Infection with Non-Lethal West Nile Virus Eg101 Strain Induces Immunity that Protects Mice against the Lethal West Nile Virus NY99 Strain



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. WNV Infection Experiment
2.3. WNV Plaque Assay
2.4. Measurement of WNV-Specific Antibodies
2.5. Plaque Reduction Neutralization Test (PRNT)
2.6. Challenge Experiment
2.7. Statistical Analysis
3. Results
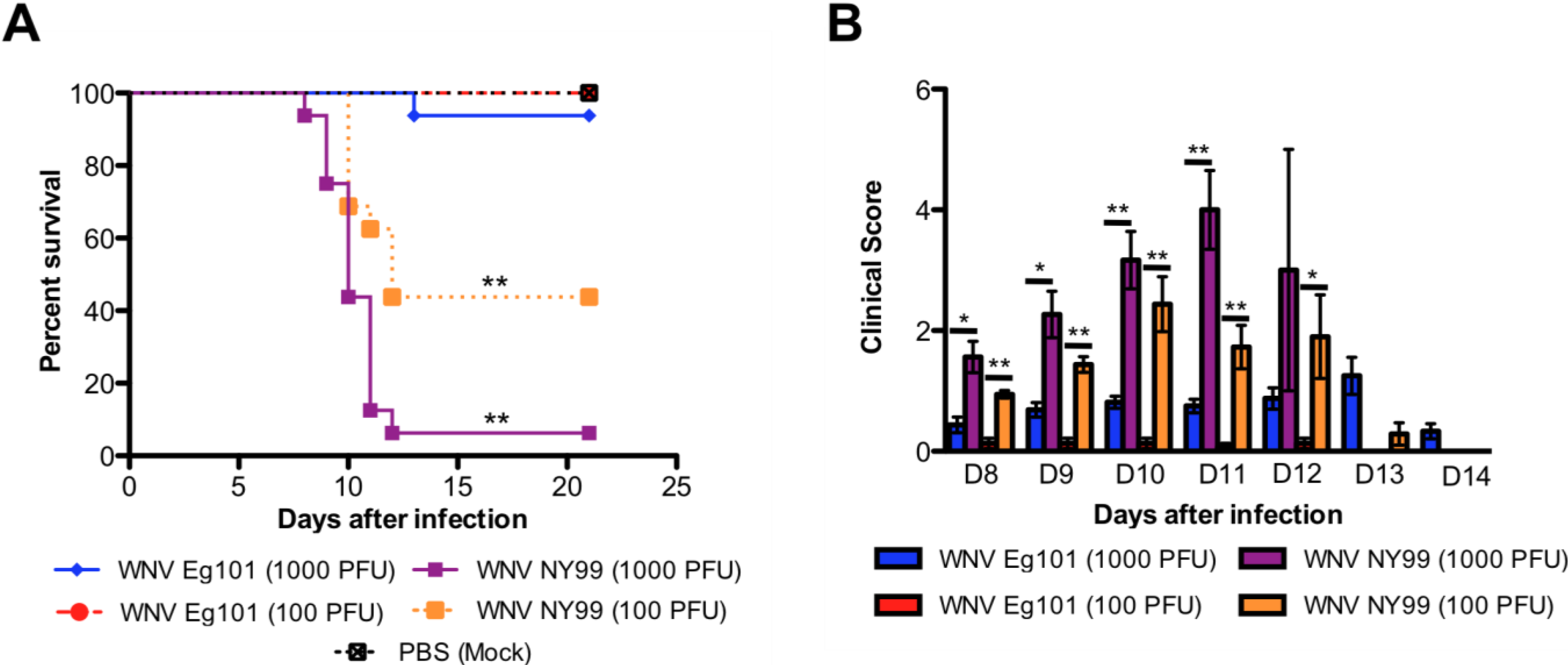
3.1. Virulence of WNV Eg101 and WNV NY99 Strains in Eight-Week-Old C57BL/6J Mice

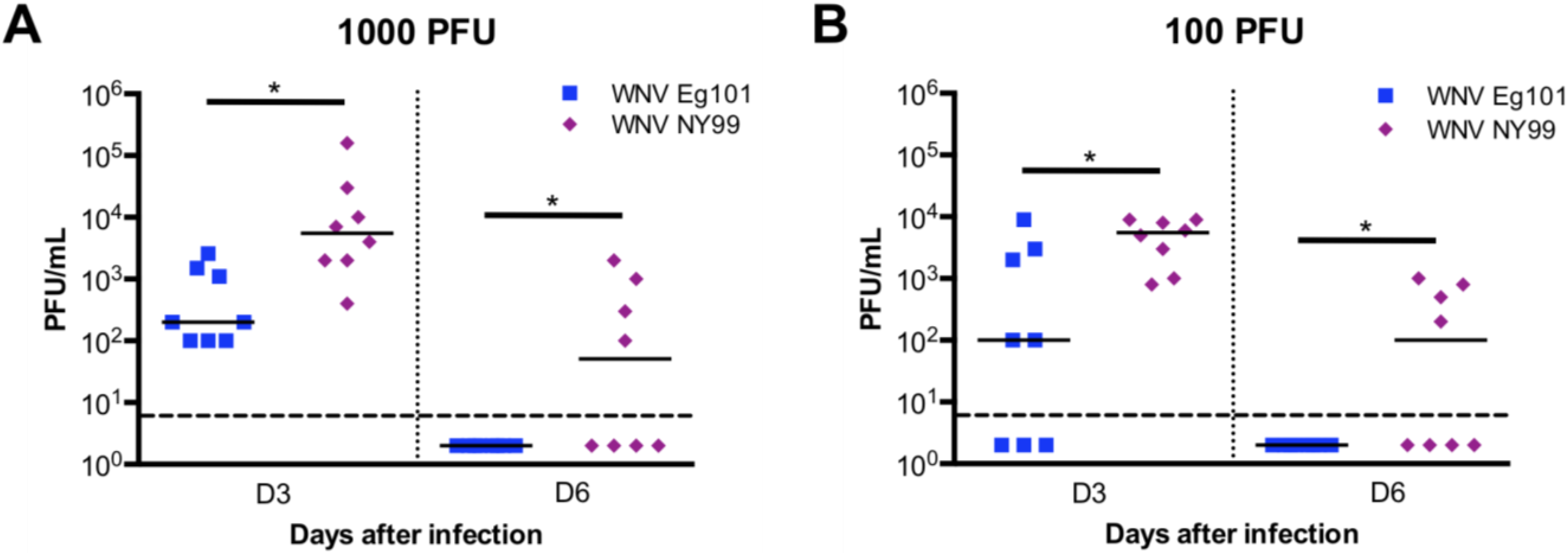
3.2. WNV Titers in the Serum of WNV Eg101 and WNV NY99 Infected Mice
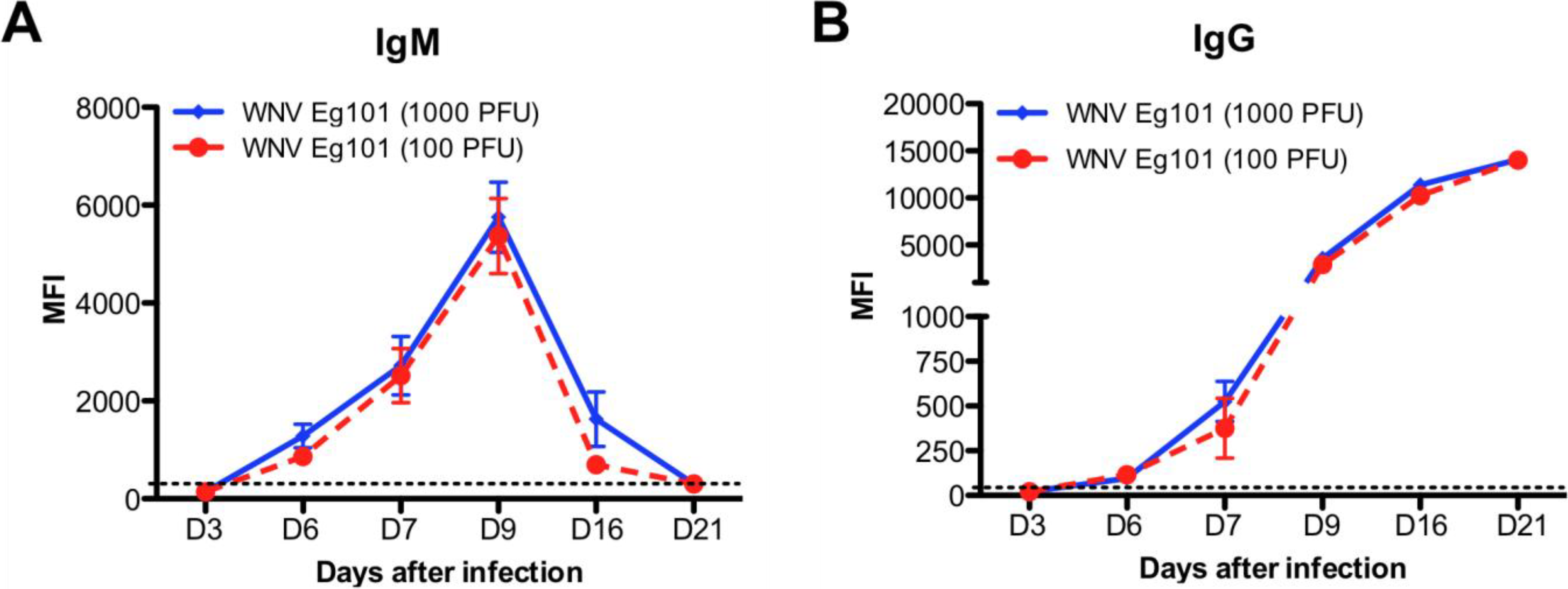
3.3. Induction of WNV-Specific Antibodies Responses in WNV Eg101 Infected Mice


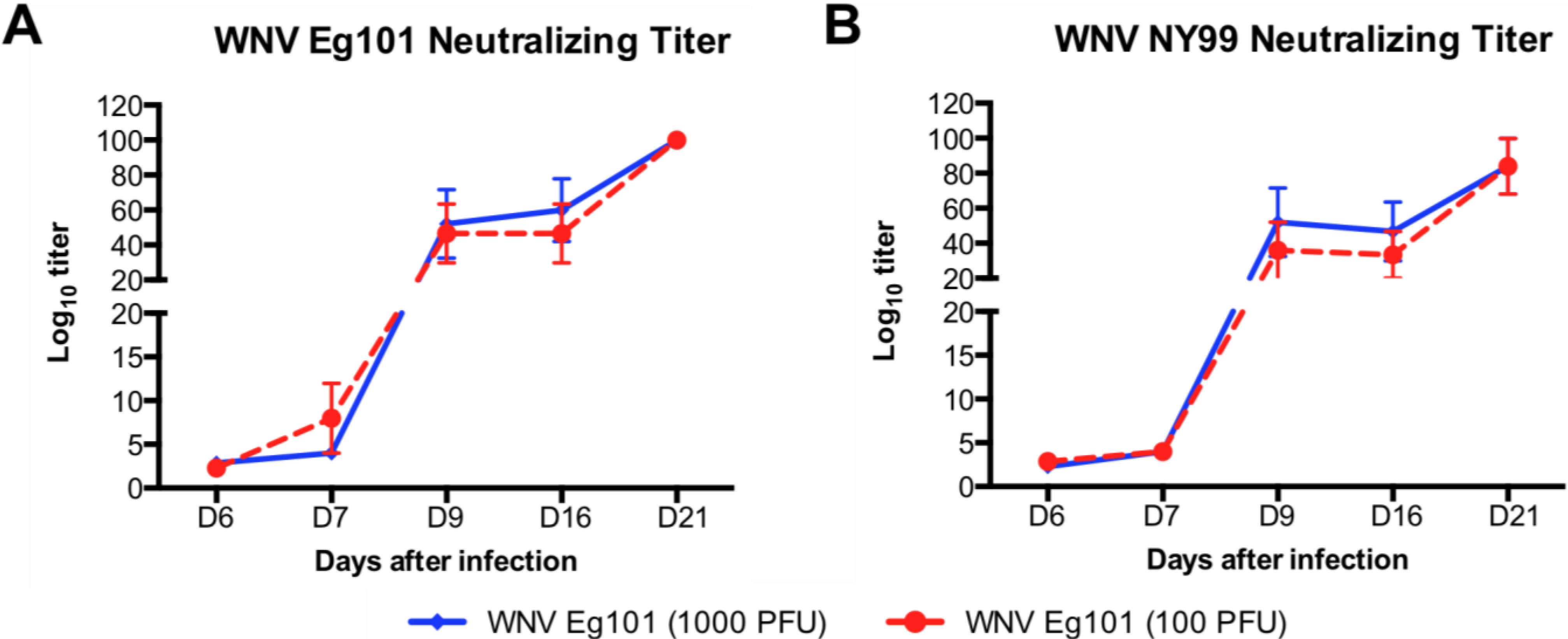
3.4. WNV Eg101 Infection Induces Cross-Reactive Antibodies that Neutralize the WNV NY99 Strain

3.5. Protection of WNV Eg101 Immunized Mice against Subcutaneous or Intracranial WNV NY99 Challenge

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subcutaneous challenge with WNV NY99 (1000 PFU)# | Intracranial challenge with WNV NY99 (1000 PFU)# | ||||
|---|---|---|---|---|---|
| Mouse group | % Morbidity (sick/challenged) | % Mortality (dead/challenged) | % Morbidity (sick/challenged) | % Mortality (dead/challenged) | |
| PBS | 100 (16/16) | 94 (15/16) | 100 (16/16) | 100 (16/16) | |
| WNV Eg101 (1000 PFU) | 0 (0/16) | 0 (0/16) | 0 (0/16) | 0 (0/16) | |
| WNV Eg101 (100 PFU) | 0 (0/16) | 0 (0/16) | 0 (0/16) | 0 (0/16) | |
4. Discussion
4.1. Eg101 Strain Is Non-Pathogenic Compared to NY99 Strain of WNV
4.2. WNV Eg101 Infection in Mice Induced Strong WNV-Specific Antibodies Responses
4.3. WNV Eg101 Immunized Mice Were Completely Protected against Pathogenic WNV NY99
