Poxviruses in Bats … so What?

Abstract
:1. Background and Significance

2. The Natural History of Poxvirus Infection in Bats

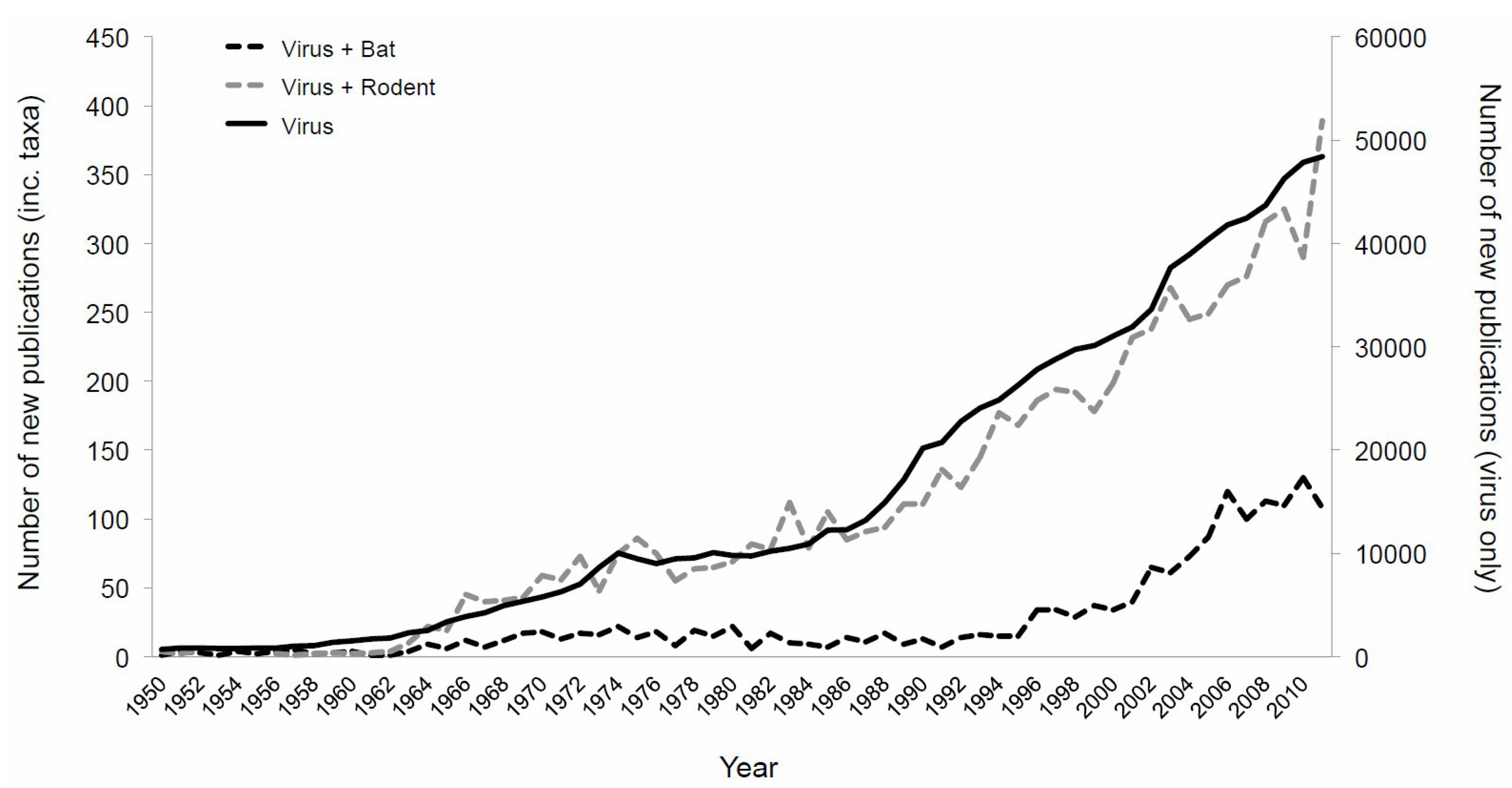{kind=link}
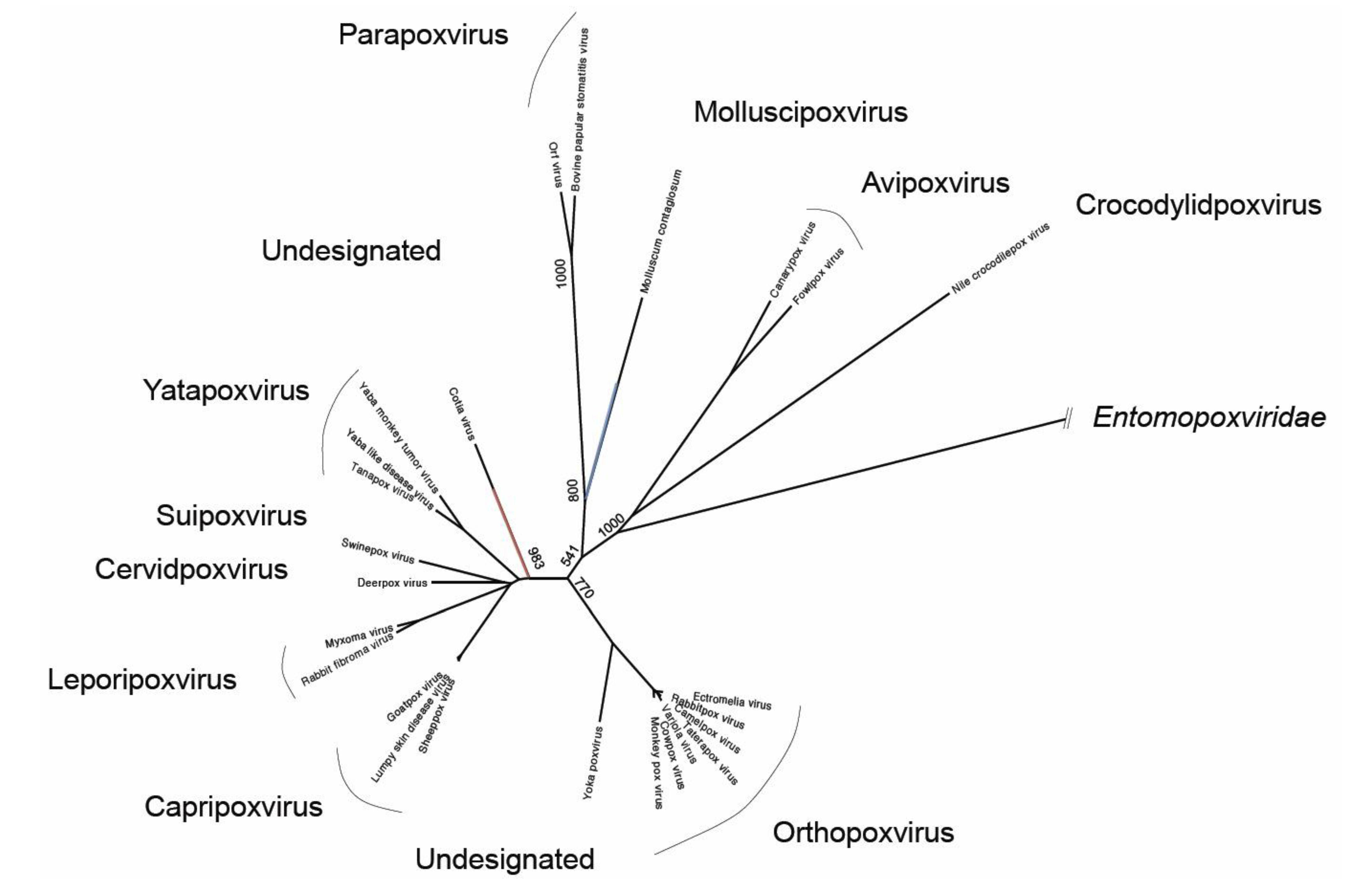{kind=link}
| Bat Species | Bat Family | Geographical Site | Clinical Signs | Evidence | Genetic Characterization | Virus Name | Reference |
|---|---|---|---|---|---|---|---|
| Eidolon helvum | Pteropodidae (Megabat) | West Africa | Apparently healthy | Sequence detection | Partial sequencing (12kb) | Eidolon helvum poxvirus 1 | [32] |
| Eptesicus fuscus | Vespertilionidae (Microbat) | USA | Tenosynovitis and osteoarthritis | EM A Isolated | Partial sequencing (19.5 kb) | Eptesipoxvirus | [35] |
| Miniopterus schreibersii | Vespertilionidae (Microbat) | Australia | Epidermal nodule | EM | NA B | NA | [38] |
2.1. Molecular Detection through Metagenomics

2.2. Viral Isolation and Clinical Infections
2.3. Interrelationships of Bat Poxviruses
3. The Zoonotic Potential of Bat Poxviruses
4. Future Directions
4.1. Coevolution of Bats and Poxviruses
4.2. Cross Species Infections
4.3. Mechanisms of Poxvirus Host Tropism
4.4. Bat Immunology and Virology
5. Concluding Remarks
Supplementary Files
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- McFadden, G. Poxvirus tropism. Nat. Rev. Microbiol. 2005, 3, 201–213. [Google Scholar] [CrossRef]
- Nollens, H.H.; Jacobson, E.R.; Gulland, F.M.; Beusse, D.O.; Bossart, G.D.; Hernandez, J.A.; Klein, P.A.; Condit, R.C. Pathology and preliminary characterization of a parapoxvirus isolated from a California sea lion (Zalophus californianus). J. Wildl. Dis. 2006, 42, 23–32. [Google Scholar] [CrossRef]
- Bratke, K.A.; McLysaght, A.; Rothenburg, S. A survey of host range genes in poxvirus genomes. Infect. Genet. Evol. 2013, 14, 406–425. [Google Scholar] [CrossRef]
- Lewis-Jones, S. Zoonotic poxvirus infections in humans. Curr. Opin. Infect. Dis. 2004, 17, 81–89. [Google Scholar] [CrossRef]
- Di Giulio, D.B.; Eckburg, P.B. Human monkeypox: An emerging zoonosis. Lancet. Infect. Dis. 2004, 4, 15–25. [Google Scholar] [CrossRef]
- Carroll, B.; Russell, P.; Gurnell, J.; Nettleton, P.; Sainsbury, A.W. Epidemics of squirrelpox virus disease in red squirrels (Sciurus vulgaris): Temporal and serological findings. Epidemiol. Infect. 2009, 137, 257–265. [Google Scholar] [CrossRef]
