Progressive Adaptation of a CpGV Isolate to Codling Moth Populations Resistant to CpGV-M



Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Viruses
2.3. Amplification of Virus
2.4. Evaluation of Viral Production of 2016-r8 and 2016-r16 Isolate
2.5. Bioassays
2.6. DNA Extraction and Restriction Endonuclease (REN) Analysis of CpGV Isolates
3. Results
3.1. REN Analysis


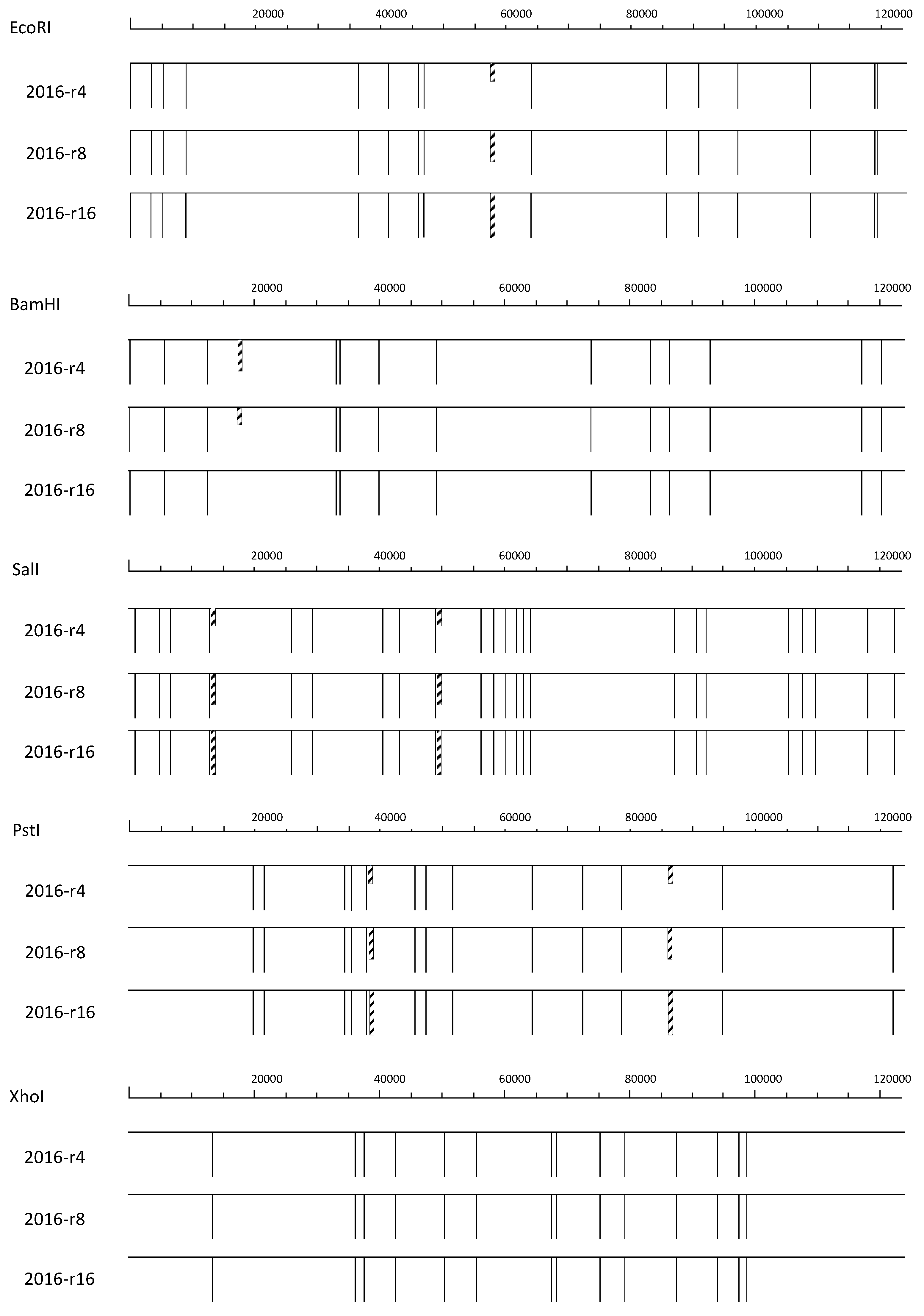{kind=link}
| Virus isolate | Production (per gram of larvae (× 1011 OB/g) ± SE | Production per larvae * (× 1010 OB/g) ± SE |
|---|---|---|
| CpGV-M | 4.84 ± 0.29 a | 2.48 ± 0.10 b |
| 2016-r8 | 5.77 ± 0.29 a | 2.99 ± 0.19 b |
| 2016-r16 | 5.41 ± 0.16 a | 2.54 ± 0.11 b |
| Host colony | Virus isolate | Total No. of insects tested | No. of Occlusion Bodies/µL (95% CI) | Slope ± SE | χ2 | Resistance factor (fold) (a) | ||
|---|---|---|---|---|---|---|---|---|
| LC50 | LC90 | LC50 | LC90 | |||||
| Susceptible | CpGV-M | 786 | 13.10 (6.55–23.20) | 223.10 (110.70–654.18) | 1.04 ± 0.09 | 5.99 | 1.0 | 1.0 |
| NPP-R1 (b) | 689 | 25.80 (14.48–39.93) | 328.55 (196.93–702.51) | 1.16 ± 0.13 | 1.28 | 2.0 | 1.5 | |
| 2016-r4 (b) | 999 | 39.65 (6.40–133.91) | 805.85 (260.20–1.36 × 103) | 0.98 ± 0.11 | 13.60 | 3.0 | 3.6 | |
| 2016-r8 | 445 | 48.37 (21.18–81.44) | 280.52 (158.02–857.03) | 1.68 ± 0.25 | 4.67 | 3.7 | 1.3 | |
| 2016-r16 | 790 | 6.76 (2.60–13.37) | 59.63 (27.54–278.55) | 1.36 ± 0.13 | 11.42 | 0.5 | 0.3 | |
| Resistant | CpGV-M | 396 | 7.84 × 103 (660.45–4.13 × 104) | 1.71 × 106 (2.15 × 105–6.72 × 108) | 0.55 ± 0.09 | 6.32 | 598 | 7664.7 |
| NPP-R1 (b) | 578 | 166.31 (91.21–278.27) | 1.28 × 104 (5.95 × 103–3.80 × 104) | 0.70 ± 0.08 | 4.81 | 12.7 | 57.4 | |
| 2016-r4 (b) | 1201 | 102.31 (63.20–146.91) | 1.57 × 103 (1.01 × 103–2.97 × 103) | 1.10 ± 0.10 | 6.21 | 7.8 | 7.0 | |
| 2016-r8 | 456 | 41.27 (26.97–58.96) | 319.24 (207.87–582.06) | 1.44 ± 0.17 | 1.83 | 3.2 | 1.4 | |
| 2016-r16 | 545 | 22.43 (13.73–34.36) | 410.67 (240.16–846.43) | 1.02 ± 0.11 | 3.60 | 1.7 | 1.8 | |
3.2. Viral Production of Isolates
3.3. Bioassays
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Cross, J.V.; Solomon, M.G.; Chandler, D.; Jarret, P.; Richardson, P.N.; Winstanley, D.; Bathon, H.; Huber, J.; Keller, B.; Langenbruch, G.A.; et al. Biocontrol of Pests of Apples and Pears in Northern and Central Europe: 1. Microbial Agents and Nematodes. Biocontrol. Sci. Technol. 1999, 9, 125–149. [Google Scholar] [CrossRef]
- Willett, M.J.; Neven, L.; Miller, C.E. The occurrence of codling moth in low latitude countries: Validation of pest distribution reports. HortTechnology 2009, 19, 633–637. [Google Scholar]
