
Bovine Lactoferrin Inhibits Toscana Virus Infection by Binding to Heparan Sulphate


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
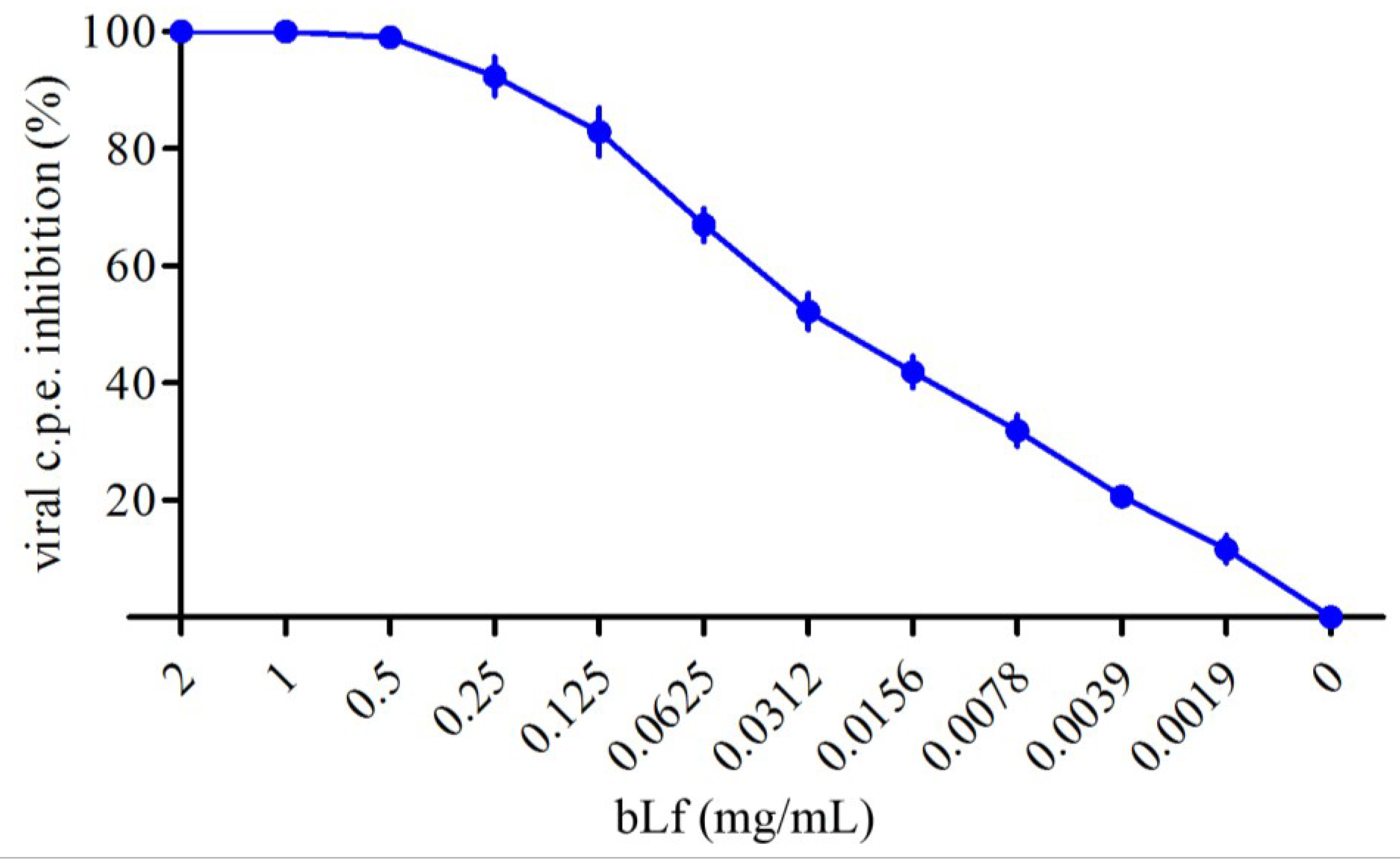
2.1. Activity of bLf on TosV Cytopathic Effect

2.2. Effect of bLf on Different Steps of Viral Infection

2.3. Neutralization of TosV Infection by Heparin

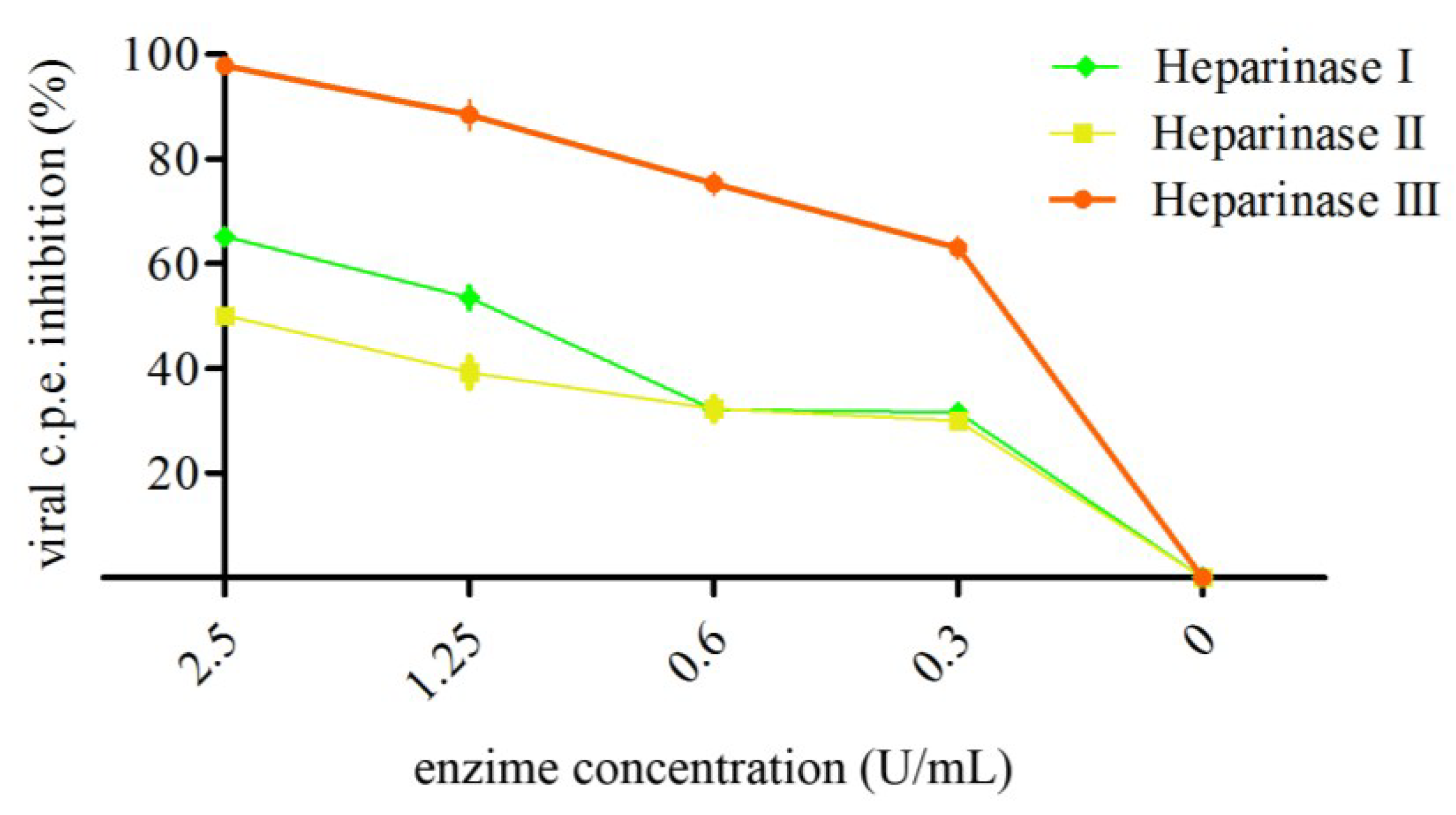
2.4. Modulation of Infectivity by Enzyme Treatment of Cell Surface GAGs


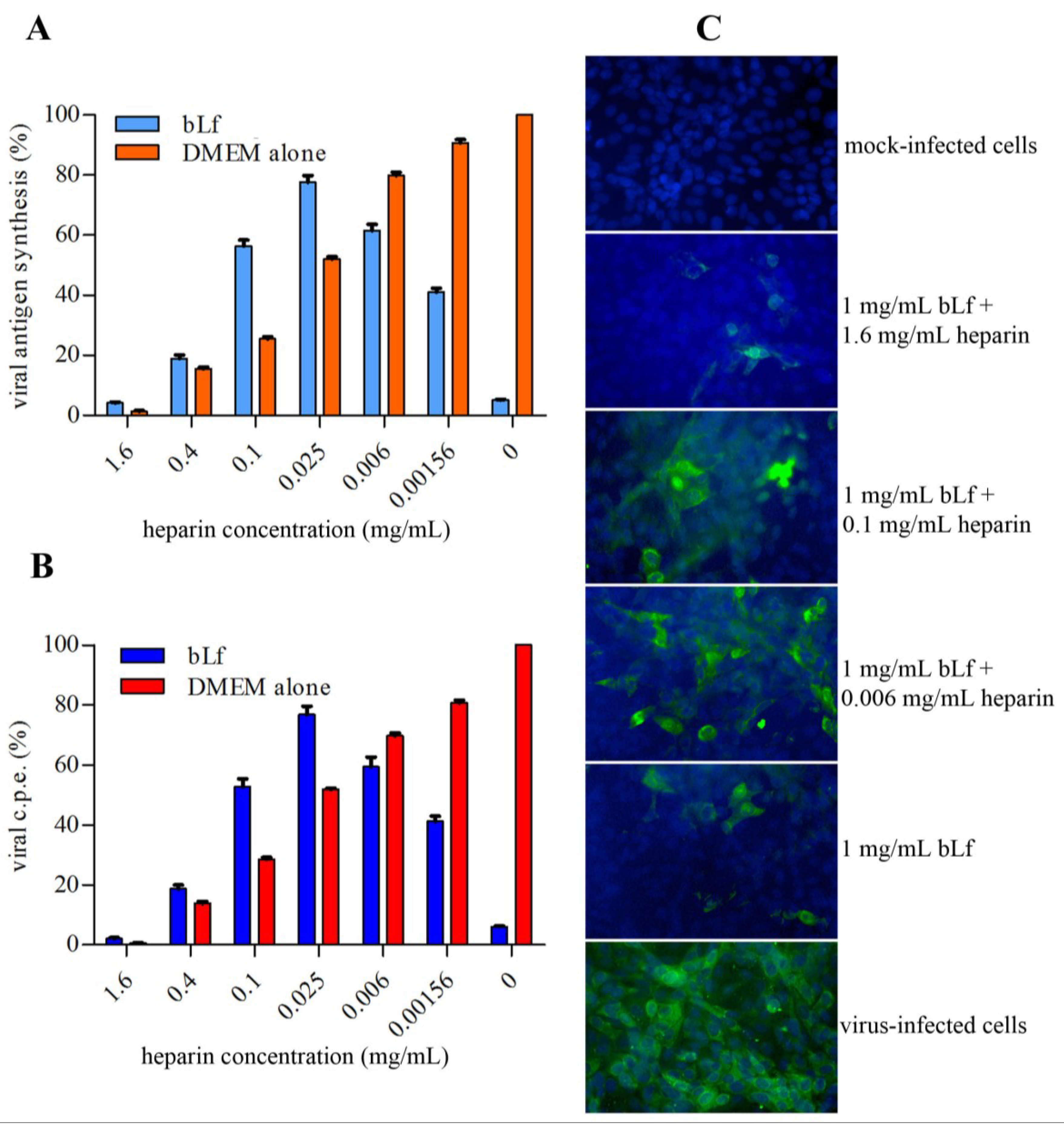
2.5. Effect of Lactoferrin and Heparin on TosV Infection
3. Discussion
4. Materials and Methods
4.1. Cells and Virus
4.2. Chemicals
4.3. Cytotoxicity Assay
4.4. Action of bLf on TosV Cytopathic Effect
4.5. Effect of bLf on Different Steps of TosV Infection
4.6. Inhibition of Infection by Heparin
4.7. Enzymatic Removal of GAGs from the Surfaces of Vero Cells
4.8. Immunofluorescence
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Accardi, L.; Grò, M.C.; di Bonito, P.; Giorgi, C. Toscana virus genomic L segment: Molecular cloning, coding strategy and amino acid sequence in comparison with other negative strand RNA viruses. Virus Res. 1993, 27, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Grò, M.C.; di Bonito, P.; Fortini, D.; Mochi, S.; Giorgi, C. Completion of molecular characterization of Toscana phlebovirus genome: Nucleotide sequence, coding strategy of M genomic segment and its amino acid sequence comparison to other phleboviruses. Virus Res. 1997, 51, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Verani, P.; Ciufolini, M.G.; Nicoletti, L.; Balducci, M.; Sabatinelli, G.; Coluzzi, M.; Paci, P.; Amaducci, L. Ecological and epidemiological studies of Toscana virus, an arbovirus isolated from Phlebotomus. Ann. Ist. Super. Sanità 1982, 18, 397–399. [Google Scholar] [PubMed]
- Verani, P.; Nicoletti, L.; Ciufolini, M.G. Antigenic and biological characterization of Toscana virus, a new Phlebotomus fever group virus isolated in Italy. Acta Virol. 1984, 28, 39–47. [Google Scholar] [PubMed]
- Charrel, R.N.; Gallian, P.; Navarro-Mari, J.M.; Nicoletti, L.; Papa, A.; Sánchez-Seco, M.P.; Tenorio, A.; de Lamballerie, X. Emergence of Toscana virus in Europe. Emerg. Infect. Dis. 2005, 11, 1657–1663. [Google Scholar] [CrossRef] [PubMed]
- Cusi, M.G.; Savellini, G.G.; Zanelli, G. Toscana virus epidemiology: From Italy to beyond. Open Virol. J. 2010, 4, 22–28. [Google Scholar] [PubMed]