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Hayes, E.B.; Gubler, D.J. West Nile virus: Epidemiology and clinical features of an emerging epidemic in the United States. Annu. Rev. Med. 2006, 57, 181–194. [Google Scholar] [CrossRef]
- Smithburn, K.; Hughes, T.; Burke, A.; Paul, J. A neurotropic virus isolated from the blood of a native of Uganda. Am. J. Trop. Med. Hyg. 1940, 20, 471–492. [Google Scholar]
- Hubalek, Z.; Halouzka, J. West Nile fever—A reemerging mosquito-borne viral disease in Europe. Emerg. Infect. Dis. 1999, 5, 643–650. [Google Scholar] [CrossRef]
- Beasley, D.W.; Barrett, A.D.; Tesh, R.B. Resurgence of West Nile neurologic disease in the United States in 2012: What happened? What needs to be done? Antivir. Res. 2013, 99, 1–5. [Google Scholar] [CrossRef]
- Donadieu, E.; Bahuon, C.; Lowenski, S.; Zientara, S.; Coulpier, M.; Lecollinet, S. Differential virulence and pathogenesis of West Nile viruses. Viruses 2013, 5, 2856–2880. [Google Scholar]
- Murray, K.O.; Mertens, E.; Despres, P. West Nile virus and its emergence in the United States of America. Vet. Res. 2010, 41, 67. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Roehrig, J.T.; Deubel, V.; Smith, J.; Parker, M.; Steele, K.; Crise, B.; Volpe, K.E.; Crabtree, M.B.; Scherret, J.H.; et al. Origin of the West Nile virus responsible for an outbreak of encephalitis in the northeastern United States. Science 1999, 286, 2333–2337. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Ebel, G.D.; Deubel, V.; Kerst, A.J.; Murri, S.; Meyer, R.; Bowen, M.; McKinney, N.; Morrill, W.E.; Crabtree, M.B.; et al. Complete genome sequences and phylogenetic analysis of West Nile virus strains isolated from the United States, Europe, and the Middle East. Virology 2002, 298, 96–105. [Google Scholar] [CrossRef]
- Petersen, L.R.; Carson, P.J.; Biggerstaff, B.J.; Custer, B.; Borchardt, S.M.; Busch, M.P. Estimated cumulative incidence of West Nile virus infection in US adults, 1999–2010. Epidemiol. Infect. 2013, 141, 591–595. [Google Scholar] [CrossRef]
