Antigenic Properties of N Protein of Hantavirus

Abstract
:1. Introduction
2. Hantavirus N Protein
2.1. Antigenic Profiling of N Protein Using Monoclonal Antibodies (MAbs) and Polyclonal Antibodies
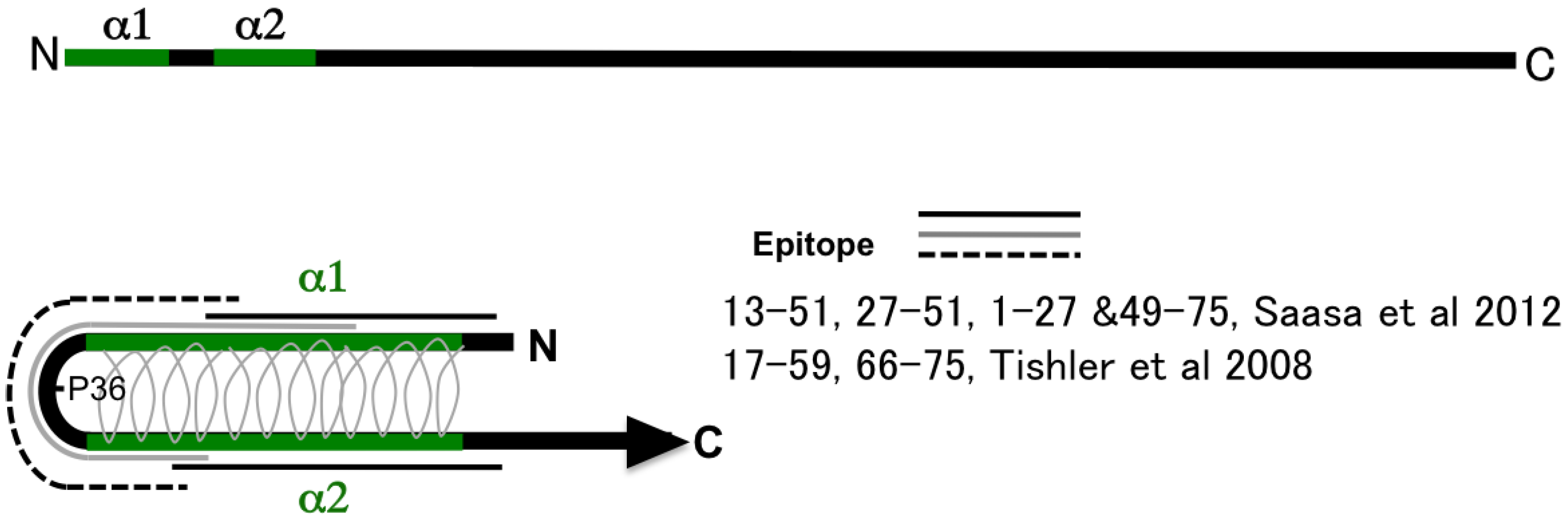
2.2. Deduced Structure of the N-Terminal Region of N Protein

2.3. Deduced Structure of the Central Part of N Protein

2.4. Deduced Structure of the C-Terminal Region of N Protein

2.5. Deduced Structure of Trimerized N Protein

2.6. Variety of Trimerization of N Protein Depending on the Viruses and Vectors

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region (aa) | HTNV | SEOV | DOBV | THAIV | PUUV | SNV | ANDV | LANV | BCCV | CARV |
|---|---|---|---|---|---|---|---|---|---|---|
| 1-429 (Entire) | M* | M* | M | M | M | S/M* | S | S | M | S |
| 50-429 | M* | M* | M | M | M | S | S* | S* | M | ND |
| 100-429 | M* | M* | M | ND | M | M | M | M | M | M |
| 155-429 | S | M | M | ND | M | S | ND | ND | ND | ND |
2.7. Association of Cellular Components with N
3. Conclusions
Acknowledgments
Conflicts of Interest
References and Notes
- Schmaljohn, C.S.; Hasty, S.E.; Dalrymple, J.M.; LeDuc, J.W.; Lee, H.W.; von Bonsdorff, C.H.; Brummer-Korvenkontio, M.; Vaheri, A.; Tsai, T.F.; Regnery, H.L.; et al. Antigenic and genetic properties of viruses linked to hemorrhagic fever with renal syndrome. Science 1985, 227, 1041–1044. [Google Scholar]
- Nichol, S.T.; Spiropoulou, C.F.; Morzunov, S.; Rollin, P.E.; Ksiazek, T.G.; Feldmann, H.; Sanchez, A.; Childs, J.; Zaki, S.; Peters, C.J. Genetic identification of a hantavirus associated with an outbreak of acute respiratory illness. Science 1993, 262, 914–917. [Google Scholar]
- Schmaljohn, C.S.; Hasty, S.E.; Harrison, S.A.; Dalrymple, J.M. Characterization hantaan virions, the prototype virus of hemorrhagic fever with renal syndrome. J. Infect. Dis. 1983, 148, 1005–1011. [Google Scholar] [PubMed]
- Jaaskelainen, K.M.; Kaukinen, P.; Minskaya, E.S.; Plyusnina, A.; Vapalahti, O.; Elliott, R.M.; Weber, F.; Vaheri, A.; Plyusnin, A. Tula and Puumala hantavirus NSs ORFs are functional and the products inhibit activation of the interferon-beta promoter. J. Med. Virol. 2007, 79, 1527–1536. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, J.O.; Jaaskelainen, K.M.; Djupsjobacka, J.; Vaheri, A.; Plyusnin, A. Tula hantavirus NSs protein accumulates in the perinuclear area in infected and transfected cells. Arch. Virol. 2010, 155, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Vera-Otarola, J.; Solis, L.; Soto-Rifo, R.; Ricci, E.P.; Pino, K.; Tischler, N.D.; Ohlmann, T.; Darlix, J.L.; Lopez-Lastra, M. The Andes hantavirus NSs protein is expressed from the viral small mRNA by a leaky scanning mechanism. J. Virol. 2012, 86, 2176–2187. [Google Scholar]
- Schmaljohn, C.S.; Sugiyama, K.; Schmaljohn, A.L.; Bishop, D.H. Baculovirus expression of the small genome segment of Hantaan virus and potential use of the expressed nucleocapsid protein as a diagnostic antigen. J. Gen. Virol. 1988, 69, 777–786. [Google Scholar] [PubMed]
- Vapalahti, O.; Lundkvist, A.; Kallio-Kokko, H.; Paukku, K.; Julkunen, I.; Lankinen, H.; Vaheri, A. Antigenic properties and diagnostic potential of puumala virus nucleocapsid protein expressed in insect cells. J. Clin. Microbiol. 1996, 34, 119–125. [Google Scholar] [PubMed]
