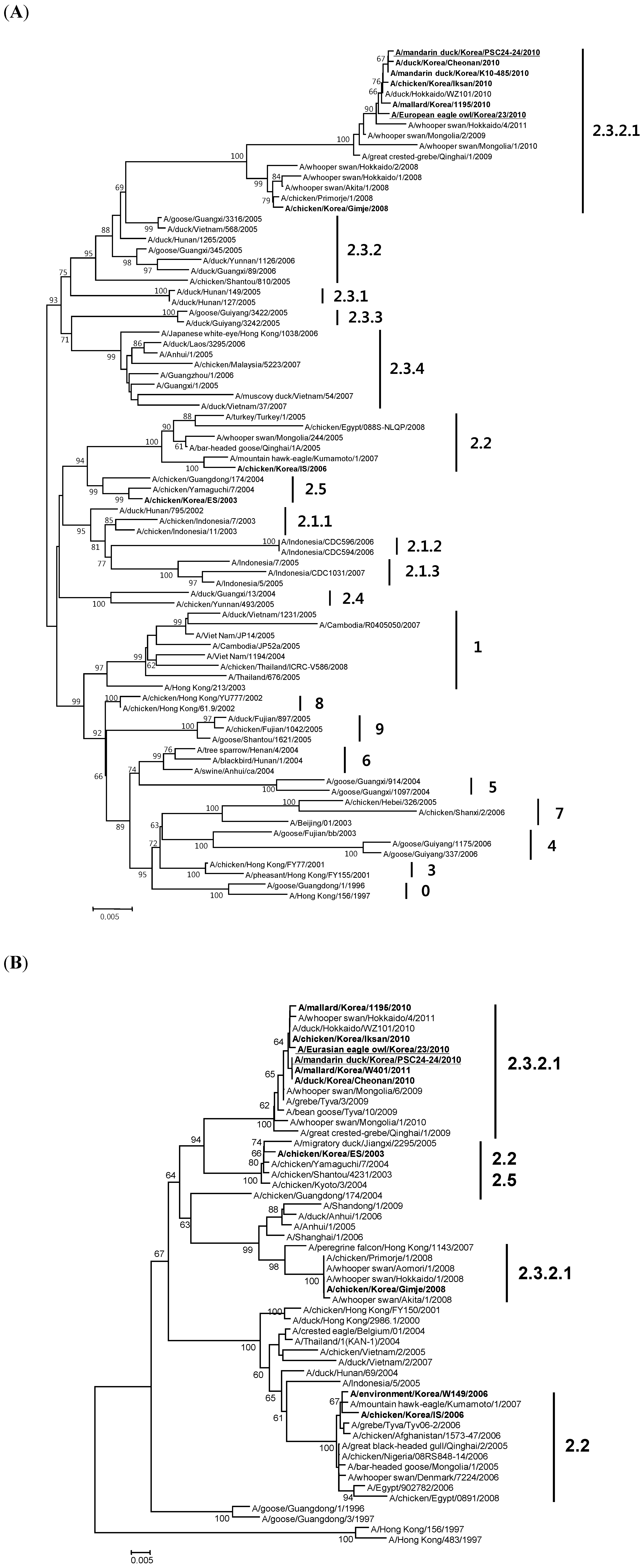
2.3. Genetic and Phylogenetic Analysis
2.4. Virus Replication in Experimentally Inoculated Shickens, Ducks and Mice
2.4.1. Chicken noculation experiments
2.4.2. Duck inoculation experiments
2.4.3. Mouse inoculation experiments

{kind=link}
{kind=link}
{kind=link}
| Duck type | Farm locationa | Age (weeks) | Flock size | Clinical signs | Mortalityb (%) | Remarks |
|---|---|---|---|---|---|---|
| Breeder duck | Cheonan, Chungnam | 36 | 10,700 | Greenish diarrhea, Reduced egg production (61%) | - | Index farm |
| Reduced feed consumption (20%) | ||||||
| Anseong, Gyeonggi | 70/88 | 45,000 | Suppression, Reduced egg production (80%) | - | ||
| Cheonan, Chungnam | 66 | 13,360 | Greenish diarrhea, Reduced egg production (50%) | 0.1% | ||
| Reduced feed consumption (20%) | ||||||
| Icheon, Gyeonggi | 52/56 | 8,300 | Suppression, Reduced egg production (over 70%) | Rare death | ||
| Hwaseong, Gyeonggi | 18 | 8,000 | Reduced feed consumption, Greenish diarrhea | - | ||
| Meat type duck | Yeongam, Jeonnam | 5 | 14,500 | Suppression, Increased mortality | 31.0 | |
| Yeongam, Jeonnam | 5 | 10,600 | Suppression, Increased mortality | 4.7 | ||
| Yeongam, Jeonnam | 3 | 20,000 | Suppression, Increased mortality | 7.9 | ||
| Naju, Jeonnam | 4 | 15,000 | Suppression, Increased mortality | 8.0 | ||
| Yeongam, Jeonnam | 5 | 11,200 | Suppression, Increased mortality | 14.6 | ||
| Naju, Jeonnam | 4 | 5,000 | Torticollis, Leg paralysis, Greenish diarrhea | > 2.7 | ||
| Increased mortality | ||||||
| Damyang, Jeonnam | 1 | 13,300 | Suppression, Diarrhea, Cyanosis of beak | 17.3 |


| Virus | Inoculation route | Virus titer (log10 TCID50/0.1 ml, mean ± standard deviation) | MDTa (hours) | IVPIb | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Brain | Trachea | Lung | Spleen | Kidney | Heart | Muscle | Cecal tonsil | ||||
| PSC24-24 | Intravenous | nt | nt | nt | nt | nt | nt | nt | nt | 58.2 | 2.74 |
| EEO/23 | Intravenous | nt | nt | nt | nt | nt | nt | nt | nt | 44.1 | 2.86 |
| Intranasal | 5.1 ± 0.5 | 5.2 ± 0.5 | 5.8 ± 0.7 | 5.0 ± 0.7 | 5.2 ± 0.8 | 5.6 ± 0.7 | 4.7 ± 0.8 | 4.9 ± 0.9 | 75 | na | |
| Virus | Group | Sample | virus titer (log10 TCID50/0.1 ml, mean±standard deviation) | HI titer c | |||||
|---|---|---|---|---|---|---|---|---|---|
| 1dpi | 3dpi | 5dpi | 7dpi | 10dpi | 14dpi | ||||
| PSC24-24 | Inoculated | OP | 1.6±0.1 (4/8) a | 2.8±0.7 (8/8) | 2.6±0.8 (6/6) | + (2/4) | - (0/4) | - (0/3) | 5.3±0.6 |