- Shchelkunov, S.N. An increasing danger of zoonotic orthopoxvirus infections. PLoS Pathog. 2013, 9, e1003756. [Google Scholar] [CrossRef]
- Haller, S.L.; Peng, C.; McFadden, G.; Rothenburg, S. Poxviruses and the evolution of host range and virulence. Infect. Genet. Evol. 2013, 21C, 15–40. [Google Scholar]
- Moss, B. Poxvirus entry and membrane fusion. Virology 2006, 344, 48–54. [Google Scholar] [CrossRef]
- Werden, S.J.; Rahman, M.M.; McFadden, G. Poxvirus host range genes. Adv. Virus Res. 2008, 71, 135–171. [Google Scholar] [CrossRef]
- Spehner, D.; Gillard, S.; Drillien, R.; Kirn, A. A cowpox virus gene required for multiplication in Chinese hamster ovary cells. J. Virol. 1988, 62, 1297–1304. [Google Scholar]
- Kerr, P.J.; Rogers, M.B.; Fitch, A.; Depasse, J.V.; Cattadori, I.M.; Hudson, P.J.; Tscharke, D.C.; Holmes, E.C.; Ghedin, E. Comparative analysis of the complete genome sequence of the California MSW strain of myxoma virus reveals potential host adaptations. J. Virol. 2013, 87, 12080–12089. [Google Scholar] [CrossRef]
- Hueffer, K.; Parrish, C.R. Parvovirus host range, cell tropism and evolution. Curr. Opin. Microbiol. 2003, 6, 392–398. [Google Scholar] [CrossRef]
- Imai, M.; Watanabe, T.; Hatta, M.; Das, S.C.; Ozawa, M.; Shinya, K.; Zhong, G.; Hanson, A.; Katsura, H.; Watanabe, S.; et al. Experimental adaptation of an influenza H5 HA confers respiratory droplet transmission to a reassortant H5 HA/H1N1 virus in ferrets. Nature 2012, 486, 420–428. [Google Scholar]
- Calisher, C.H.; Childs, J.E.; Field, H.E.; Holmes, K.V.; Schountz, T. Bats: Important reservoir hosts of emerging viruses. Clin. Microbiol. Rev. 2006, 19, 531–545. [Google Scholar] [CrossRef]
- Teeling, E.C.; Springer, M.S.; Madsen, O.; Bates, P.; O’Brien, J.S.; Murphy, W.J. A molecular phylogeny for bats illuminates biogeography and the fossil record. Science 2005, 307, 580–584. [Google Scholar] [CrossRef]
- Yang, Z. Likelihood and Bayes estimation of ancestral population sizes in hominoids using data from multiple loci. Genetics 2002, 162, 1811–1823. [Google Scholar]
- Springer, M.S.; Teeling, E.C.; Madsen, O.; Stanhope, M.J.; de Jong, W.W. Integrated fossil and molecular data reconstruct bat echolocation. Proc. Natl. Acad. Sci. USA 2001, 98, 6241–6246. [Google Scholar]