- Rizzo, C.; Salcuni, P.; Nicoletti, L.; Ciufolini, M.G.; Russo, F.; Masala, R.; Frongia, O.; Finarelli, A.C.; Gramegna, M.; Gallo, L.; et al. Epidemiological surveillance of West Nile neuroinvasive diseases in Italy, 2008 to 2011. Euro. Surveill. 2012, 17, 20172. [Google Scholar]
- Papa, A. West Nile virus infections in Greece: An update. Expert Rev. Anti. Infect. Ther. 2012, 10, 743–750. [Google Scholar] [CrossRef]
- Calistri, P.; Giovannini, A.; Hubalek, Z.; Ionescu, A.; Monaco, F.; Savini, G.; Lelli, R. Epidemiology of west nile in europe and in the mediterranean basin. Open Virol. J. 2010, 4, 29–37. [Google Scholar]
- Diamond, M.S.; Shrestha, B.; Marri, A.; Mahan, D.; Engle, M. B cells and antibody play critical roles in the immediate defense of disseminated infection by West Nile encephalitis virus. J. Virol. 2003, 77, 2578–2586. [Google Scholar] [CrossRef]
- Diamond, M.S.; Sitati, E.M.; Friend, L.D.; Higgs, S.; Shrestha, B.; Engle, M. A critical role for induced IgM in the protection against West Nile virus infection. J. Exp. Med. 2003, 198, 1853–1862. [Google Scholar] [CrossRef]
- Diamond, M.S.; Pierson, T.C.; Fremont, D.H. The structural immunology of antibody protection against West Nile virus. Immunol. Rev. 2008, 225, 212–225. [Google Scholar] [CrossRef]
- Ben-Nathan, D.; Lustig, S.; Tam, G.; Robinzon, S.; Segal, S.; Rager-Zisman, B. Prophylactic and therapeutic efficacy of human intravenous immunoglobulin in treating West Nile virus infection in mice. J. Infect. Dis. 2003, 188, 5–12. [Google Scholar]
- Shrestha, B.; Diamond, M.S. Role of CD8+ T cells in control of West Nile virus infection. J. Virol. 2004, 78, 8312–8321. [Google Scholar] [CrossRef]
- Sitati, E.M.; Diamond, M.S. CD4+ T-cell responses are required for clearance of West Nile virus from the central nervous system. J. Virol. 2006, 80, 12060–12069. [Google Scholar] [CrossRef]
