
Genetic Diversity of Koala Retroviral Envelopes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmids
2.2. Cells
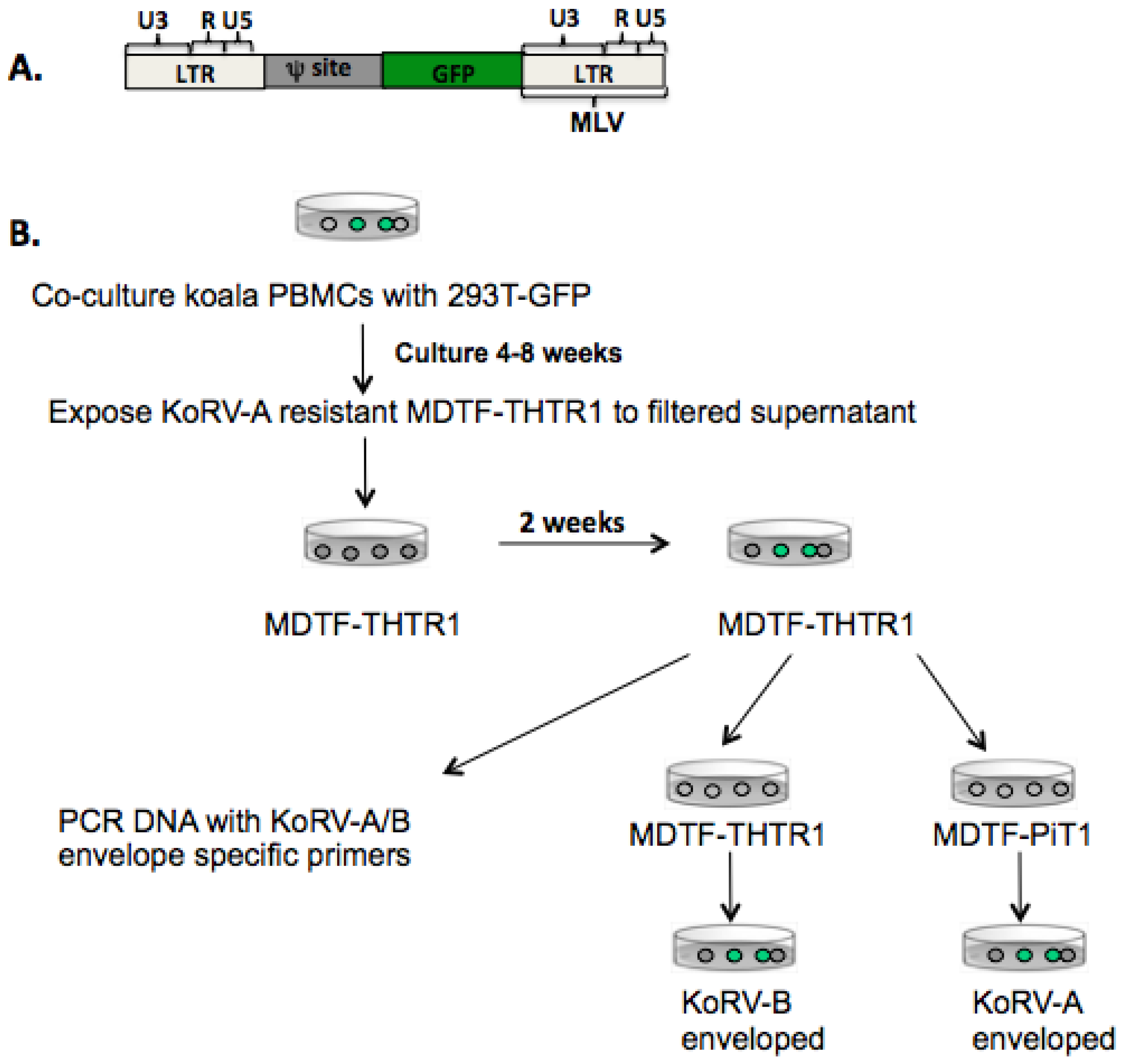
2.3. Transfection and Transduction
2.4. Preparation of Koala Samples
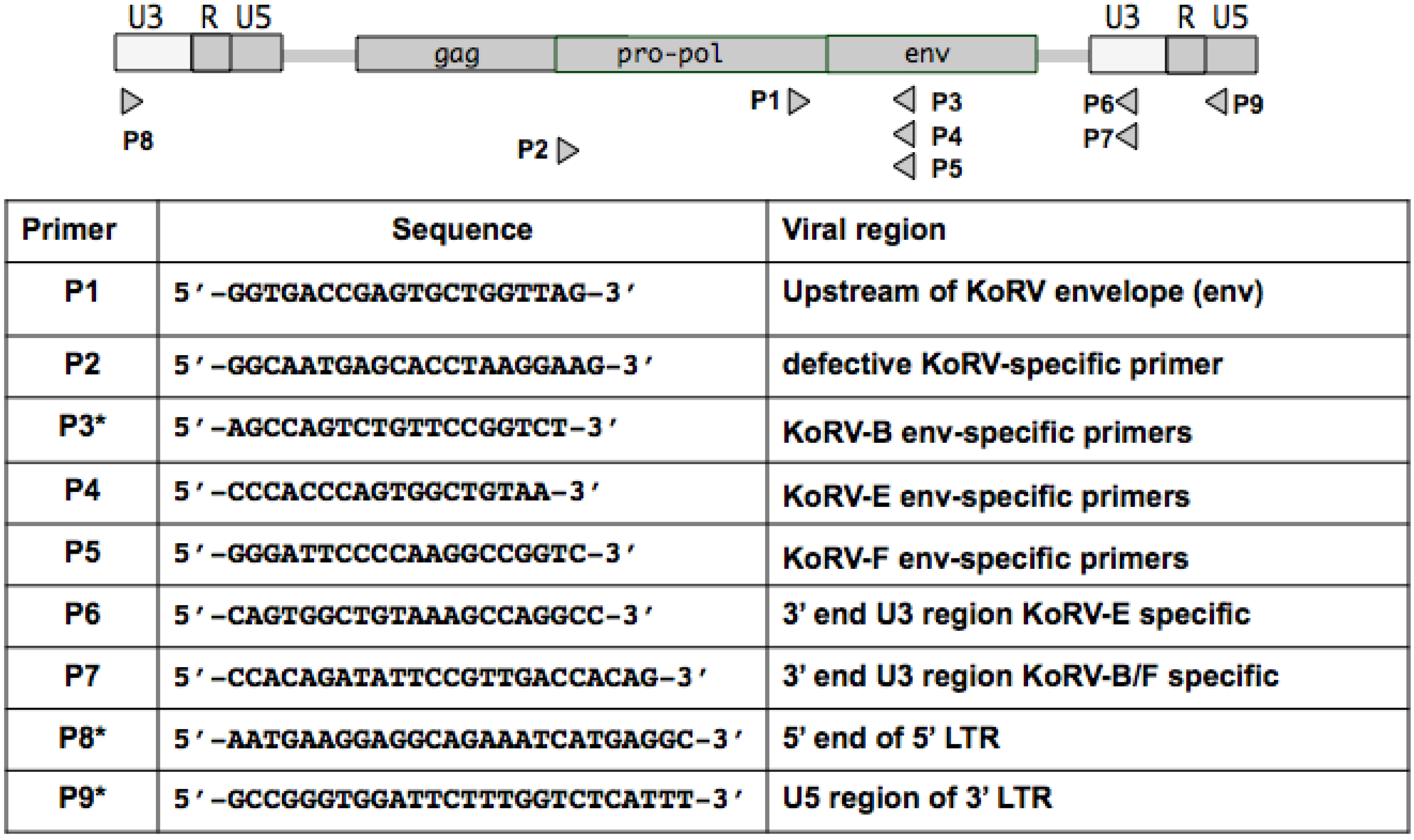
2.5. PCR and RT-PCR

3. Results
3.1. KoRV-B Envelope Can Pseudotype Infectious KoRV-A Viral Particles
3.2. Detection of Two Additional KoRV Envelope Variants by PCR Amplication


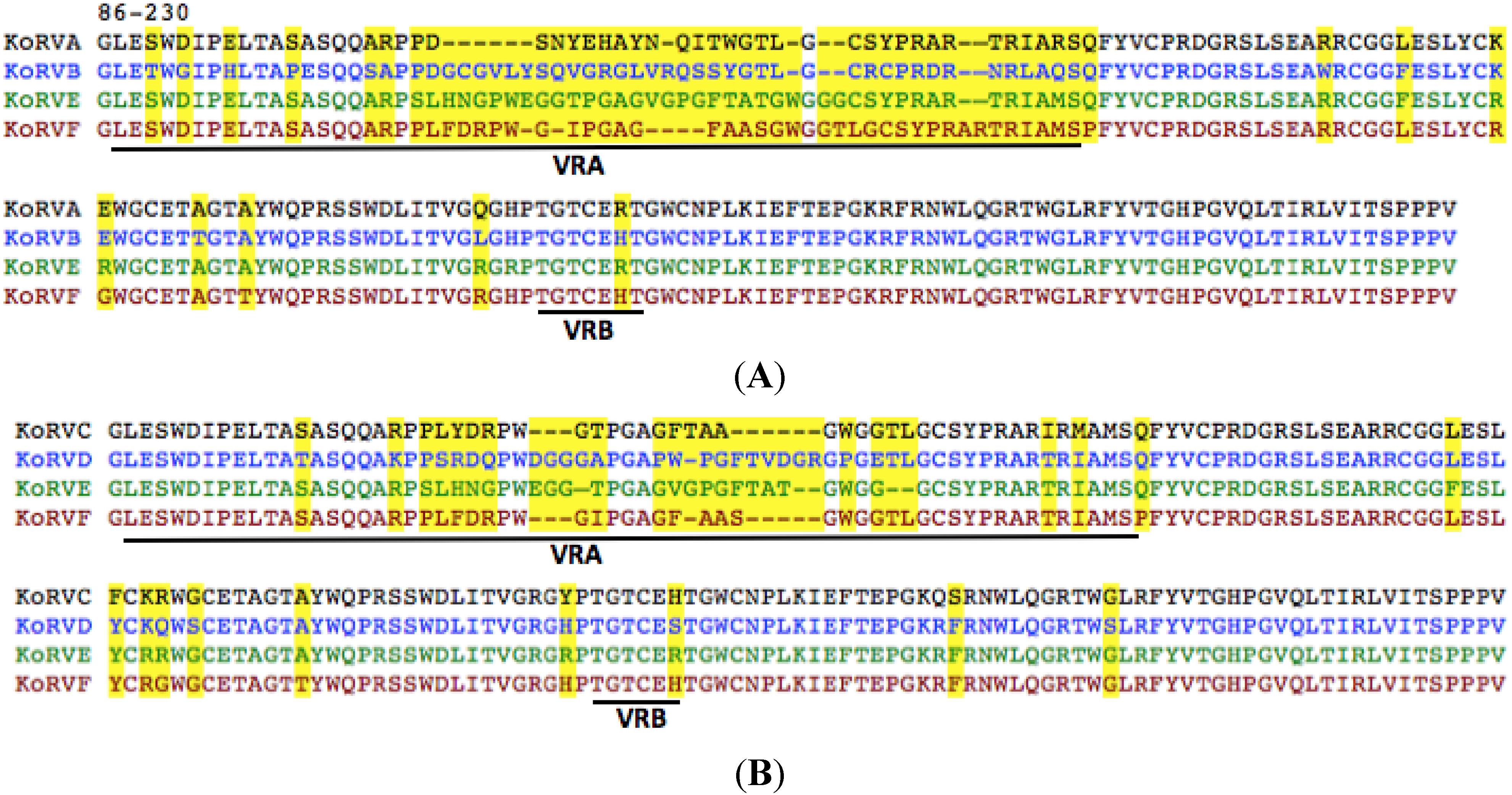
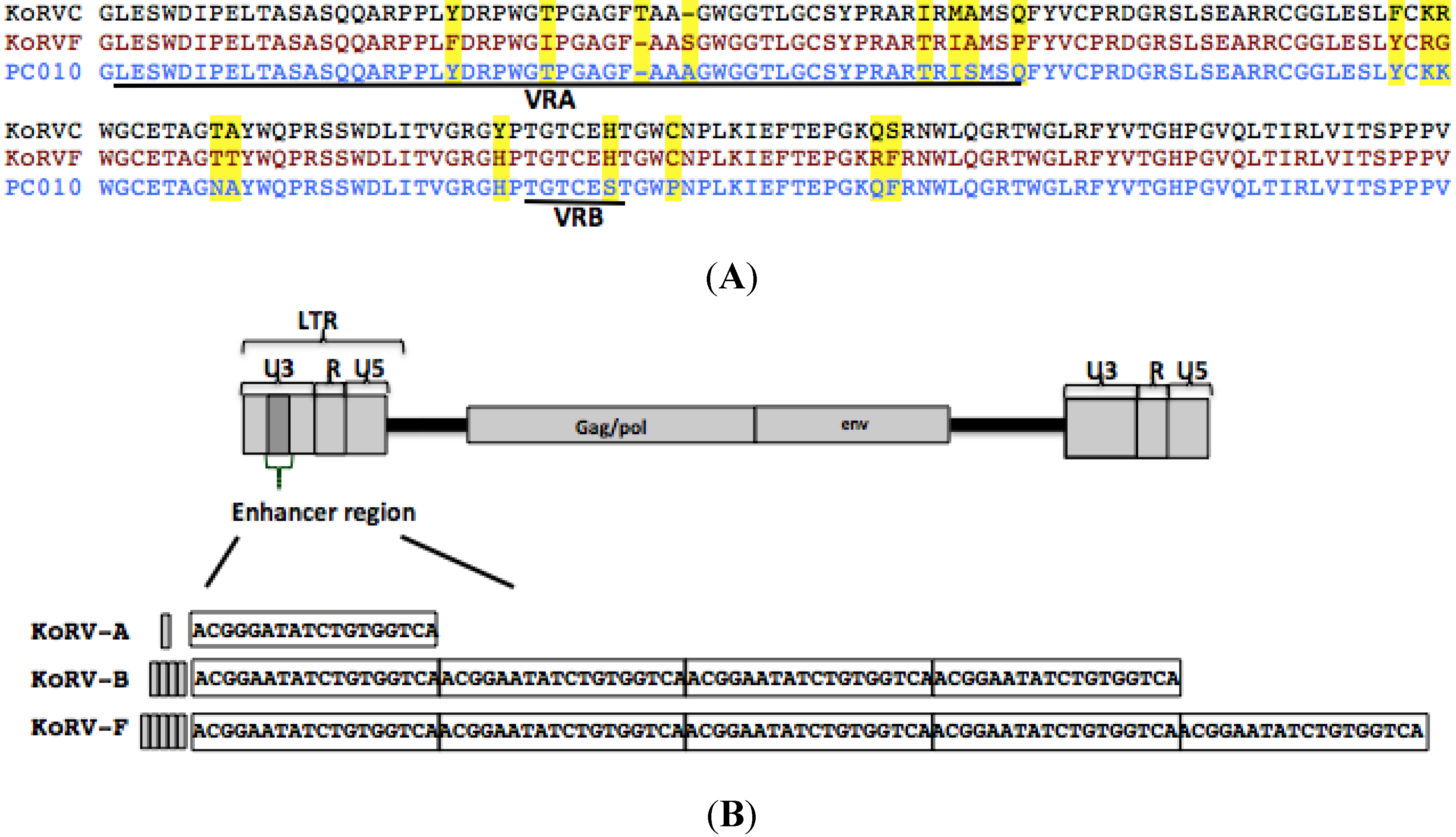
3.3. The KoRV-F Env ORF Is Closely Related to Previously Characterized KoRV Variants

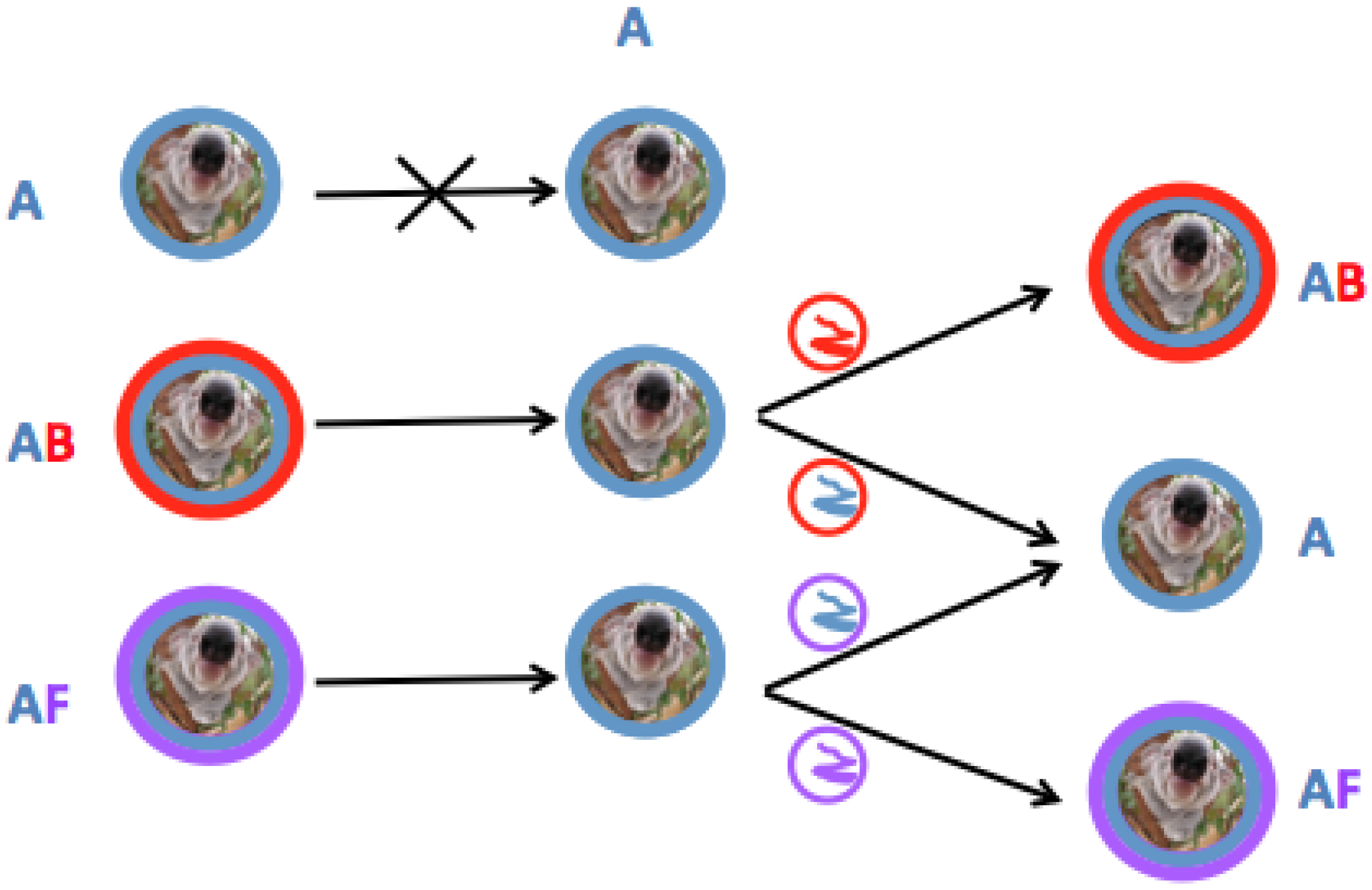
3.4. KoRV-E and KoRV-F Are Not Transmitted as Endogenous Retroviral Elements

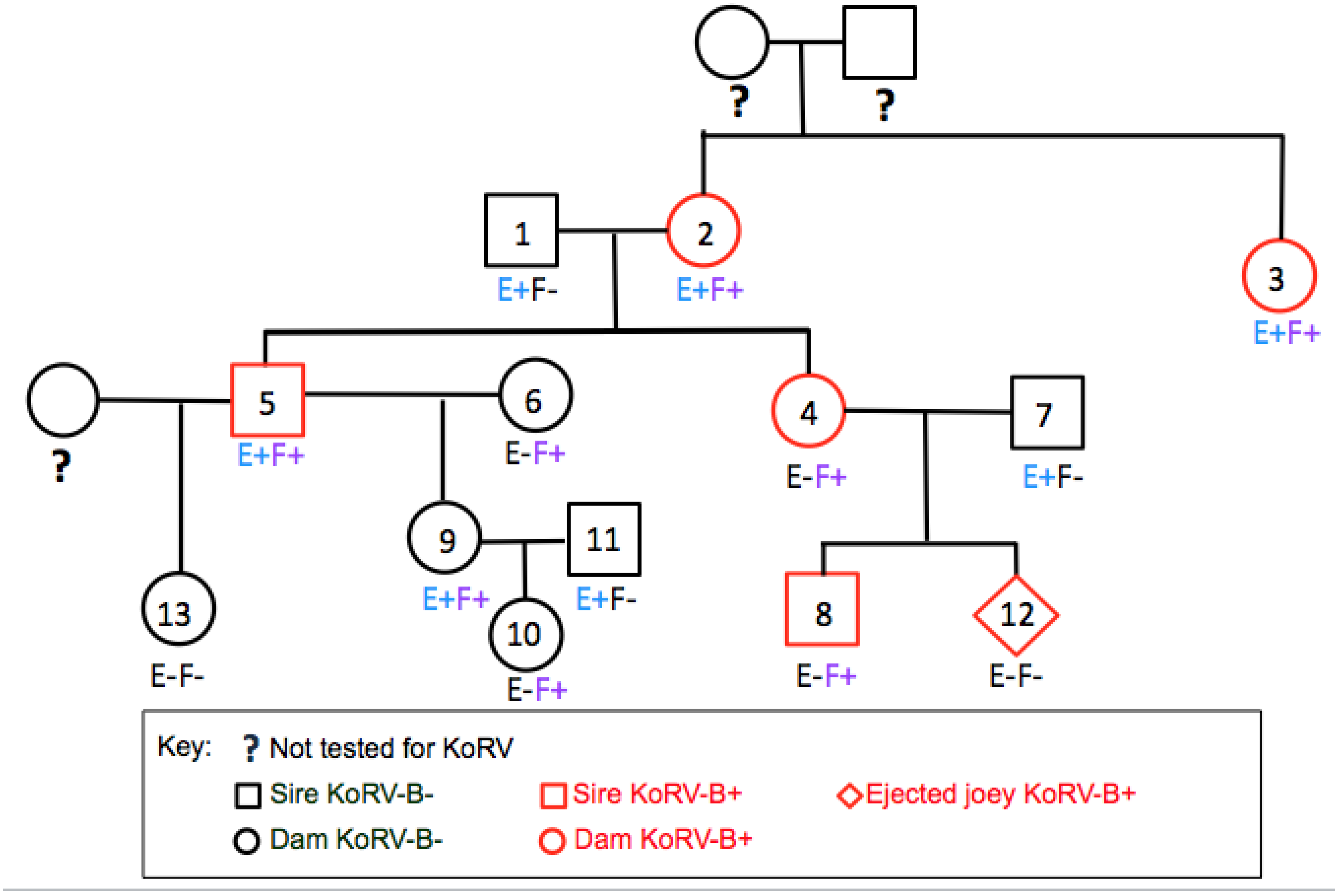
3.5. Defective KoRV-B, -E and -F Were Detected in Koalas Containing Intact KoRV-B Virus or KoRV-E and -F Envelopes
4. Discussion