- Reyes, M.; Franck, P.; Charmillot, P.J.; Ioriatti, C.; Olivares, J.; Pasqualini, E.; Sauphanor, B. Diversity of insecticide resistance mechanisms and spectrum in European populations of the codling moth, Cydia pomonella. Pest Manage. Sci. 2007, 63, 890–902. [Google Scholar] [CrossRef]
- Gröner, A. Specificity and safety of baculoviruses. In The Biology of Baculoviruses: Biological Properties and Molecular Biology; Granados, R.R., Federici, B.A., Eds.; Academic Press: San Diego, CA., USA, 1986; Volume 1, pp. 177–202. [Google Scholar]
- Moscardi, F.; Souza, M.L.; Castro, M.E.B.; Moscardi, M.; Szewczyk, B. Baculovirus Pesticides: Present State and Future Perspectives. In Microbes and Microbial Technology, 1st ed.; Ahmad, I., Ahmad, F., Pichtel, J., Eds.; Springer: New York, NY, USA, 2011; pp. 415–445. [Google Scholar]
- Rohrmann, G.F. Baculovirus Molecular Biology; National Library of Medicine: Bethesda, MD, USA, 2013. [Google Scholar]
- Thiem, S.M.; Cheng, X.W. Baculovirus host-range. Virologica Sinica 2009, 24, 436–457. [Google Scholar] [CrossRef]
- Jehle, J.A.; Blissard, G.W.; Bonning, B.C.; Cory, J.S.; Herniou, E.A.; Rohrmann, G.F.; Theilmann, D.A.; Thiem, S.M.; Vlak, J.M. On the classification and nomenclature of baculoviruses: A proposal for revision. Arch. Virol. 2006, 151, 1257–1266. [Google Scholar] [CrossRef] [PubMed]
- Tanada, Y. A granulosis virus of the codling moth, Carpocapsa pomonella (Linnaeus) (Olethreutidae, Lepidoptera). J. Insect Pathol. 1964, 6, 378–380. [Google Scholar]
- Lacey, L.A.; Thompson, D.; Vincent, C.; Arthurs, S.P. Codling moth granulovirus: A comprehensive review. Biocontrol. Sci. Techn. 2008, 18, 639–663. [Google Scholar] [CrossRef]
- Crook, N.E.; Spencer, R.A.; Payne, C.C.; Leisy, D.J. Variation in Cydia pomonella granulosis virus isolates and physical maps of the DNA from three variants. J. Gen. Virol. 1985, 66, 2423–2430. [Google Scholar] [CrossRef]
- Luque, T.; Finch, R.; Crook, N.; O’Reilly, D.R.; Winstanley, D. The complete sequence of the Cydia pomonella granulovirus genome. J. Gen. Virol. 2001, 82, 2531–2547. [Google Scholar] [PubMed]
- Fritsch, E.; Undorf-Span, K.; Kienle, J.; Zebitz, C.P.W.; Huber, J. Codling moth granulovirus: Variations in the susceptibility of local codling moth populations. Nachrichtenblatt des Deutschen Pflanzenschutzdienstes 2005, 57, 29–34. [Google Scholar]
- Sauphanor, B.; Berling, M.; Toubon, J.F.; Reyes, M.; Delnatte, J.; Allemoz, P. Carpocapse des pommes: Cas de résistance au virus de la granulose en vergers biologiques. Phytoma Def. Veg. 2006, 590, 24–27. [Google Scholar]
- Schmitt, A.; Bisutti, I.L.; Ladurner, E.; Benuzzi, M.; Sauphanor, B.; Kienzle, J.; Zingg, D.; Undorf-Spahn, K.; Fritsch, E.; Huber, J.; et al. The occurrence and distribution of resistance of codling moth to Cydia pomonella granulovirus in Europe. J. Appl. Entomol. 2013, 137, 641–649. [Google Scholar] [CrossRef]