- Shrestha, B.; Ng, T.; Chu, H.J.; Noll, M.; Diamond, M.S. The relative contribution of antibody and CD8+ T cells to vaccine immunity against West Nile encephalitis virus. Vaccine 2008, 26, 2020–2033. [Google Scholar] [CrossRef]
- Yamshchikov, G.; Borisevich, V.; Seregin, A.; Chaporgina, E.; Mishina, M.; Mishin, V.; Kwok, C.W.; Yamshchikov, V. An attenuated West Nile prototype virus is highly immunogenic and protects against the deadly NY99 strain: a candidate for live WN vaccine development. Virology 2004, 330, 304–312. [Google Scholar] [CrossRef]
- Monath, T.P.; Liu, J.; Kanesa-Thasan, N.; Myers, G.A.; Nichols, R.; Deary, A.; McCarthy, K.; Johnson, C.; Ermak, T.; Shin, S.; et al. A live, attenuated recombinant West Nile virus vaccine. Proc. Natl. Acad. Sci. USA 2006, 103, 6694–6699. [Google Scholar] [CrossRef]
- Ng, T.; Hathaway, D.; Jennings, N.; Champ, D.; Chiang, Y.W.; Chu, H.J. Equine vaccine for West Nile virus. Dev. Biol. 2003, 114, 221–227. [Google Scholar]
- Melnick, J.L.; Paul, J.R.; Riordan, J.T.; Barnett, V.H.; Goldblum, N.; Zabin, E. Isolation from human sera in Egypt of a virus apparently identical to West Nile virus. Proc. Soc. Exp. Biol. Med. 1951, 77, 661–665. [Google Scholar] [CrossRef]
- Hurlbut, H.S.; Rizk, F.; Taylor, R.M.; Work, T.H. A study of the ecology of West Nile virus in Egypt. Am. J. Trop. Med. Hyg. 1956, 5, 579–620. [Google Scholar]
- Shirato, K.; Kimura, T.; Mizutani, T.; Kariwa, H.; Takashima, I. Different chemokine expression in lethal and non-lethal murine West Nile virus infection. J. Med. Virol. 2004, 74, 507–513. [Google Scholar] [CrossRef]
- Kumar, M.; Roe, K.; Nerurkar, P.V.; Namekar, M.; Orillo, B.; Verma, S.; Nerurkar, V.R. Impaired virus clearance, compromised immune response and increased mortality in type 2 diabetic mice infected with west nile virus. PLoS One 2012, 7, e44682. [Google Scholar]