5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Canfield, P.J.; Brown, A.S.; Kelly, W.R.; Sutton, R.H. Spontaneous lymphoid neoplasia in the koala (Phascolarctos cinereus). J. Comp. Pathol. 1987, 97, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Canfield, P.J.; Sabine, J.M.; Love, D.N. Virus particles associated with leukaemia in a koala. Aust. Vet. J. 1988, 65, 327–328. [Google Scholar] [CrossRef] [PubMed]
- Hanger, J.J.; Bromham, L.D.; McKee, J.J.; O’Brien, T.M.; Robinson, W.F. The nucleotide sequence of koala (Phascolarctos cinereus) retrovirus: A novel type c endogenous virus related to gibbon ape leukemia virus. J. Virol. 2000, 74, 4264–4272. [Google Scholar] [CrossRef]
- Tarlinton, R.; Meers, J.; Young, P. Biology and evolution of the endogenous koala retrovirus. Cell. Mol. Life Sci. 2008, 65, 3413–3421. [Google Scholar] [CrossRef] [PubMed]
- Fiebig, U.; Hartmann, M.G.; Bannert, N.; Kurth, R.; Denner, J. Transspecies transmission of the endogenous koala retrovirus. J. Virol. 2006, 80, 5651–5654. [Google Scholar] [CrossRef] [PubMed]