- Li, W.; Shi, Z.; Yu, M.; Ren, W.; Smith, C.; Epstein, J.H.; Wang, H.; Crameri, G.; Hu, Z.; Zhang, H.; et al. Bats are natural reservoirs of SARS-like coronaviruses. Science 2005, 310, 676–679. [Google Scholar] [CrossRef]
- Ge, X.Y.; Li, J.L.; Yang, X.L.; Chmura, A.A.; Zhu, G.; Epstein, J.H.; Mazet, J.K.; Hu, B.; Zhang, W.; Peng, C.; et al. Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor. Nature 2013, 503, 535–538. [Google Scholar] [CrossRef]
- Halpin, K.; Young, P.L.; Field, H.E.; MacKenzie, J.S. Isolation of Hendra virus from pteropid bats: a natural reservoir of Hendra virus. J. Gen. Virol. 2000, 81, 1927–1932. [Google Scholar]
- Chua, K.B.; Hooi, P.S., L.K.C.; Wee, K.F.; Khong, J.H.; Chua, B.H.; Chan, Y.P.; Lim, M.E.; Lam Sai Kit, K. Isolation of Nipah virus from Malaysian Island flying-foxes. Microbes Infect. 2002, 4, 145–151. [Google Scholar] [CrossRef]
- Towner, J.S.; Amman, B.R.; Sealy, T.K.; Carroll, S.A.; Comer, J.A.; Kemp, A.; Swanepoel, R.; Paddock, C.D.; Balinandi, S.; Khristova, M.L.; et al. Isolation of genetically diverse Marburg viruses from Egyptian fruit bats. PLoS Pathog. 2009, 5, e1000536. [Google Scholar] [CrossRef]
- Leroy, E.M.; Kumulungui, B.; Pourrut, X.; Rouquet, P.; Hassanin, A.; Yaba, P.; Delicat, A.; Paweska, J.T.; Gonzalez, J.P.; Swanepoel, R. Fruit bats as reservoirs of Ebola virus. Nature 2005, 438, 575–576. [Google Scholar] [CrossRef]
- Rupprecht, C.E.; Turmelle, A.; Kuzmin, I.V. A perspective on lyssavirus emergence and perpetuation. Curr. Opin. Virol. 2011, 1, 662–670. [Google Scholar] [CrossRef]
- Drexler, J.F.; Geipel, A.; Konig, A.; Corman, V.M.; van Riel, D.; Leijten, L.M.; Bremer, C.M.; Rasche, A.; Cottontail, V.M.; Maganga, G.D.; et al. Bats carry pathogenic hepadnaviruses antigenically related to hepatitis B virus and capable of infecting human hepatocytes. Proc. Natl. Acad. Sci. USA 2013, 110, 16151–16156. [Google Scholar] [CrossRef]
- Tong, S.; Li, Y.; Rivailler, P.; Conrardy, C.; Castillo, D.A.; Chen, L.M.; Recuenco, S.; Ellison, J.A.; Davis, C.T.; York, I.A.; et al. A distinct lineage of influenza A virus from bats. Proc. Natl. Acad. Sci. USA 2012, 109, 4269–4274. [Google Scholar] [CrossRef]
- Wang, L.F.; Walker, P.J.; Poon, L.L. Mass extinctions, biodiversity and mitochondrial function: Are bats “special” as reservoirs for emerging viruses? Curr. Opin. Virol. 2011, 1, 649–657. [Google Scholar] [CrossRef]