| CL | - (0/8) | 1.9±1.1 (6/8) | 1.8±0.9 (4/6) | 2.4±1.0 (3/4) | 0.9 (1/4) | - (0/3) | |||
| Contact | OP | - (0/3) | 2.2±0.6 (2/3) | 2.1±0.4 (3/3) | 2.0±1.4 (3/3) | - (0/2) | - (0/2) | 5.0±0.0 | |
| CL | - (0/3) | - (0/3) | 1.4±0.1 (2/3) | 1.1±0.8 (3/3) | 0.6 (1/2) | - (0/2) | |||
| EEO/23 | Inoculated | OP | + (2/8) | 2.6±0.6 (8/8) | 2.6±1.7 (5/6) | 1.9±1.0 (4/4) | nt | nt | nt |
| CL | + (3/8) | 1.2±0.8 (5/8) | 1.4±0.7 (6/6) | - (0/4) | nt | nt | |||
| Contact b | OP | - (0/3) | 2.5 (1/2) | 1.6 (2/2) | 0.8 (2/2) | + (1/1) | - (0/1) | 7.0 | |
| CL | - (0/3) | - (0/2) | + (2/2) | 2.5 (1/2) | - (0/1) | - (0/1) | |||
| Virus | Route | Animal status | Number of tested | Virus titer (log10 TCID50/0.1 ml, mean ± standard deviation) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Brain | Trachea | Lung | Spleen | Kidney | Heart | Muscle | Cecal tonsil | ||||
| PSC24-24 | Inoculated | Dead a | 3 | 5.2 ± 0.5 | 3.5 ± 1.0 | 4.1 ± 0.6 | - | 4.0 ± 0.8 | 4.1 ± 2.1 | 4.2 | 3.4 |
| Sacrificed b | 2 | 3.1 ± 0.4 | 5.6 ± 0.1 | 4.6 ± 0.1 | 3.4 ± 0.8 | 4.0 ± 0.5 | 4.5 ± 1.1 | 3.4 ± 1.1 | 2.4 ± 0.2 | ||
| Contact | Dead c | 1 | 2.6 | 2.5 | 3.6 | 1.6 | 3.5 | 4.5 | - | 2.6 | |
| EEO/23 | Inoculated | Dead d | 6 | 5.0 ± 1.2 | 4.0 ± 2.2 | 5.1 ± 1.0 | 4.3 ± 2.0 | 4.3 ± 0.9 | 3.7 ± 1.3 | 3.0 ± 0.3 | 3.9 ± 1.0 |
| Sacrificed b | 2 | 2.6 | 3.0 ± 0.3 | 3.6 ± 0.1 | 2.7 ± 0.0 | 2.5 ± 0.1 | 1.8 ± 0.4 | + | 2.4 | ||
| Contact | Dead c, e | 1 | 1.6 | + | + | - | 2.3 | 2.5 | 1.6 | 1.5 | |

| Virus | dpi | Virus titer (log10 TCID50/0.1 ml, mean ± standard deviation) | MLD50 (EID50) | MID50 (EID50) | ||||
|---|---|---|---|---|---|---|---|---|
| Brain | Lung | Liver | Spleen | Kidney | ||||
| PSC24-24 | 3dpi | - | 5.7±0.1 | - | 2.4±0.9 | - | 103.4 | 102.3 |
| 6dpi | 3.4±1.1 | 4.5±0.1 | - | + | - | |||
| EEO/23 | 3dpi | - | 5.7±0.4 | - | 1.2b | - | 103.6 | 101.5 |
| 6dpi | 2.8a | 3.8±0.0c | - | - | - | |||
3. Discussion
4. Experimental Section
4.1. Viruses used in this Study
4.2. Sequencing and Phylogenetic Analysis
4.3. Animal Experiments
4.3.1. Chicken inoculation experiments
4.3.2. Duck inoculation experiments
4.3.3. Mouse inoculation experiments
4.4. Statistical Analysis
5. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- FAO/OIE/WHO. FAO–OIE–WHO Technical update: Current evolution of avian influenza H5N1 viruses. 7 September 2011. Available online: http://www.oie.int/doc/ged/D11128.PDF.