- Francisci, D.; Papili, R.; Camanni, G.; Morosi, S.; Ferracchiato, N.; Valente, M.; Ciufolini, M.G.; Baldelli, F. Evidence of Toscana virus circulation in Umbria: first report. Eur. J. Epidemiol. 2003, 18, 457–459. [Google Scholar] [CrossRef] [PubMed]
- Di Nicuolo, G.; Pagliano, P.; Battisti, S.; Starace, M.; Mininni, V.; Attanasio, V.; Faella, F.S. Toscana virus central nervous system infections in southern Italy. J. Clin. Microbiol. 2005, 43, 6186–6188. [Google Scholar]
- Venturi, G.; Madeddu, G.; Rezza, G.; Ciccozzi, M.; Pettinato, M.L.; Cilliano, M.; Fiorentini, C.; Mura, M.S.; Ciufolini, M.G. Detection of Toscana virus central nervous system infections in Sardinia Island, Italy. J. Clin. Virol. 2007, 40, 90–91. [Google Scholar] [CrossRef] [PubMed]
- Colomba, C.; Saporito, L.; Ciufolini, M.G.; Marchi, A.; Rotolo, V.; de Grazia, S.; Titone, L.; Giammanco, G.M. Prevalence of Toscana sandfly fever virus antibodies in neurological patients and control subjects in Sicily. New Microbiol. 2012, 35, 161–165. [Google Scholar] [PubMed]
- Alkan, C.; Bichaud, L.; de Lamballerie, X.; Alten, B.; Gould, E.A.; Charrel, R.N. Sandfly-borne phleboviruses of Eurasia and Africa: epidemiology, genetic diversity, geographic range, control measures. Antivir. Res. 2013, 100, 54–74. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, L.; Verani, P.; Caciolli, S.; Ciufolini, M.G.; Renzi, A.; Bartolozzi, D.; Paci, P.; Leoncini, F.; Padovani, P.; Traini, E.; et al. Central nervous system involvement during infection by Phlebovirus toscana of residents in natural foci in central Italy (1977–1988). Am. J. Trop. Med. Hyg. 1991, 45, 429–434. [Google Scholar] [PubMed]
- Dupouey, J.; Bichaud, L.; Ninove, L.; Zandotti, C.; Thirion-Perrier, L.; de Lamballerie, X.; Charrel, R.N. Toscana virus infections: A case series from France. J. Infect. 2014, 68, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Tesh, R.B. The genus Phlebovirus and its vectors. Annu. Rev. Entomol. 1988, 33, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Braito, A.; Corbisiero, R.; Corradini, S.; Marchi, B.; Sancasciani, N.; Fiorentini, C.; Ciufolini, M.G. Evidence of Toscana virus infections without central nervous system involvement: A serological study. Eur. J. Epidemiol. 1997, 13, 761–764. [Google Scholar]