- Simmons, G.S.; Young, P.R.; Hanger, J.J.; Jones, K.; Clarke, D.; McKee, J.J.; Meers, J. Prevalence of koala retrovirus in geographically diverse populations in australia. Aust. Vet. J. 2012, 90, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Tarlinton, R.E.; Meers, J.; Young, P.R. Retroviral invasion of the koala genome. Nature 2006, 442, 79–81. [Google Scholar] [CrossRef] [PubMed]
- Avila-Arcos, M.C.; Ho, S.Y.; Ishida, Y.; Nikolaidis, N.; Tsangaras, K.; Honig, K.; Medina, R.; Rasmussen, M.; Fordyce, S.L.; Calvignac-Spencer, S.; et al. One hundred twenty years of koala retrovirus evolution determined from museum skins. Mol. Biol. Evol. 2013, 30, 299–304. [Google Scholar] [CrossRef]
- Ishida, Y.; Zhao, K.; Greenwood, A.D.; Roca, A.L. Proliferation of endogenous retroviruses in the early stages of a host germ line invasion. Mol. Biol. Evol. 2015, 32, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Bolin, L.L.; Levy, L.S. Viral determinants of felv infection and pathogenesis: Lessons learned from analysis of a natural cohort. Viruses 2011, 3, 1681–1698. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, T.; Shojima, T.; Yoshikawa, R.; Ohata, T. Isolation of koala retroviruses from koalas in japan. J. Vet. Med. Sci. 2011, 73, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Logg, C.R.; Baranick, B.T.; Lemp, N.A.; Kasahara, N. Adaptive evolution of a tagged chimeric gammaretrovirus: Identification of novel cis-acting elements that modulate splicing. J. Mol. Biol. 2007, 369, 1214–1229. [Google Scholar] [CrossRef] [PubMed]
- Lander, M.R.; Chattopadhyay, S.K. A mus dunni cell line that lacks sequences closely related to endogenous murine leukemia viruses and can be infected by ectropic, amphotropic, xenotropic, and mink cell focus-forming viruses. J. Virol. 1984, 52, 695–698. [Google Scholar] [PubMed]