- Li, Y.; Shi, J.; Zhong, G.; Deng, G.; Tian, G.; Ge, J.; Zeng, X.; Song, J.; Zhao, D.; Liu, L.; Jiang, Y.; Guan, Y.; Bu, Z.; Chen, H. Continued evolution of H5N1 influenza viruses in wild birds, domestic poultry, and humans in China from 2004 to 2009. J. Virol. 2010, 84, 8389–8397. [Google Scholar] [CrossRef]
- Jiang, W.–M.; Liu, S.; Chen, J.; Hou, G.–Y.; Li, J.–P.; Cao, Y.–F.; Zhuang, Q.–Y.; Li, Y.; Huang, B.–X.; Chen, J.–M. Molecular epidemiological surveys of H5 subtype highly pathogenic avian influenza viruses in poultry in China during 2007–2009. J. Gen. Virol. 2010, 91, 2491–2496. [Google Scholar] [CrossRef]
- Jadhao, S.; Nguyen, D.; Uyeki, T.; Shaw, M.; Maines, T.; Rowe, T.; Smith, C.; Huynh, L.; Nghiem, H.; Nguyen, D.; Nguyen, H.; Nguyen, H.; Hoang, L.; Nguyen, T.; Phuong, L.; Klimov, A.; Tumpey, T.; Cox, N.; Donis, R.; Matsuoka, Y.; Katz, J. Genetic analysis of avian influenza A viruses isolated from domestic waterfowl in live–bird markets of Hanoi, Vietnam, preceding fatal H5N1 human infections in 2004. Arch. Virol. 2009, 154, 1249–1261. [Google Scholar] [CrossRef]
- Kayali, G.; El–Shesheny, R.; Kutkat, M.A.; Kandeil, A.M.; Mostafa, A.; Ducatez, M.F.; McKenzie, P.P.; Govorkova, E.A.; Nasraa, M.H.; Webster, R.G.; Webby, R.J.; Ali, M.A. Continuing threat of influenza (H5N1) virus circulation in Egypt. Emerg. Infect. Dis. 2011, 17, 2306–2308. [Google Scholar] [CrossRef]
- Islam, M.R.; Haque, M.E.; Giasuddin, M.; Chowdhury, E.H.; Samad, M.A.; Parvin, R.; Nooruzzaman, M.; Rahman, M.M.; Monoura, P. New introduction of clade 2.3.2.1 avian influenza virus (H5N1) into Bangladesh. Transbound. Emerg. Dis. 2012, 59, 460–463. [Google Scholar] [CrossRef]
- WHO/OIE/FAO; H5N1 Evolution Working Group. Continued evolution of highly pathogenic avian influenza A (H5N1): updated nomenclature. Influenza Other Respi. Viruses 2012, 6, 1–5. [Google Scholar] [CrossRef]
- Xu, X.; Subbarao, K.; Cox, N.J.; Guo, Y. Genetic characterization of the pathogenic influenza A/Goose/Guangdong/1/96 (H5N1) virus: similarity of its hemagglutinin gene to those of H5N1 viruses from the 1997 outbreaks in Hong Kong. Virol. 1999, 261, 15–19. [Google Scholar] [CrossRef]
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of influenza A viruses. Microbiol. Rev. 1992, 56, 152–179. [Google Scholar]
- Ellis, T.M.; Barry Bousfield, R.; Bissett, L.A.; Dyrting, K.C.; Luk, G.S.M.; Tsim, S.T.; Sturm–ramirez, K.; Webster, R.G.; Guan, Y.; Peiris, J.S.M. Investigation of outbreaks of highly pathogenic H5N1 avian influenza in waterfowl and wild birds in Hong Kong in late 2002. Avian Pathol. 2004, 33, 492–505. [Google Scholar] [CrossRef]