- Braito, A.; Ciufolini, M.G.; Pippi, L.; Corbisiero, R.; Fiorentini, C.; Gistri, A.; Toscano, L. Phlebotomus-transmitted Toscana virus infections of the central nervous system: A seven-year experience in Tuscany. Scand. J. Infect. Dis. 1998, 30, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Braito, A.; Corbisiero, R.; Corradini, S.; Fiorentini, C.; Ciufolini, M.G. Toscana virus infections of the central nervous system in children: A report of 14 cases. J. Pediatr. 1998, 132, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Valassina, M.; Meacci, F.; Valensin, P.E.; Cusi, M.G. Detection of neurotropic viruses circulating in Tuscany: The incisive role of Toscana virus. J. Med. Virol. 2000, 60, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Valassina, M.; Cusi, M.G.; Valensin, P.E. A Mediterranean arbovirus: the Toscana virus. J. Neurovirol. 2003, 9, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Valassina, M.; Valentini, M.; Pugliese, A.; Valensin, P.E.; Cusi, M.G. Serological survey of Toscana virus infections in a high-risk population in Italy. Clin. Diagn. Lab. Immunol. 2003, 10, 483–484. [Google Scholar]
- Dionisio, D.; Valassina, M.; Ciufolini, M.G.; Vivarelli, A.; Esperti, F.; Cusi, M.G.; Marchi, A.; Mazzoli, F.; Lupi, C. Encephalitis without meningitis due to sandfly fever virus serotype toscana. Clin. Infect. Dis. 2001, 32, 1241–1243. [Google Scholar] [CrossRef] [PubMed]
- Baldelli, F.; Ciufolini, M.G.; Francisci, D.; Marchi, A.; Venturi, G.; Fiorentini, C.; Luchetta, M.L.; Bruto, L.; Pauluzzi, S. Unusual presentation of life-threatening Toscana virus meningoencephalitis. Clin. Infect. Dis. 2004, 38, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Zanelli, G.; Bianco, C.; Cusi, M.G. Testicular involvement during Toscana virus infection: An unusual manifestation? Infection 2013, 41, 735–736. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Garcia, F.A.; Moreno-Docon, A.; Lopez-Lopez, M.; Albert-Lacal, L.; Martinez-Toldos, M.C.; Segovia-Hernandez, M.; Fernandez-Barreiro, A. A case of meningitis due to Toscana virus in Murcia. Rev. Neurol. 2007, 45, 317–318. [Google Scholar] [PubMed]
- Pauli, C.; Schwarz, T.F.; Meyer, C.G.; Jager, G. Neurological symptoms after an infection by the sandfly fever virus. Dtsch. Med. Wochenschr. 1995, 120, 1468–1472. [Google Scholar] [CrossRef] [PubMed]
- Serata, D.; Rapinesi, C.; del Casale, A.; Simonetti, A.; Mazzarini, L.; Ambrosi, E.; Kotzalidis, G.D.; Fensore, C.; Girardi, P.; Tatarelli, R. Personality changes after Toscana virus (Toscana virus) encephalitis in a 49-year-old man: A case report. Int. J. Neurosci. 2011, 121, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, J.; Bewermeyer, H.; Hartmann-Klosterkoetter, U.; Emmerich, P.; Schilling, S.; Valassina, M. Toscana virus causing severe meningoencephalitis in an elderly traveller. J. Neurol. Neurosurg. Psychiatry 2005, 76, 1605–1606. [Google Scholar] [CrossRef] [PubMed]