- Ulrich, R.; Lundkvist, A.; Meisel, H.; Koletzki, D.; Sjolander, K.B.; Gelderblom, H.R.; Borisova, G.; Schnitzler, P.; Darai, G.; Kruger, D.H. Chimaeric HBV core particles carrying a defined segment of Puumala hantavirus nucleocapsid protein evoke protectiveimmunity In an animal model. Vaccine 1998, 16, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Brus Sjolander, K.; Golovljova, I.; Plyusnin, A.; Lundkvist, A. Diagnostic potential of puumala virus nucleocapsid protein expressed in Drosophila melanogaster cells. J. Clin. Microbiol. 2000, 38, 2324–2329. [Google Scholar]
- Gott, P.; Stohwasser, R.; Schnitzler, P.; Darai, G.; Bautz, E.K. RNA binding of recombinant nucleocapsid proteins of hantaviruses. Virology 1993, 194, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Severson, W.; Villegas, N.; Schmaljohn, C.S.; Jonsson, C.B. The RNA binding domain of the hantaan virus N protein maps to a central, conserved region. J. Virol. 2002, 76, 3301–3308. [Google Scholar] [CrossRef] [PubMed]
- Severson, W.; Xu, X.; Kuhn, M.; Senutovitch, N.; Thokala, M.; Ferron, F.; Longhi, S.; Canard, B.; Jonsson, C.B. Essential amino acids of the hantaan virus N protein in its interaction with RNA. J. Virol. 2005, 79, 10032–10039. [Google Scholar] [CrossRef] [PubMed]
- Alminaite, A.; Backstrom, V.; Vaheri, A.; Plyusnin, A. Oligomerization of hantaviral nucleocapsid protein: charged residues in the N-terminal coiled-coil domain contribute to intermolecular interactions. J. Gen. Virol. 2008, 89, 2167–2174. [Google Scholar] [CrossRef] [PubMed]
- Kaukinen, P.; Vaheri, A.; Plyusnin, A. Hantavirus nucleocapsid protein: A multifunctional molecule with both housekeeping and ambassadorial duties. Arch. Virol. 2005, 150, 1693–1713. [Google Scholar] [CrossRef] [PubMed]
- Vaheri, A.; Strandin, T.; Hepojoki, J.; Sironen, T.; Henttonen, H.; Makela, S.; Mustonen, J. Uncovering the mysteries of hantavirus infections. Nat. Rev. Microbiol. 2013, 11, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.L.; Friedman, R. Evolutionary diversification of protein-coding genes of hantaviruses. Mol. Biol. Evol. 2000, 17, 1558–1568. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.J.; Bennett, S.N.; Sumibcay, L.; Arai, S.; Hope, A.G.; Mocz, G.; Song, J.W.; Cook, J.A.; Yanagihara, R. Evolutionary insights from a genetically divergent hantavirus harbored by the European common mole (Talpa europaea). PLoS One 2009, 4, e6149. [Google Scholar] [CrossRef] [PubMed]
- Arai, S.; Nguyen, S.T.; Boldgiv, B.; Fukui, D.; Araki, K.; Dang, C.N.; Ohdachi, S.D.; Nguyen, N.X.; Pham, T.D.; Boldbaatar, B.; et al. Novel bat-borne hantavirus, Vietnam. Emerg. Infect. Dis. 2013, 19, 1159–1161. [Google Scholar] [CrossRef] [PubMed]
- Weiss, S.; Witkowski, P.T.; Auste, B.; Nowak, K.; Weber, N.; Fahr, J.; Mombouli, J.V.; Wolfe, N.D.; Drexler, J.F.; Drosten, C.; et al. Hantavirus in bat, Sierra Leone. Emerg. Infect. Dis. 2012, 18, 159–161. [Google Scholar] [CrossRef] [PubMed]
- Young, J.C.; Mills, J.N.; Enria, D.A.; Dolan, N.E.; Khan, A.S.; Ksiazek, T.G. New World hantaviruses. Br. Med. Bull. 1998, 54, 659–673. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, I.; Yoshimatsu, K.; Lee, B.H.; Okumura, M.; Taruishi, M.; Araki, K.; Kariwa, H.; Takashima, I.; Arikawa, J. Development of a serotyping ELISA system for Thailand virus infection. Arch. Virol. 2008, 153, 1537–1542. [Google Scholar] [CrossRef] [PubMed]