- Chen, H.; Li, Y.; Li, Z.; Shi, J.; Shinya, K.; Deng, G.; Qi, Q.; Tian, G.; Fan, S.; Zhao, H.; Sun, Y.; Kawaoka, Y. Properties and dissemination of H5N1 viruses isolated during an influenza outbreak in migratory waterfowl in western China. J. Virol. 2006, 80, 5976–5983. [Google Scholar] [CrossRef]
- Chen, H.; Smith, G.J.D.; Zhang, S.Y.; Qin, K.; Wang, J.; Li, K.S.; Webster, R.G.; Peiris, J.S.M.; Guan, Y. Avian flu H5N1 virus outbreak in migratory waterfowl. Nature 2005, 436, 191–192. [Google Scholar]
- Li, Y.; Liu, L.; Zhang, Y.; Duan, Z.; Tian, G.; Zeng, X.; Shi, J.; Zhang, L.; Chen, H. New avian influenza virus (H5N1) in wild birds, Qinghai, China. Emerg. Infect. Dis. 2011, 17, 265–267. [Google Scholar] [CrossRef]
- Sakoda, Y.; Sugar, S.; Batchluun, D.; Erdene–Ochir, T.–O.; Okamatsu, M.; Isoda, N.; Soda, K.; Takakuwa, H.; Tsuda, Y.; Yamamoto, N.; Kishida, N.; Matsuno, K.; Nakayama, E.; Kajihara, M.; Yokoyama, A.; Takada, A.; Sodnomdarjaa, R.; Kida, H. Characterization of H5N1 highly pathogenic avian influenza virus strains isolated from migratory waterfowl in Mongolia on the way back from the southern Asia to their northern territory. Virol. 2010, 406, 88–94. [Google Scholar] [CrossRef]
- Lee, C.–W.; Suarez, D.L.; Tumpey, T.M.; Sung, H.–W.; Kwon, Y.–K.; Lee, Y.–J.; Choi, J.–G.; Joh, S.–J.; Kim, M.–C.; Lee, E.–K.; Park, J.–M.; Lu, X.; Katz, J.M.; Spackman, E.; Swayne, D.E.; Kim, J.–H. Characterization of highly pathogenic H5N1 avian influenza A viruses isolated from South Korea. J. Virol. 2005, 79, 3692–3702. [Google Scholar] [CrossRef]
- Lee, Y.J.; Choi, Y.K.; Kim, Y.J.; Song, M.S.; Jeong, O.M.; Lee, E.K.; Jeon, W.J.; Jeong, W.; Joh, S.J.; Choi, K.S.; Her, M.; Kim, M.C.; Kim, A.; Kim, M.J.; Ho Lee, E.; Oh, T.G.; Moon, H.J.; Yoo, D.W.; Kim, J.H.; Sung, M.H.; Poo, H.; Kwon, J.H.; Kim, C.J. Highly pathogenic avian influenza virus (H5N1) in domestic poultry and relationship with migratory birds, South Korea. Emerg. Infect. Dis. 2008, 14, 487–490. [Google Scholar] [CrossRef]
- Kim, H.–R.; Park, C.–K.; Lee, Y.–J.; Woo, G.–H.; Lee, K.–K.; Oem, J.–K.; Kim, S.–H.; Jean, Y.–H.; Bae, Y.–C.; Yoon, S.–S.; Roh, I.–S.; Jeong, O.–M.; Kim, H.–Y.; Choi, J.–S.; Byun, J.–W.; Song, Y.–K.; Kwon, J.–H.; Joo, Y.–S. An outbreak of highly pathogenic H5N1 avian influenza in Korea, 2008. Vet. Microbiol. 2010, 141, 362–366. [Google Scholar] [CrossRef]