- Baker, M.L.; Schountz, T.; Wang, L.F. Antiviral immune responses of bats: A review. Zoonoses Public Health 2013, 60, 104–116. [Google Scholar] [CrossRef]
- Luis, A.D.; Hayman, D.T.; O’Shea, T.J.; Cryan, P.M.; Gilbert, A.T.; Pulliam, J.R.; Mills, J.N.; Timonin, M.E.; Willis, C.K.; Cunningham, A.A.; et al. A comparison of bats and rodents as reservoirs of zoonotic viruses: Are bats special? Proc. Biol. Sci. 2013, 280. [Google Scholar] [CrossRef]
- Anthony, S.J.; Epstein, J.H.; Murray, K.A.; Navarrete-Macias, I.; Zambrana-Torrelio, C.M.; Solovyov, A.; Ojeda-Flores, R.; Arrigo, N.C.; Islam, A.; Khan, S.A.; et al. A strategy to estimate unknown viral diversity in mammals. mBio 2013, 4, e00598–e00513. [Google Scholar]
- Baker, K.S.; Leggett, R.M.; Bexfield, N.H.; Alston, M.; Daly, G.; Todd, S.; Tachedjian, M.; Holmes, C.E.; Crameri, S.; Wang, L.F.; et al. Metagenomic study of the viruses of African straw-coloured fruit bats: Detection of a chiropteran poxvirus and isolation of a novel adenovirus. Virology 2013, 441, 95–106. [Google Scholar] [CrossRef]
- Li, L.; Victoria, J.G.; Wang, C; Jones, M.; Fellers, G.M.; Kunz, T.H.; Delwart, E. Bat guano virome: Predominance of dietary viruses from insects and plants plus novel mammalian viruses. J. Virol. 2010, 84, 6955–6965. [Google Scholar]
- He, B.; Li, Z.; Yang, F.; Zheng, J.; Feng, Y.; Guo, H.; Li, Y.; Wang, Y.; Su, N.; Zhang, F.; et al. Virome profiling of bats from Myanmar by metagenomic analysis of tissue samples reveals more novel Mammalian viruses. PLoS One 2013, 8, e61950. [Google Scholar]
- Emerson, G.L.; Nordhausen, R.; Garner, M.M.; Huckabee, J.R.; Johnson, S.; Wohrle, R.D.; Davidson, W.B.; Wilkins, K.; Li, Y.; Doty, J.B.; et al. Novel poxvirus in big brown bats, northwestern United States. Emerg. Infect. Dis. 2013, 19, 1002–1004. [Google Scholar]
- Lopesode, S.; Lacerda, J.P.; Fonseca, I.E.; Castro, D.P.; Forattini, O.P.; Rabello, E.X. Cotia Virus: A New Agent Isolated from Sentinel Mice in S Ao Paulo, Brazil. Am. J. Trop. Med. Hyg. 1965, 14, 156–157. [Google Scholar]
- Afonso, P.P.; Silva, P.M.; Schnellrath, L.C.; Jesus, D.M.; Hu, J.; Yang, Y.; Renne, R.; Attias, M.; Condit, R.C.; Moussatche, N.; et al. Biological characterization and next-generation genome sequencing of the unclassified Cotia virus SPAn232 (Poxviridae). J. Virol. 2012, 86, 5039–5054. [Google Scholar] [CrossRef]