- Bartels, S.; de Boni, L.; Kretzschmar, H.A.; Heckmann, J.G. Lethal encephalitis caused by the Toscana virus in an elderly patient. J. Neurol. 2012, 259, 175–177. [Google Scholar] [CrossRef] [PubMed]
- Legrand, D.; Mazurier, J.; Colavizza, D.; Montreuil, J.; Spik, G. Properties of the iron-binding site of the N-terminal lobe of human and bovine lactotransferrins. Importance of the glycan moiety and of the non-covalent interactions between the N- and C-terminal lobes in the stability of the iron-binding site. Biochem. J. 1990, 266, 575–581. [Google Scholar] [PubMed]
- Gennaro, R.; Dewald, B.; Horisberger, U.; Gubler, H.U.; Baggiolini, M. A novel type of cytoplasmic granule in bovine neutrophils. J. Cell Biol. 1983, 96, 1651–1661. [Google Scholar] [CrossRef] [PubMed]
- Levay, P.F.; Viljoen, M. Lactoferrin: A general review. Haematologica 1995, 80, 252–267. [Google Scholar] [PubMed]
- Vorland, L.H. Lactoferrin: A multifunctional glycoprotein. APMIS 1999, 107, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Van der Strate, B.W.A.; Beljaars, L.; Molema, G.; Harmsen, M.C.; Meijer, D.K.F. Antiviral activities of lactoferrin. Antivir. Res. 2001, 52, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, M.; Superti, F. Recent developments in antiviral research. In Antiviral Activity of Lactoferrin; Pandalai, S.G., Ed.; Transworld Research Network: Trivandrum, India, 2001; pp. 193–203. [Google Scholar]
- Harmsen, M.C.; Swart, P.J.; de B´ethune, M.P.; Pawels, R.; de Clercq, E.; Th´e, T.H.; Meijer, D.K.F. Antiviral effects of plasma and milk proteins: lactoferrin shows a potent activity against both human immunodeficiencyvirus and human cytomegalovirus replication in vitro. J. Infect. Dis. 1995, 172, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Beljaars, L.; van der Strate, B.W.; Bakker, H.I.; Reker-Smit, C.; van Loenen-Weemaes, A.M.; Wiegmans, F.C.; Harmsen, M.C.; Molema, G.; Meijer, D.K. Inhibition of cytomegalovirus infection by lactoferrin in vitro and in vivo. Antivir. Res. 2004, 63, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, K.; Motsuchi, W.; Tanaka, S.; Dosako, S. Inhibition with lactoferrin of in vitro infection with human herpes virus. Jpn. J. Med. Sci. Biol. 1994, 47, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, M.; Trybala, E.; Superti, F.; Johansson, M.; Bergström, T. Inhibition of herpes simplex virus infection by lactoferrin is dependent on interference with the virus binding to glycosaminoglycans. Virology 2004, 18, 405–413. [Google Scholar] [CrossRef]