- Berling, M.; Blachere-Lopez, C.; Soubabere, O.; Lery, X.; Bonhomme, A.; Sauphanor, B.; Lopez-Ferber, M. Cydia pomonella granulovirus genotypes overcome virus resistance in the codling moth and improve virus efficiency by selection against resistant hosts. Appl. Environ. Microb. 2009, 75, 925–930. [Google Scholar] [CrossRef]
- Finney, J.R. Probit Analisis, 3rd ed.; Cambridge University Press: Cambridge, UK, 1971. [Google Scholar]
- POLO-PC software; Version 2.0; a user’s manual for Probit or Logit analysis; LeOra Software: Berkeley, CA, USA, 1987.
- Gebhardt, M.M.; Eberle, K.E.; Radtke, P.; Jehle, J.A. Baculovirus resistance in codling moth is virus isolate-dependent and the consequence of a mutation in viral gene pe38. Proc. Natl. Acad. Sci. USA 2014, 111, 15711–15716. [Google Scholar] [CrossRef] [PubMed]
- Eberle, K.E.; Wennmann, J.T.; Kleespies, R.G.; Jehle, J.A. Basic techniques in insect virology. In Manual of Techniques in Invertebrate Pathology, 2nd ed.; Lacey, L.A., Ed.; Academic Press: Walthman, Ma, USA, 2012. [Google Scholar]
- Reiser, M.; Groner, A.; Sander, E. Cryptophlebia-leucotreta (Lep, Tortricidae) a promising alternate host for mass-production of CpGV for biological pest-control. Z. Pflanzenk. Pflanzen. 1993, 100, 586–598. [Google Scholar]
- Undorf-Spahn, K.; Fritsch, E.; Huber, J.; Kienzle, J.; Zebitz, C.P.W.; Jehle, J.A. High stability and no fitness costs of the resistance of codling moth to Cydia pomonella granulovirus (CpGV-M). J. Invertebr. Pathol. 2012, 111, 136–142. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Graillot, B.; Berling, M.; Blachere-López, C.; Siegwart, M.; Besse, S.; López-Ferber, M. Progressive Adaptation of a CpGV Isolate to Codling Moth Populations Resistant to CpGV-M. Viruses 2014, 6, 5135-5144. https://doi.org/10.3390/v6125135
Graillot B, Berling M, Blachere-López C, Siegwart M, Besse S, López-Ferber M. Progressive Adaptation of a CpGV Isolate to Codling Moth Populations Resistant to CpGV-M. Viruses. 2014; 6(12):5135-5144. https://doi.org/10.3390/v6125135
Chicago/Turabian StyleGraillot, Benoît, Marie Berling, Christine Blachere-López, Myriam Siegwart, Samantha Besse, and Miguel López-Ferber. 2014. "Progressive Adaptation of a CpGV Isolate to Codling Moth Populations Resistant to CpGV-M" Viruses 6, no. 12: 5135-5144. https://doi.org/10.3390/v6125135
APA StyleGraillot, B., Berling, M., Blachere-López, C., Siegwart, M., Besse, S., & López-Ferber, M. (2014). Progressive Adaptation of a CpGV Isolate to Codling Moth Populations Resistant to CpGV-M. Viruses, 6(12), 5135-5144. https://doi.org/10.3390/v6125135