- Mills, J.N.; Amman, B.R.; Glass, G.E. Ecology of Hantaviruses and Their Hosts in North America. Vector Borne Zoonotic Dis. 2010, 10, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Palma, R.E.; Polop, J.J.; Owen, R.D.; Mills, J.N. Ecology of rodent-associated hantaviruses in the Southern Cone of South America: Argentina, Chile, Paraguay, and Uruguay. J. Wildl. Dis. 2012, 48, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Ruo, S.L.; Sanchez, A.; Elliott, L.H.; Brammer, L.S.; McCormick, J.B.; Fisher-Hoch, S.P. Monoclonal antibodies to three strains of hantaviruses: Hantaan, R22, and Puumala. Arch. Virol. 1991, 119, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yoshimatsu, K.; Arikawa, J.; Tamura, M.; Yoshida, R.; Lundkvist, A.; Niklasson, B.; Kariwa, H.; Azuma, I. Characterization of the nucleocapsid protein of hantaan virus strain 76–118 using monoclonal antibodies. J. Gen. Virol. 1996, 77, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Lundkvist, A.; Fatouros, A.; Niklasson, B. Antigenic variation of European haemorrhagic fever with renal syndrome virus strains characterized using bank vole monoclonal antibodies. J. Gen. Virol. 1991, 72, 2097–2103. [Google Scholar] [CrossRef] [PubMed]
- Lundkvist, A.; Bjorsten, S.; Niklasson, B.; Ahlborg, N. Mapping of B-cell determinants in the nucleocapsid protein of Puumala virus: definition of epitopes specific for acute immunoglobulin G recognition in humans. Clin. Diagn. Lab. Immunol. 1995, 2, 82–86. [Google Scholar] [PubMed]
- Elgh, F.; Lundkvist, A.; Alexeyev, O.A.; Wadell, G.; Juto, P. A major antigenic domain for the human humoral response to Puumala virus nucleocapsid protein is located at the amino-terminus. J. Virol. Methods 1996, 59, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Tischler, N.D.; Rosemblatt, M.; Valenzuela, P.D. Characterization of cross-reactive and serotype-specific epitopes on the nucleocapsid proteins of hantaviruses. Virus. Res. 2008, 135, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Saasa, N.; Yoshida, H.; Shimizu, K.; Sanchez-Hernandez, C.; Romero-Almaraz, M.D.; Koma, T.; Sanada, T.; Seto, T.; Yoshii, K.; Ramos, C.; et al. The N-terminus of the Montano virus nucleocapsid protein possesses broadly cross-reactive conformation-dependent epitopes conserved in rodent-borne hantaviruses. Virology 2012, 428, 48–57. [Google Scholar] [PubMed]
- Kucinskaite-Kodze, I.; Petraityte-Burneikiene, R.; Zvirbliene, A.; Hjelle, B.; Medina, R.A.; Gedvilaite, A.; Razanskiene, A.; Schmidt-Chanasit, J.; Mertens, M.; Padula, P.; et al. Characterization of monoclonal antibodies against hantavirus nucleocapsid protein and their use for immunohistochemistry on rodent and human samples. Arch. Virol. 2011, 156, 443–456. [Google Scholar]