- Marchetti, M.; Ammendolia, M.G.; Superti, F. Glycosaminoglycans are not indispensable for the anti-herpes simplex virus type 2 activity of lactoferrin. Biochimie 2009, 91, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Ammendolia, M.G.; Marchetti, M.; Superti, F. Bovine lactoferrin prevents the entry and intercellular spread of herpes simplex virus type 1 in Green Monkey Kidney cells. Antivir. Res. 2007, 76, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Yi, M.; Kaneko, S.; Yu, D.Y.; Murakami, S. Hepatitis C virus envelope proteins bind lactoferrin. J. Virol. 1997, 71, 5997–6002. [Google Scholar] [PubMed]
- Murphy, M.E.; Kariwa, H.; Mizutani, T.; Yoshimatsu, K.; Arikawa, J.; Takashima, I. In vitro antiviral activity of lactoferrin and ribavirin upon hantavirus. Arch. Virol. 2000, 145, 1571–1582. [Google Scholar] [CrossRef] [PubMed]
- Hara, K.; Ikeda, M.; Saito, S.; Matsumoto, S.; Numata, K.; Kato, N.; Tanaka, K.; Sekihara, H. Lactoferrin inhibits hepatitis B virus infection in cultured human hepatocytes. Hepatol. Res. 2002, 24, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Sano, H.; Nagai, K.; Tsutsumi, H.; Kuroki, Y. Lactoferrin and surfactant protein A exhibit distinct binding specificity to F protein and differently modulate respiratory syncytial virus infection. Eur. J. Immunol. 2003, 33, 2894–2902. [Google Scholar] [CrossRef] [PubMed]
- Pietrantoni, A.; Dofrelli, E.; Tinari, A.; Ammendolia, M.G.; Puzelli, S.; Fabiani, C.; Donatelli, I.; Superti, F. Bovine lactoferrin inhibits influenza A virus induced programmed cell death in vitro. Biometals 2010, 23, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Pietrantoni, A.; Ammendolia, M.G.; Superti, F. Bovine lactoferrin: involvement of metal saturation and carbohydrates in the inhibition of influenza virus infection. Biochem. Cell Biol. 2012, 90, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Chien, Y.J.; Chen, W.J.; Hsu, W.L.; Chiou, S.S. Bovine lactoferrin inhibits Japanese encephalitis virus by binding to heparan sulfate and receptor for low density lipoprotein. Virology 2008, 379, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.A.; Sousa, I.P., Jr.; Silva, J.L.; Oliveira, A.C.; Gonçalves, R.B.; Gomes, A.M. Inhibition of Mayaro virus infection by bovine lactoferrin. Virology 2014, 452–453, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.; Magura, C.E.; Hurley, W.L. Heparin-binding properties of lactoferrin and lysozyme. Comp. Biochem. Physiol. 1992, 103, 889–895. [Google Scholar]