- McLelland, D.J.; Reardon, T.; Bourne, S.; Dickason, C.; Kessell, A.; Boardman, W. Outbreak of Skin Nodules Associated with Riouxgolvania beveridgei (Nematoda: Muspiceida) in the Southern Bentwing Bat (Miniopterus schreibersii bassanii), South Australia. J. Wildl. Dis. 2013, 49, 1009–1013. [Google Scholar] [CrossRef]
- Vogelnest, L.; Stewart, S.; Sangster, C. Poxvirus infection outbreak in common ringtails (Pseudocheirus peregrinus). Aust. Vet. J. 2012, 90, 143–145. [Google Scholar] [CrossRef]
- Afonso, C.L.; Tulman, E.R.; Delhon, G.; Lu, Z.; Viljoen, G.J.; Wallace, D.B.; Kutish, G.F.; Rock, D.L. Genome of crocodilepox virus. J. Virol. 2006, 80, 4978–4991. [Google Scholar] [CrossRef]
- Fox, R.; Thiemann, A.; Everest, D.; Steinbach, F.; Dastjerdi, A.; Finnegan, C. Molluscum contagiosum in two donkeys. Vet. Rec. 2012, 170, 649. [Google Scholar]
- Thompson, C.H.; Yager, J.A.; Van Rensburg, I.B. Close relationship between equine and human molluscum contagiosum virus demonstrated by in situ hybridisation. Res. Vet. Sci. 1998, 64, 157–161. [Google Scholar] [CrossRef]
- Bagnall, B.G.; Wilson, G.R. Molluscum contagiosum in a red kangaroo. Australas. J. Dermatol. 1974, 15, 115–120. [Google Scholar] [CrossRef]
- Emerson, G.L.; Li, Y.; Frace, M.A.; Olsen-Rasmussen, M.A.; Khristova, M.L.; Govil, D.; Sammons, S.A.; Regnery, R.L.; Karem, K.L.; Damon, I.K.; et al. The phylogenetics and ecology of the orthopoxviruses endemic to North America. PLoS One 2009, 4, e7666. [Google Scholar] [CrossRef]
- Alexander, A.D.; Flyger, V.; Herman, Y.F.; McConnell, S.J.; Rothstein, N.; Yager, R.H. Survey of wild mammals in a Chesapeake Bay area for selected zoonoses. J. Wildl. Dis. 1972, 8, 119–126. [Google Scholar] [CrossRef]
- Babkin, I.V.; Shchelkunov, S.N. The time scale in poxvirus evolution. Mol. Biol. (Mosk.) 2006, 40, 20–24. [Google Scholar] [CrossRef]
- Tong, S.; Zhu, X.; Li, Y.; Shi, M.; Zhang, J.; Bourgeois, M.; Yang, H.; Chen, X.; Recuenco, S.; Gomez, J.; et al. New world bats harbor diverse influenza A viruses. PLoS Pathog. 2013, 9, e1003657. [Google Scholar] [CrossRef]
- Chu, D.K.; Poon, L.L.; Guan, Y.; Peiris, J.S. Novel astroviruses in insectivorous bats. J. Virol. 2008, 82, 9107–9114. [Google Scholar] [CrossRef]