- Song, J.W.; Baek, L.J.; Schmaljohn, C.S.; Yanagihara, R. Thottapalayam virus, a prototype shrewborne hantavirus. Emerg. Infect. Dis. 2007, 13, 980–985. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, M.; Tegshduuren, E.; Yoshimatsu, K.; Petraityte, R.; Sasnauskas, K.; Hammerschmidt, B.; Friedrich, R.; Mertens, M.; Groschup, M.H.; Arai, S.; et al. Novel serological tools for detection of Thottapalayam virus, a Soricomorpha-borne hantavirus. Arch. Virol. 2012, 157, 2179–2187. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Hjelle, B.; Lanzi, R.; Morris, C.; Anderson, B.; Jenison, S. Antibody responses to Four Corners hantavirus infections in the deer mouse (Peromyscus maniculatus): Identification of an immunodominant region of the viral nucleocapsid protein. J. Virol. 1995, 69, 1939–1943. [Google Scholar] [PubMed]
- Gott, P.; Zoller, L.; Darai, G.; Bautz, E.K.F. A Major Antigenic Domain Of Hantaviruses Is Located On the Aminoproximal Site Of the Viral Nucleocapsid Protein. Virus Genes 1997, 14, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Lundkvist, A.; Meisel, H.; Koletzki, D.; Lankinen, H.; Cifire, F.; Geldmacher, A.; Sibold, C.; Gott, P.; Vaheri, A.; Kruger, D.H.; et al. Mapping of B-cell epitopes in the nucleocapsid protein of Puumala hantavirus. Viral. Immunol. 2002, 15, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Elgh, F.; Lundkvist, A.; Alexeyev, O.A.; Stenlund, H.; Avsic-Zupanc, T.; Hjelle, B.; Lee, H.W.; Smith, K.J.; Vainionpaa, R.; Wiger, D.; et al. Serological diagnosis of hantavirus infections by an enzyme-linked immunosorbent assay based on detection of immunoglobulin G and M responses to recombinant nucleocapsid proteins of five viral serotypes. J. Clin. Microbiol. 1997, 35, 1122–1130. [Google Scholar] [PubMed]
- Elgh, F.; Linderholm, M.; Wadell, G.; Tarnvik, A.; Juto, P. Development of humoral cross-reactivity to the nucleocapsid protein of heterologous hantaviruses in nephropathia epidemica. FEMS Immunol. Med. Microbiol. 1998, 22, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Alfadhli, A.; Love, Z.; Arvidson, B.; Seeds, J.; Willey, J.; Barklis, E. Hantavirus nucleocapsid protein oligomerization. J. Virol. 2001, 75, 2019–2023. [Google Scholar] [CrossRef] [PubMed]
- Alfadhli, A.; Steel, E.; Finlay, L.; Bachinger, H.P.; Barklis, E. Hantavirus nucleocapsid protein coiled-coil domains. J. Biol. Chem. 2002, 277, 27103–27108. [Google Scholar] [CrossRef] [PubMed]
- Boudko, S.P.; Kuhn, R.J.; Rossmann, M.G. The coiled-coil domain structure of the Sin Nombre virus nucleocapsid protein. J. Mol. Biol. 2007, 366, 1538–1544. [Google Scholar] [CrossRef] [PubMed]
- Alminaite, A.; Halttunen, V.; Kumar, V.; Vaheri, A.; Holm, L.; Plyusnin, A. Oligomerization of hantavirus nucleocapsid protein: analysis of the N-terminal coiled-coil domain. J. Virol. 2006, 80, 9073–9081. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Boudreaux, D.M.; Estrada, D.F.; Egan, C.W.; St Jeor, S.C.; De Guzman, R.N. NMR structure of the N-terminal coiled coil domain of the Andes hantavirus nucleocapsid protein. J. Biol. Chem. 2008, 283, 28297–28304. [Google Scholar] [CrossRef] [PubMed]
- Amada, T.; Yoshimatsu, K.; Koma, T.; Shimizu, K.; Gamage, C.D.; Shiokawa, K.; Nishio, S.; Ahlm, C.; Arikawa, J. Development of an immunochromatography strip test based on truncated nucleocapsid antigens of three representative hantaviruses. Virol. J. 2014, in press. [Google Scholar]
- Okumura, M.; Yoshimatsu, K.; Araki, K.; Lee, B.H.; Asano, A.; Agui, T.; Arikawa, J. Epitope analysis of monoclonal antibody E5/G6, which binds to a linear epitope in the nucleocapsid protein of hantaviruses. Arch. Virol. 2004, 149, 2427–2434. [Google Scholar] [CrossRef] [PubMed]