- De Boer, S.M.; Kortekaas, J.; de Haan, C.A.; Rottier, P.J.; Moormann, R.J.; Bosch, B.J. Heparan sulfate facilitates Rift Valley fever virus entry into the cell. J. Virol. 2012, 86, 13767–13771. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Maguire, T.; Hileman, R.E.; Fromm, J.R.; Esko, J.D.; Linhardt, R.J.; Marks, R.M. Dengue virus infectivity depends on envelope protein binding to target cell heparan sulfate. Nat. Med. 1997, 3, 866–871. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, H.; Oda, H.; Yamauchi, K.; Abe, F. Lactoferrin for prevention of common viral infections. J. Infect. Chemother. 2014. [Google Scholar] [CrossRef]
- Waarts, B.L.; Aneke, O.J.; Smit, J.M.; Kimata, K.; Bittman, R.; Meijer, D.K.; Wilschut, J. Antiviral activity of human lactoferrin: Inhibition of alphavirus interaction with heparan sulfate. Virology 2005, 333, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Shimazaki, K.; Tazume, T.; Uji, K.; Tanaka, M.; Kumura, H.; Mikawa, K.; Shimo-Oka, T. Properties of a Heparin-binding peptide derived from bovine lactoferrin. J. Dairy Sci. 1998, 81, 2841–2849. [Google Scholar] [CrossRef] [PubMed]
- Shimazaki, K.; Uji, K.; Tazume, T.; Kumura, H.; Shimo-Oka, T. Approach to identification and comparison of the heparin-interacting sites of lactoferrin using synthetic peptides. In Lactoferrin: Structure, Function and Applications; Shimazaki, K., Tsuda, H., Tomita, M., Kuwata, T., Perraudin, J.P., Eds.; Elsevier Science B.V: Amsterdam, The Netherland, 2000; pp. 37–46. [Google Scholar]
- Haywood, A.M. Virus receptors: Binding, adhesion, strengthening, and changes in viral structure. J. Virol. 1994, 68, 1–5. [Google Scholar] [PubMed]
- Rostand, K.S.; Esko, J.D. Microbial adherence to and invasion through proteoglycans. Infect. Immun. 1997, 6, 1–8. [Google Scholar]
- Byrnes, A.P.; Griffin, D.E. Binding of Sindbis virus to cell surface heparan sulfate. J. Virol. 1998, 72, 7349–7356. [Google Scholar] [PubMed]