- Kim, H.–R.; Lee, Y.–J.; Park, C.–K.; Oem, J.–K.; Lee, O.–S.; Kang, H.–M.; Choi, J.–G.; Bae, Y.–C. Highly pathogenic avian influenza (H5N1) outbreaks in wild birds and poultry, South Korea. Emerg. Infect. Dis. 2012, 18, 480–483. [Google Scholar]
- Lee, D.–H.; Park, J.–K.; Youn, H.–N.; Lee, Y.–N.; Lim, T.–H.; Kim, M.–S.; Lee, J.–B.; Park, S.–Y.; Choi, I.–S.; Song, C.–S. Surveillance and isolation of HPAI H5N1 from wild mandarin ducks (Aix galericulata). J. Wildlife Dis. 2011, 47, 994–998. [Google Scholar]
- Kim, H.–R.; Kim, B.–S.; Bae, Y.–C.; Moon, O.–K.; Oem, J.–K.; Kang, H.–M.; Choi, J.–G.; Lee, O.S.; Lee, Y.–J. H5N1 subtype highly pathogenic avian influenza virus isolated from healthy mallard captured in South Korea. Vet. Microbiol. 2011, 151, 386–389. [Google Scholar] [CrossRef]
- Kang, H.–M.; Batchuluun, D.; Kim, M.–C.; Choi, J.–G.; Erdene–Ochir, T.–O.; Paek, M.–R.; Sugir, T.; Sodnomdarjaa, R.; Kwon, J.–H.; Lee, Y.–J. Genetic analyses of H5N1 avian influenza virus in Mongolia, 2009 and its relationship with those of eastern Asia. Vet. Microbiol. 2011, 147, 170–175. [Google Scholar] [CrossRef]
- Sakoda, Y.; Ito, H.; Uchida, Y.; Okamatsu, M.; Yamamoto, N.; Soda, K.; Nomura, N.; Kuribayashi, S.; Shichinohe, S.; Sunden, Y.; Umemura, T.; Usui, T.; Ozaki, H.; Yamaguchi, T.; Murase, T.; Ito, T.; Saito, T.; Takada, A.; Kida, H. Reintroduction of H5N1 highly pathogenic avian influenza virus by migratory water birds, causing poultry outbreaks in 2010–2011 winter season in Japan. J. Gen. Virol. 2012, 93, 541–550. [Google Scholar]
- Sharshov, K.; Silko, N.; Sousloparov, I.; Zaykovskaya, A.; Shestopalov, A.; Drozdov, I. Avian influenza (H5N1) outbreak among wild birds, Russia, 2009. Emerg. Infect. Dis. 2010, 16, 349–351. [Google Scholar] [CrossRef]
- Song, J.; Feng, H.; Xu, J.; Zhao, D.; Shi, J.; Li, Y.; Deng, G.; Jiang, Y.; Li, X.; Zhu, P.; Guan, Y.; Bu, Z.; Kawaoka, Y.; Chen, H. The PA protein directly contributes to the virulence of H5N1 avian influenza viruses in domestic ducks. J. Virol. 2011, 85, 2180–2188. [Google Scholar] [CrossRef]
- Choi, C.–Y.; Nam, H.–Y.; Lee, W.–S. Diets and foraging tactics of Eurasian eagle owls (Bubo bubo) in two different habitat types. Kor. J. Env. Eco. 2007, 21, 30–37. [Google Scholar]