- Mir, M.A.; Brown, B.; Hjelle, B.; Duran, W.A.; Panganiban, A.T. Hantavirus N protein exhibits genus-specific recognition of the viral RNA panhandle. J. Virol. 2006, 80, 11283–11292. [Google Scholar] [CrossRef] [PubMed]
- Mir, M.A.; Sheema, S.; Haseeb, A.; Haque, A. Hantavirus nucleocapsid protein has distinct m7G cap- and RNA-binding sites. J. Biol. Chem. 2010, 285, 11357–11368. [Google Scholar] [CrossRef] [PubMed]
- Yoshimatsu, K.; Lee, B.H.; Araki, K.; Morimatsu, M.; Ogino, M.; Ebihara, H.; Arikawa, J. The multimerization of hantavirus nucleocapsid protein depends on type-specific epitopes. J. Virol. 2003, 77, 943–952. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Rossi, C.; Schmaljohn, C.S. Expression of non-conserved regions of the S genome segments of three hantaviruses: evaluation of the expressed polypeptides for diagnosis of haemorrhagic fever with renal syndrome. J. Gen. Virol. 1993, 4, 1115–1124. [Google Scholar] [CrossRef]
- Koma, T.; Yoshimatsu, K.; Taruishi, M.; Miyashita, D.; Endo, R.; Shimizu, K.; Yasuda, S.P.; Amada, T.; Seto, T.; Murata, R.; et al. Development of a serotyping enzyme-linked immunosorbent assay system based on recombinant truncated hantavirus nucleocapsid proteins for New World hantavirus infection. J. Virol. Methods 2012, 185, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Kaukinen, P.; Kumar, V.; Tulimaki, K.; Engelhardt, P.; Vaheri, A.; Plyusnin, A. Oligomerization of Hantavirus N protein: C-terminal alpha-helices interact to form a shared hydrophobic space. J. Virol. 2004, 78, 13669–13677. [Google Scholar] [CrossRef] [PubMed]
- Mir, M.A.; Panganiban, A.T. Trimeric hantavirus nucleocapsid protein binds specifically to the viral RNA panhandle. J. Virol. 2004, 78, 8281–8288. [Google Scholar] [CrossRef] [PubMed]
- Kaukinen, P.; Vaheri, A.; Plyusnin, A. Mapping of the regions involved in homotypic interactions of Tula hantavirus N protein. J. Virol. 2003, 77, 10910–10916. [Google Scholar] [CrossRef] [PubMed]
- Morii, M.; Yoshimatsu, K.; Arikawa, J.; Zhou, G.; Kariwa, H.; Takashima, I. Antigenic characterization of Hantaan and Seoul virus nucleocapsid proteins expressed by recombinant baculovirus: application of a truncated protein, lacking an antigenic region common to the two viruses, as a serotyping antigen. J. Clin. Microbiol. 1998, 36, 2514–2521. [Google Scholar] [PubMed]
- Koma, T.; Yoshimatsu, K.; Pini, N.; Safronetz, D.; Taruishi, M.; Levis, S.; Endo, R.; Shimizu, K.; Yasuda, S.P.; Ebihara, H.; et al. Truncated hantavirus nucleocapsid proteins for serotyping Sin Nombre, Andes, and Laguna Negra hantavirus infections in humans and rodents. J. Clin. Microbiol. 2010, 48, 1635–1642. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.-H.; Yoshimatsu, K.; Maeda, A.; Ochiai, K.; Morimatsu, M.; Araki, K.; Ogino, M.; Morikawa, S.; Arikawa, J. Association of the nucleocapsid protein of the Seoul and Hantaan hantaviruses with small ubiquitin-like modifier-1-related molecules. Virus Research 2003, 98, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Kaukinen, P.; Vaheri, A.; Plyusnin, A. Non-covalent interaction between nucleocapsid protein of Tula hantavirus and small ubiquitin-related modifier-1, SUMO-1. Virus Res. 2003, 92, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, A.; Yasuda, J.; Takano, H.; Tanaka, Y.; Hatakeyama, M.; Shida, H. Regulation of human T-cell leukemia virus type 1 (HTLV-1) budding by ubiquitin ligase Nedd4. Microbes Infect. 