- Bernard, K.A.; Klimstra, W.B.; Johnston, R.E. Mutations in the E2 glycoprotein of Venezuelan equine encephalitis virus confer heparan sulfate interaction, low morbidity, and rapid clearance from blood of mice. Virology 2000, 276, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Summerford, C.; Samulski, R.J. Membrane-associated heparan sulfate proteoglycan is a receptor for adeno-associated virus type 2 virions. J. Virol. 1998, 72, 1438–1445. [Google Scholar] [PubMed]
- Jackson, T.; Ellard, F.M.; Ghazaleh, R.A.; Brookes, S.M.; Blakemore, W.E.; Corteyn, A.H.; Stuart, D.I.; Newman, J.W.; King, A.M. Efficient infection of cells in culture by type O foot-and-mouth disease virus requires binding to cell surface heparan sulfate. J. Virol. 1996, 70, 5282–5287. [Google Scholar]
- Compton, T.; Nowlin, D.M.; Cooper, N.R. Initiation of human cytomegalovirus-infection requires initial interaction with cell surface heparan sulfate. Virology 1993, 193, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Birkmann, A.; Mahr, K.; Ensser, A.; Yaguboglu, S.; Titgemeyer, F.; Fleckenstein, B.; Neipel, F. Cell surface heparan sulfate is a receptor for human herpesvirus 8 and interacts with envelope glycoprotein K8.1. J. Virol. 2001, 75, 11583–11593. [Google Scholar] [CrossRef] [PubMed]
- Shukla, D.; Spear, P.G. Herpesviruses and heparan sulfate: An intimate relationship in aid of viral entry. J. Clin. Invest. 2001, 108, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Trybala, E.; Roth, A.; Johansson, M.; Liljeqvist, J.A.; Rekabdar, E.; Larm, O.; Bergstrom, T. Glycosaminoglycan-binding ability is a feature of wild-type strains of herpes simplex virus type 1. Virology 2002, 302, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.; Yanagishita, M.; Roderiquez, G.; Bouhabib, D.C.; Oravecz, T.; Hascall, V.C.; Norcross, M.A. Cell surface heparan sulfate proteoglycan mediates HIV-1 infection of T-cell lines. Aids Res. Hum. Retrovir. 1993, 9, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Goodfellow, I.G.; Sioofy, A.B.; Powell, R.M.; Evans, D.J. Echoviruses bind heparan sulfate at the cell surface. J. Virol. 2001, 75, 4918–4921. [Google Scholar] [CrossRef] [PubMed]
- Germi, R.; Crance, J.M.; Garin, D.; Guimet, J.; Lortat-Jacob, H.; Ruigrok, R.W.; Zarski, J.P.; Drouet, E. Heparan sulfate-mediated binding of infectious dengue virus type 2 and yellow fever virus. Virology 2002, 292, 162–168. [Google Scholar] [CrossRef]