- Ge, X.; Li, J.; Peng, C.; Wu, L.; Yang, X.; Wu, Y.; Zhang, Y.; Shi, Z. Genetic diversity of novel circular ssDNA viruses in bats in China. J. Gen. Virol. 2011, 92, 2646–2653. [Google Scholar] [CrossRef]
- Li, Y.; Ge, X.; Zhang, H.; Zhou, P.; Zhu, Y.; Zhang, Y.; Yuan, J.; Wang, L.F.; Shi, Z. Host range, prevalence, and genetic diversity of adenoviruses in bats. J. Virol. 2010, 84, 3889–3897. [Google Scholar] [CrossRef]
- Towner, J.S.; Pourrut, X.; Albarino, C.G.; Nkogue, C.N.; Bird, B.H.; Grard, G.; Ksiazek, T.G.; Gonzalez, J.P.; Nichol, S.T.; Leroy, E.M. Marburg virus infection detected in a common African bat. PLoS One 2007, 2, e764. [Google Scholar] [CrossRef]
- Vijaykrishna, D.; Smith, G.J.; Zhang, J.X.; Peiris, J.S.; Chen, H.; Guan, Y. Evolutionary insights into the ecology of coronaviruses. J. Virol. 2007, 81, 4012–4020. [Google Scholar] [CrossRef]
- Bourhy, H.; Kissi, B.; Tordo, N. Molecular diversity of the Lyssavirus genus. Virology 1993, 194, 70–81. [Google Scholar] [CrossRef]
- Jones, G.; Teeling, E.C. The evolution of echolocation in bats. Trends Ecol. Evol. 2006, 21, 149–156. [Google Scholar] [CrossRef]
- Iyer, L.M.; Balaji, S.; Koonin, E.V.; Aravind, L. Evolutionary genomics of nucleo-cytoplasmic large DNA viruses. Virus Res. 2006, 117, 156–184. [Google Scholar] [CrossRef]
- Hughes, A.L.; Friedman, R. Poxvirus genome evolution by gene gain and loss. Mol. Phylogenet. Evol. 2005, 35, 186–195. [Google Scholar] [CrossRef]
- Fenner, F. Adventures with poxviruses of vertebrates. FEMS Microbiol. Rev. 2000, 24, 123–133. [Google Scholar] [CrossRef]
- Dzal, Y.; McGuire, L.P.; Veselka, N.; Fenton, M.B. Going, going, gone: The impact of white-nose syndrome on the summer activity of the little brown bat (Myotis lucifugus.). Biol. Lett. 2011, 7, 392–394. [Google Scholar] [CrossRef]
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef]
- Streicker, D.G.; Turmelle, A.S.; Vonhof, M.J.; Kuzmin, I.V.; McCracken, G.F.; Rupprecht, C.E. Host phylogeny constrains cross-species emergence and establishment of rabies virus in bats. Science 2010, 329, 676–679. [Google Scholar] [CrossRef]
- Streicker, D.G.; Lemey, P.; Velasco-Villa, A.; Rupprecht, C.E. 2012. Rates of viral evolution are linked to host geography in bat rabies. PLoS Pathog. 2012, 8, e1002720. [Google Scholar] [CrossRef]