2004, 6, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, J.; Hunter, E.; Nakao, M.; Shida, H. Functional involvement of a novel Nedd4-like ubiquitin ligase on retrovirus budding. EMBO Rep. 2002, 3, 636–640. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, J.; Nakao, M.; Kawaoka, Y.; Shida, H. Nedd4 regulates egress of Ebola virus-like particles from host cells. J. Virol. 2003, 77, 9987–9992. [Google Scholar] [CrossRef] [PubMed]
- Urata, S.; Noda, T.; Kawaoka, Y.; Yokosawa, H.; Yasuda, J. Cellular factors required for Lassa virus budding. J. Virol. 2006, 80, 4191–4195. [Google Scholar] [CrossRef] [PubMed]
- Cheng, E.; Haque, A.; Rimmer, M.A.; Hussein, I.T.; Sheema, S.; Little, A.; Mir, M.A. Characterization of the Interaction between hantavirus nucleocapsid protein (N) and ribosomal protein S19 (RPS19). J. Biol. Chem. 2011, 286, 11814–11824. [Google Scholar] [CrossRef] [PubMed]
- Haque, A.; Mir, M.A. Interaction of hantavirus nucleocapsid protein with ribosomal protein S19. J. Virol. 2010, 84, 12450–12453. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, H.N.; Jonsson, C.B. New and Old World hantaviruses differentially utilize host cytoskeletal components during their life cycles. Virology 2008, 374, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, H.N.; Chung, D.H.; Plane, S.J.; Sztul, E.; Chu, Y.K.; Guttieri, M.C.; McDowell, M.; Ali, G.; Jonsson, C.B. Dynein-dependent transport of the hantaan virus nucleocapsid protein to the endoplasmic reticulum-Golgi intermediate compartment. J. Virol. 2007, 81, 8634–8647. [Google Scholar] [CrossRef] [PubMed]
- Ravkov, E.V.; Nichol, S.T.; Peters, C.J.; Compans, R.W. Role Of Actin Microfilaments In Black Creek Canal Virus Morphogenesis. J. Virol. 1998, 72, 2865–2870. [Google Scholar] [PubMed]
- Hepojoki, J.; Strandin, T.; Wang, H.; Vapalahti, O.; Vaheri, A.; Lankinen, H. Cytoplasmic tails of hantavirus glycoproteins interact with the nucleocapsid protein. J. Gen. Virol. 2010, 91, 2341–2350. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Alminaite, A.; Vaheri, A.; Plyusnin, A. Interaction between hantaviral nucleocapsid protein and the cytoplasmic tail of surface glycoprotein Gn. Virus Res. 2010, 151, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Yoshimatsu, K.; Koma, T.; Arikawa, J. Role of Nucleocapsid Protein of Hantaviruses in Intracellular Traffic of Viral Glycoproteins. Virus Res. 2013, 178, 349–356. [Google Scholar] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yoshimatsu, K.; Arikawa, J. Antigenic Properties of N Protein of Hantavirus. Viruses 2014, 6, 3097-3109. https://doi.org/10.3390/v6083097
Yoshimatsu K, Arikawa J. Antigenic Properties of N Protein of Hantavirus. Viruses. 2014; 6(8):3097-3109. https://doi.org/10.3390/v6083097
Chicago/Turabian StyleYoshimatsu, Kumiko, and Jiro Arikawa. 2014. "Antigenic Properties of N Protein of Hantavirus" Viruses 6, no. 8: 3097-3109. https://doi.org/10.3390/v6083097
APA StyleYoshimatsu, K., & Arikawa, J. (2014). Antigenic Properties of N Protein of Hantavirus. Viruses, 6(8), 3097-3109. https://doi.org/10.3390/v6083097