- Keawcharoen, J.; van Riel, D.; van Amerongen, G.; Bestebroer, T.; Beyer, W.E.; van Lavieren, R.; Osterhaus, A.D.; Fouchier, R.A.; Kuiken, T. Wild ducks as long–distance vectors of highly pathogenic avian influenza virus (H5N1). Emerg. Infect. Dis. 2008, 14, 600–607. [Google Scholar] [CrossRef]
- Kwon, Y.K.; Thomas, C.; Swayne, D.E. Variability in pathobiology of South Korean H5N1 high–pathogenicity avian influenza virus infection for 5 species of migratory waterfowl. Vet. Pathol. Online 2010, 47, 495–506. [Google Scholar]
- Jeong, O.M.; Kim, M.C.; Kim, M.J.; Kang, H.M.; Kim, H.R.; Kim, Y.J.; Joh, S.J.; Kwon, J.H.; Lee, Y.J. Experimental infection of chickens, ducks and quails with the highly pathogenic H5N1 avian influenza virus. J. Vet. Sci. 2009, 10, 53–60. [Google Scholar]
- Lee, Y.–J.; Kang, H.–M.; Choi, J.–G.; Jeong, O.–M.; Kim, M.–C.; Paek, M.–R.; Kim, H.–R.; Park, C.–K.; Jean, Y.–H.; Roh, I.–S.; Joo, Y.–S.; Kwon, J.–H. Higher virulence of the Korean clade 2.3.2.1 highly pathogenic avian influenza A (H5N1) virus isolated in 2008 in young duckling. J. Zoonoses 2011, 2, 21–27. [Google Scholar]
- Pantin–Jackwood, M.J.; Suarez, D.L.; Spackman, E.; Swayne, D.E. Age at infection affects the pathogenicity of Asian highly pathogenic avian influenza H5N1 viruses in ducks. Virus Res. 2007, 130, 151–161. [Google Scholar]
- Isoda, N.; Sakoda, Y.; Kishida, N.; Bai, G.R.; Matsuda, K.; Umemura, T.; Kida, H. Pathogenicity of a highly pathogenic avian influenza virus, A/chicken/Yamaguchi/7/04 (H5N1) in different species of birds and mammals. Arch. Virol. 2006, 151, 1267–1279. [Google Scholar] [CrossRef]
- Pantin–Jackwood, M.J.; Smith, D.M.; Wasilenko, J.L.; Cagle, C.; Shepherd, E.; Sarmento, L.; Kapczynski, D.R.; Afonso, C.L. Effect of age on the pathogenesis and innate immune responses in Pekin ducks infected with different H5N1 highly pathogenic avian influenza viruses. Virus Res. 2012, 167, 196–206. [Google Scholar] [CrossRef]
- Hulse–Post, D.J.; Franks, J.; Boyd, K.; Salomon, R.; Hoffmann, E.; Yen, H.L.; Webby, R.J.; Walker, D.; Nguyen, T.D.; Webster, R.G. Molecular changes in the polymerase genes (PA and PB1) associated with high pathogenicity of H5N1 influenza virus in mallard ducks. J. Virol. 2007, 81, 8515–8524. [Google Scholar] [CrossRef]
- Marjuki, H.; Scholtissek, C.; Franks, J.; Negovetich, N.; Aldridge, J.; Salomon, R.; Finkelstein, D.; Webster, R. Three amino acid changes in PB1–F2 of highly pathogenic H5N1 avian influenza virus affect pathogenicity in mallard ducks. Arch. Virol. 2010, 155, 925–934. [Google Scholar] [CrossRef]