- Xu, W.; Stadler, C.K.; Gorman, K.; Jensen, N.; Kim, D.; Zheng, H.; Tang, S.; Switzer, W.M.; Pye, G.W.; Eiden, M.V. An exogenous retrovirus isolated from koalas with malignant neoplasias in a us zoo. Proc. Natl. Acad. Sci. USA 2013, 110, 11547–11552. [Google Scholar] [CrossRef] [PubMed]
- Hobbs, M.; Pavasovic, A.; King, A.G.; Prentis, P.J.; Eldridge, M.D.; Chen, Z.; Colgan, D.J.; Polkinghorne, A.; Wilkins, M.R.; Flanagan, C.; et al. A transcriptome resource for the koala (phascolarctos cinereus): Insights into koala retrovirus transcription and sequence diversity. BMC Genomics 2014, 15, e786. [Google Scholar] [CrossRef]
- Shojima, T.; Yoshikawa, R.; Hoshino, S.; Shimode, S.; Nakagawa, S.; Ohata, T.; Nakaoka, R.; Miyazawa, T. Identification of a novel subgroup of koala retrovirus from koalas in Japanese zoos. J. Virol. 2013, 87, 9943–9948. [Google Scholar] [CrossRef] [PubMed]
- Shimode, S.; Nakagawa, S.; Yoshikawa, R.; Shojima, T.; Miyazawa, T. Heterogeneity of koala retrovirus isolates. FEBS Lett. 2014, 588, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.G.; Miller, A.D. A family of retroviruses that utilize related phosphate transporters for cell entry. J. Virol. 1994, 68, 8270–8276. [Google Scholar] [PubMed]
- Van Zeijl, M.; Johann, S.V.; Closs, E.; Cunningham, J.; Eddy, R.; Shows, T.B.; O’Hara, B. A human amphotropic retrovirus receptor is a second member of the gibbon ape leukemia virus receptor family. Proc. Natl. Acad. Sci. USA 1994, 91, 1168–1172. [Google Scholar] [CrossRef]
- Yonezawa, A.; Masuda, S.; Katsura, T.; Inui, K. Identification and functional characterization of a novel human and rat riboflavin transporter, rft1. Am. J. Physiol. Cell Physiol. 2008, 295, C632–C641. [Google Scholar] [CrossRef] [PubMed]
- Marin, M.; Tailor, C.S.; Nouri, A.; Kabat, D. Sodium-dependent neutral amino acid transporter type 1 is an auxiliary receptor for baboon endogenous retrovirus. J. Virol. 2000, 74, 8085–8093. [Google Scholar] [CrossRef] [PubMed]