- Dechecchi, M.C.; Melotti, P.; Bonizzato, A.; Santacatterina, M.; Chilosi, M.; Cabrini, G. Heparan sulfate glycosaminoglycans are receptors sufficient to mediate the initial binding of adenovirus types 2 and 5. J. Virol. 2001, 75, 8772–8780. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Thorp, S.C. Cell surface heparan sulfate and its roles in assisting viral infections. Med. Res. Rev. 2002, 22, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Depaquit, J.; Grandadam, M.; Fouque, F.; Andry, P.E.; Peyrefitte, C. Arthropod-borne viruses transmitted by Phlebotomine sandflies in Europe: A review. Euro Surveill. 2010, 15, e19507. [Google Scholar]
- Peyrefitte, C.N.; Grandadam, M.; Bessaud, M.; Andry, P.E.; Fouque, F.; Caro, V.; Diancourt, L.; Schuffenecker, I.; Pagès, F.; Tolou, H.; et al. Diversity of Phlebotomus perniciosus in Provence, southeastern France: Detection of two putative new phlebovirus sequences. Vector Borne Zoonotic Dis. 2013, 13, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Remoli, M.E.; Fortuna, C.; Marchi, A.; Bucci, P.; Argentini, C.; Bongiorno, G.; Maroli, M.; Gradoni, L.; Gramiccia, M.; Ciufolini, M.G. Viral isolates of a novel putative phlebovirus in the Marche Region of Italy. Am. J. Trop. Med. Hyg. 2014, 90, 760–763. [Google Scholar] [CrossRef] [PubMed]
- Venturi, G.; Ciccozzi, M.; Montieri, S.; Bartoloni, A.; Francisci, D.; Nicoletti, L.; Fortuna, C.; Marongiu, L.; Rezza, G.; Ciufolini, M.G. Genetic variability of the M genome segment of clinical and environmental Toscana virus strains. J. Gen. Virol. 2007, 88, 1288–1294. [Google Scholar] [CrossRef] [PubMed]
- Pietrantoni, A.; Ammendolia, M.G.; Tinari, A.; Siciliano, R.; Valenti, P.; Superti, F. Bovine lactoferrin peptidic fragments involved in inhibition of Echovirus 6 in vitro infection. Antivir. Res. 2006, 69, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Groves, M.L. The isolation of a red protein from milk. J. Am. Chem. Soc. 1960, 82, 3345–3350. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pietrantoni, A.; Fortuna, C.; Remoli, M.E.; Ciufolini, M.G.; Superti, F. Bovine Lactoferrin Inhibits Toscana Virus Infection by Binding to Heparan Sulphate. Viruses 2015, 7, 480-495. https://doi.org/10.3390/v7020480
Pietrantoni A, Fortuna C, Remoli ME, Ciufolini MG, Superti F. Bovine Lactoferrin Inhibits Toscana Virus Infection by Binding to Heparan Sulphate. Viruses. 2015; 7(2):480-495. https://doi.org/10.3390/v7020480
Chicago/Turabian StylePietrantoni, Agostina, Claudia Fortuna, Maria Elena Remoli, Maria Grazia Ciufolini, and Fabiana Superti. 2015. "Bovine Lactoferrin Inhibits Toscana Virus Infection by Binding to Heparan Sulphate" Viruses 7, no. 2: 480-495. https://doi.org/10.3390/v7020480