- Kumar, M.; Roe, K.; Orillo, B.; Muruve, D.A.; Nerurkar, V.R.; Gale, M., Jr.; Verma, S. Inflammasome adaptor protein Apoptosis-associated speck-like protein containing CARD (ASC) is critical for the immune response and survival in west Nile virus encephalitis. J. Virol. 2013, 87, 3655–3667. [Google Scholar] [CrossRef]
- Roe, K.; Kumar, M.; Lum, S.; Orillo, B.; Nerurkar, V.R.; Verma, S. West Nile virus-induced disruption of the blood-brain barrier in mice is characterized by the degradation of the junctional complex proteins and increase in multiple matrix metalloproteinases. J. Gen. Virol. 2012, 93, 1193–1203. [Google Scholar] [CrossRef]
- Kumar, M.; Roe, K.; Nerurkar, P.V.; Orillo, B.; Thompson, K.; Verma, S.; Nerurkar, V.R. Reduced immune cell infiltration and increased pro-inflammatory mediators in the brain of type 2 diabetic mouse model infected with West Nile virus. J. Neuroinflammation 2014, 11, 80. [Google Scholar] [CrossRef]
- Kumar, M.; Nerurkar, V.R. Integrated analysis of microRNAs and their disease related targets in the brain of mice infected with West Nile virus. Virology 2014, 452–453, 143–151. [Google Scholar] [CrossRef]
- Kumar, M.; Verma, S.; Nerurkar, V.R. Pro-inflammatory cytokines derived from West Nile virus (WNV)-infected SK-N-SH cells mediate neuroinflammatory markers and neuronal death. J. Neuroinflammation 2010, 7, 73. [Google Scholar] [CrossRef]
- Namekar, M.; Kumar, M.; O'Connell, M.; Nerurkar, V.R. Effect of Serum Heat-Inactivation and Dilution on Detection of Anti-WNV Antibodies in Mice by West Nile Virus E-protein Microsphere Immunoassay. PLoS One 2012, 7, e45851. [Google Scholar]