- Jordan, I.; Horn, D.; Oehmke, S.; Leendertz, F.H.; Sandig, V. Cell lines from the Egyptian fruit bat are permissive for modified vaccinia Ankara. Virus Res. 2009, 145, 54–62. [Google Scholar] [CrossRef]
- Crameri, G.; Todd, S.; Grimley, S.; McEachern, J.A.; Marsh, G.A.; Smith, C.; Tachedjian, M.; de Jong, C.; Virtue, E.R.; Yu, M.; et al. Establishment, immortalisation and characterisation of pteropid bat cell lines. PLoS One 2009, 4, e8266. [Google Scholar] [CrossRef]
- Jordan, I.; Munster, V.J.; Sandig, V. Authentication of the R06E fruit bat cell line. Viruses 2012, 4, 889–900. [Google Scholar] [CrossRef]
- Mourya, D.T.; Lakra, R.J.; Yadav, P.D.; Tyagi, P.; Raut, C.G.; Shete, A.M.; Singh, D.K. Establishment of cell line from embryonic tissue of Pipistrellus ceylonicus bat species from India & its susceptibility to different viruses. Indian J. Med. Res. 2013, 138, 224–231. [Google Scholar]
- Biesold, S.E.; Ritz, D.; Gloza-Rausch, F.; Wollny, R.; Drexler, J.F.; Corman, V.M.; Kalko, E.K.; Oppong, S.; Drosten, C.; Muller, M.A. Type I interferon reaction to viral infection in interferon-competent, immortalized cell lines from the African fruit bat Eidolon helvum. PLoS One 2011, 6, e28131. [Google Scholar] [CrossRef]
- Sheppard, S.K.; Didelot, X.; Meric, G.; Torralbo, A.; Jolley, K.A.; Kelly, D.J.; Bentley, S.D.; Maiden, M.C.; Parkhill, J.; Falush, D. Genome-wide association study identifies vitamin B5 biosynthesis as a host specificity factor in Campylobacter. Proc. Natl Acad. Sci. USA 2013, 110, 11923–11927. [Google Scholar] [CrossRef]
- Zhang, G.; Cowled, C.; Shi, Z.; Huang, Z.; Bishop-Lilly, K.A.; Fang, X.; Wynne, J.W.; Xiong, Z.; Baker, M.L.; Zhao, W. Comparative analysis of bat genomes provides insight into the evolution of flight and immunity. Science 2013, 339, 456–460. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Baker, K.S.; Murcia, P.R. Poxviruses in Bats … so What? Viruses 2014, 6, 1564-1577. https://doi.org/10.3390/v6041564
Baker KS, Murcia PR. Poxviruses in Bats … so What? Viruses. 2014; 6(4):1564-1577. https://doi.org/10.3390/v6041564
Chicago/Turabian StyleBaker, Kate S., and Pablo R. Murcia. 2014. "Poxviruses in Bats … so What?" Viruses 6, no. 4: 1564-1577. https://doi.org/10.3390/v6041564
APA StyleBaker, K. S., & Murcia, P. R. (2014). Poxviruses in Bats … so What? Viruses, 6(4), 1564-1577. https://doi.org/10.3390/v6041564