- Hu, J.; Zhao, K.; Liu, X.; Wang, X.; Chen, Z.; Liu, X. Two highly pathogenic avian influenza H5N1 viruses of clade 2.3.2.1 with similar genetic background but with different pathogenicity in mice and ducks. Transbound. Emerg. Dis. 2012. [Google Scholar] [CrossRef]
- Subbarao, E.K.; London, W.; Murphy, B.R. A single amino acid in the PB2 gene of influenza A virus is a determinant of host range. J. Virol. 1993, 67, 1761–1764. [Google Scholar]
- Li, Z.; Chen, H.; Jiao, P.; Deng, G.; Tian, G.; Li, Y.; Hoffmann, E.; Webster, R.G.; Matsuoka, Y.; Yu, K. Molecular basis of replicaton of duck H5N1 influenza viruses in a mammalian mouse model. J. Virol. 2005, 79, 12058–12064. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, Y.; Shinya, K.; Deng, G.; Jiang, Y.; Li, G.; Guan, Y.; Tian, G.; Li, Y.; Shi, J.; Liu, L.; Zeng, X.; Bu, Z.; Xia, X.; Kawaoka, Y.; Chen, H. Identification of amino acids in HA and PB2 critical for the transmission of H5N1 avian influenza viruses in a mammalian host. PLoS Pathog. 2009, 5, e1000709. [Google Scholar] [CrossRef]
- Gabriel, G.; Dauber, B.; Wolff, T.; Planz, O.; Klenk, H.–D.; Stech, J. The viral polymerase mediates adaptation of an avian influenza virus to a mammalian host. Proc. Natl. Acad. Sci. U. S. A. 2005, 102, 18590–18595. [Google Scholar] [CrossRef]
- Conenello, G.M.; Zamarin, D.; Perrone, L.A.; Tumpey, T.; Palese, P. A single mutation in the PB1–F2 of H5N1 (HK/97) and 1918 influenza A viruses contributes to increased virulence. PLos Pathog. 2007, 3, 1414–1421. [Google Scholar]
- Matrosovich, M.; Tuzikov, A.; Bovin, N.; Gambaryan, A.; Klimov, A.; Castrucci, M.R.; Donatelli, I.; Kawaoka, Y. Early alterations of the receptor–binding properties of H1, H2, and H3 avian influenza virus hemagglutinins after their introduction into mammals. J. Virol. 2000, 74, 8502–8512. [Google Scholar] [CrossRef]
- Maines, T.R.; Lu, X.H.; Erb, S.M.; Edwards, L.; Guarner, J.; Greer, P.W.; Nguyen, D.C.; Szretter, K.J.; Chen, L.–M.; Thawatsupha, P.; Chittaganpitch, M.; Waicharoen, S.; Nguyen, D.T.; Nguyen, T.; Nguyen, H.H.T.; Kim, J.–H.; Hoang, L.T.; Kang, C.; Phuong, L.S.; Lim, W.; Zaki, S.; Donis, R.O.; Cox, N.J.; Katz, J.M.; Tumpey, T.M. Avian influenza (H5N1) viruses isolated from humans in Asia in 2004 exhibit increased virulence in mammals. J. Virol. 2005, 79, 11788–11800. [Google Scholar] [CrossRef]