- Lieberman, M.M.; Nerurkar, V.R.; Luo, H.; Cropp, B.; Carrion, R., Jr.; de la Garza, M.; Coller, B.A.; Clements, D.; Ogata, S.; Wong, T.; Martyak, T.; Weeks-Levy, C. Immunogenicity and protective efficacy of a recombinant subunit West Nile virus vaccine in rhesus monkeys. Clin. Vaccine Immunol. 2009, 16, 1332–1337. [Google Scholar] [CrossRef]
- GraphPad Prism, version 6.0e; GraphPad Software: La Jolla, CA, USA, 2014.
- Weiner, L.P.; Cole, G.A.; Nathanson, N. Experimental encephalitis following peripheral inoculation of West Nile virus in mice of different ages. J. Hyg. (Lond.) 1970, 68, 435–446. [Google Scholar] [CrossRef]
- Pinto, A.K.; Richner, J.M.; Poore, E.A.; Patil, P.P.; Amanna, I.J.; Slifka, M.K.; Diamond, M.S. A hydrogen peroxide-inactivated virus vaccine elicits humoral and cellular immunity and protects against lethal West Nile virus infection in aged mice. J. Virol. 2013, 87, 1926–1936. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kumar, M.; O'Connell, M.; Namekar, M.; Nerurkar, V.R. Infection with Non-Lethal West Nile Virus Eg101 Strain Induces Immunity that Protects Mice against the Lethal West Nile Virus NY99 Strain. Viruses 2014, 6, 2328-2339. https://doi.org/10.3390/v6062328
Kumar M, O'Connell M, Namekar M, Nerurkar VR. Infection with Non-Lethal West Nile Virus Eg101 Strain Induces Immunity that Protects Mice against the Lethal West Nile Virus NY99 Strain. Viruses. 2014; 6(6):2328-2339. https://doi.org/10.3390/v6062328
Chicago/Turabian StyleKumar, Mukesh, Maile O'Connell, Madhuri Namekar, and Vivek R. Nerurkar. 2014. "Infection with Non-Lethal West Nile Virus Eg101 Strain Induces Immunity that Protects Mice against the Lethal West Nile Virus NY99 Strain" Viruses 6, no. 6: 2328-2339. https://doi.org/10.3390/v6062328
APA StyleKumar, M., O'Connell, M., Namekar, M., & Nerurkar, V. R. (2014). Infection with Non-Lethal West Nile Virus Eg101 Strain Induces Immunity that Protects Mice against the Lethal West Nile Virus NY99 Strain. Viruses, 6(6), 2328-2339. https://doi.org/10.3390/v6062328