- Rasko, J.E.; Battini, J.L.; Gottschalk, R.J.; Mazo, I.; Miller, A.D. The rd114/simian type d retrovirus receptor is a neutral amino acid transporter. Proc. Natl. Acad. Sci. USA 1999, 96, 2129–2134. [Google Scholar] [CrossRef] [PubMed]
- Quigley, J.; Burns, C.; Anderson, M.; Lynch, E.; Sabo, K.; Overbaugh, J.; Abkowitz, J. Cloning of the cellular receptor for feline leukemia virus subgroup c (felv-c), a retrovirus that induces red cell aplasia. Blood 2000, 95, 1093–1099. [Google Scholar] [PubMed]
- Tailor, C.S.; Lavillette, D.; Marin, M.; Kabat, D. Cell surface receptors for gammaretroviruses. Curr. Top. Microbiol. Immunol. 2003, 281, 29–106. [Google Scholar] [PubMed]
- Bromham, L.D. The human zoo: Endogenous retroviruses in the human genome. Trends Ecol. Evol. 2002, 17, 91–97. [Google Scholar] [CrossRef]
- Tarlinton, R.; Meers, J.; Hanger, J.; Young, P. Real-time reverse transcriptase PCR for the endogenous koala retrovirus reveals an association between plasma viral load and neoplastic disease in koalas. J. Gen. Virol. 2005, 86, 783–787. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, W.; Gorman, K.; Santiago, J.C.; Kluska, K.; Eiden, M.V. Genetic Diversity of Koala Retroviral Envelopes. Viruses 2015, 7, 1258-1270. https://doi.org/10.3390/v7031258
Xu W, Gorman K, Santiago JC, Kluska K, Eiden MV. Genetic Diversity of Koala Retroviral Envelopes. Viruses. 2015; 7(3):1258-1270. https://doi.org/10.3390/v7031258
Chicago/Turabian StyleXu, Wenqin, Kristen Gorman, Jan Clement Santiago, Kristen Kluska, and Maribeth V. Eiden. 2015. "Genetic Diversity of Koala Retroviral Envelopes" Viruses 7, no. 3: 1258-1270. https://doi.org/10.3390/v7031258
APA StyleXu, W., Gorman, K., Santiago, J. C., Kluska, K., & Eiden, M. V. (2015). Genetic Diversity of Koala Retroviral Envelopes. Viruses, 7(3), 1258-1270. https://doi.org/10.3390/v7031258