- Lee, D.–H.; Lee, H.–J.; Lee, Y.–J.; Kang, H.–M.; Jeong, O.–M.; Kim, M.–C.; Kwon, J.–S.; Kwon, J.–H.; Kim, C.–B.; Lee, J.–B.; Park, S.–Y.; Choi, I.–S.; Song, C.–S. DNA barcoding techniques for avian influenza virus surveillance in migratory bird habitats. J. Wildlife Dis. 2010, 46, 649–654. [Google Scholar]
- Choi, J.–G.; Kang, H.–M.; Kim, M.–C.; Paek, M.–R.; Kim, H.–R.; Kim, B.–S.; Kwon, J.–H.; Kim, J.–H.; Lee, Y.–J. Genetic relationship of H3 subtype avian influenza viruses isolated from domestic ducks and wild birds in Korea and their pathogenic potential in chickens and ducks. Vet. Microbiol. 2012, 155, 147–157. [Google Scholar] [CrossRef]
- Hoffmann, E.; Stech, J.; Guan, Y.; Webster, R.G.; Perez, D.R. Universal primer set for the full–length amplification of all influenza A viruses. Arch. Virol. 2001, 146, 2275–2289. [Google Scholar] [CrossRef]
- Li, O.T.W.; Barr, I.; Leung, C.Y.H.; Chen, H.; Guan, Y.; Peiris, J.S.M.; Poon, L.L.M. Reliable universal RT–PCR assays for studying influenza polymerase subunit gene sequences from all 16 haemagglutinin subtypes. J. Virol. Methods 2007, 142, 218–222. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Choi, J.-G.; Kang, H.-M.; Jeon, W.-J.; Choi, K.-S.; Kim, K.-I.; Song, B.M.; Lee, H.-S.; Kim, J.-H.; Lee, Y.-J. Characterization of Clade 2.3.2.1 H5N1 Highly Pathogenic Avian Influenza Viruses Isolated from Wild Birds (Mandarin Duck and Eurasian Eagle Owl) in 2010 in Korea. Viruses 2013, 5, 1153-1174. https://doi.org/10.3390/v5041153
Choi J-G, Kang H-M, Jeon W-J, Choi K-S, Kim K-I, Song BM, Lee H-S, Kim J-H, Lee Y-J. Characterization of Clade 2.3.2.1 H5N1 Highly Pathogenic Avian Influenza Viruses Isolated from Wild Birds (Mandarin Duck and Eurasian Eagle Owl) in 2010 in Korea. Viruses. 2013; 5(4):1153-1174. https://doi.org/10.3390/v5041153
Chicago/Turabian StyleChoi, Jun-Gu, Hyun-Mi Kang, Woo-Jin Jeon, Kang-Seuk Choi, Kwang-Il Kim, Byung Min Song, Hee-Soo Lee, Jae-Hong Kim, and Youn-Jeong Lee. 2013. "Characterization of Clade 2.3.2.1 H5N1 Highly Pathogenic Avian Influenza Viruses Isolated from Wild Birds (Mandarin Duck and Eurasian Eagle Owl) in 2010 in Korea" Viruses 5, no. 4: 1153-1174. https://doi.org/10.3390/v5041153
APA StyleChoi, J.-G., Kang, H.-M., Jeon, W.-J., Choi, K.-S., Kim, K.-I., Song, B. M., Lee, H.-S., Kim, J.-H., & Lee, Y.-J. (2013). Characterization of Clade 2.3.2.1 H5N1 Highly Pathogenic Avian Influenza Viruses Isolated from Wild Birds (Mandarin Duck and Eurasian Eagle Owl) in 2010 in Korea. Viruses, 5(4), 1153-1174. https://doi.org/10.3390/v5041153