
Insect-Specific Flaviviruses: A Systematic Review of Their Discovery, Host Range, Mode of Transmission, Superinfection Exclusion Potential and Genomic Organization


Abstract
:1. Introduction

2. Classical Insect-Specific Flaviviruses
2.1. Discovery, Geographic Distribution and Natural Host range

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| a Virus | Isolate Available | Geographic Distribution | Natural Host Range | References |
|---|---|---|---|---|
| Aedes flavivirus (AEFV) | Yes | Japan (2003), Italy (2008), USA (2011), b Thailand (2012) | Ae. albopictus, Ae. flavopictus, Cx. pipiens | [12,19,24,25,26,27] |
| Aedes galloisi flavivirus (AGFV) | Yes | Japan (2003) | Ae. galloisi | [28] |
| Calbertado virus (CLBOV) | Yes | Canada (2003), USA (2006) | Cx. tarsalis, Cx. pipiens | [29,30] |
| Cell fusing agent virus (CFAV) | Yes | Laboratory (1975), Puerto Rico (2002), Indonesia (2004), Mexico (2007), Thailand (2008), bUnited States (2012) | Ae. albopictus, Ae. aegypti, Culex spp. | [17,19,20,21,22,24] |
| Culex flavivirus (CxFV) | Yes | Japan (2003), Indonesia (2004), China (2006), Guatemala (2006), USA (2006), Mexico (2007), Trinidad (2008), Uganda (2008), Argentina (2009) | Cx. interrogator, Cx. maxi, Cx. nigripalpus, Cx. pipiens, Cx. quinquefasciatus, Cx. restuans, Cx. tarsalis, Cx. tritaeniorhynchus, Cx. usquatus | [29,31,32,33,34,35,36,37,38] |
| c Culex theileri flavivirus (CTFV) | Yes | Spain (2006), Portugal (2009–2010), Greece (2010), Thailand (date not specified) | Cx. fuscocephala, Cx. pipiens, Cx. theileri | [25,39,40,41] (Genbank Accession No. AY457040) |
| d Hanko virus (HANKV) | Yes | Finland (2005), Spain (2006), Italy (ca. 2007), Portugal (ca. 2007) | Ae. caspius, Ae. detritus, Ae. vexans, Cx. pipiens, Cx. perexiguus, Cx. theileri | [25,39,42,43] |
| Kamiti River virus (KRV) | Yes | Kenya (1999) | Ae. macintoshi | [44,45] |
| Nakiwogo virus (NAKV) | Yes | Uganda (2008) | Mansonia africana nigerrima | [38] |
| e Nienokoue virus(NIEV) | f Yes | Cote d’Ivoire (2004) | Culex spp. | (Genbank Accession No. NC_024299) |
| Palm Creek virus (PCV) | Yes | Australia (2010) | Coquillettidia xanthogaster | [46] |
| Quang Binh virus (QBV) | Yes | Vietnam (2002), China (2009) | An. sinensis, Cx. tritaeniorhynchus | [47,48,49] |
2.2. In Vitro and in Vivo Replication Potential in Vertebrates and Arthropod Cells
2.3. Transmission
2.4. Competitive Interaction between cISFs and Dual-Host Flaviviruses
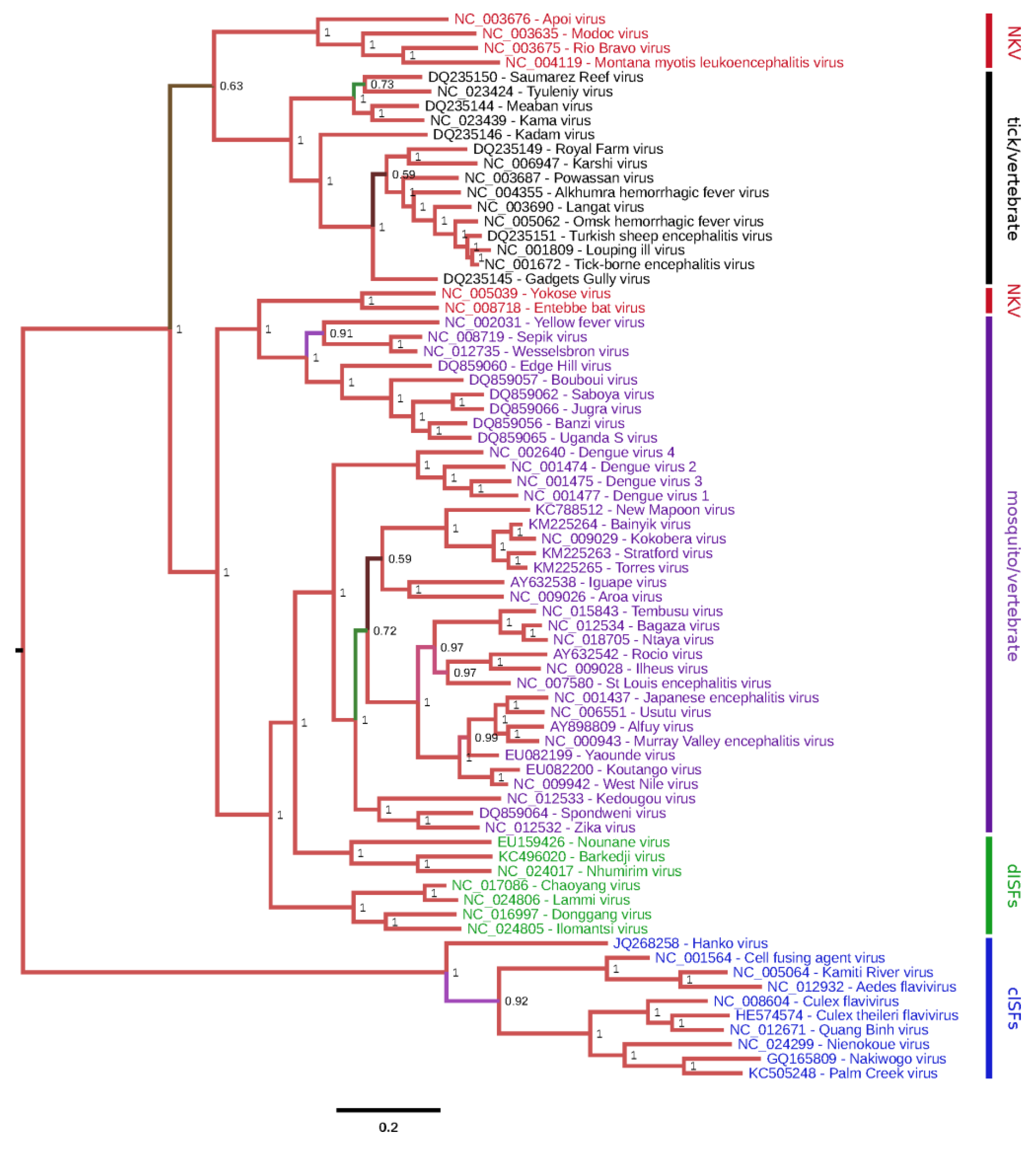
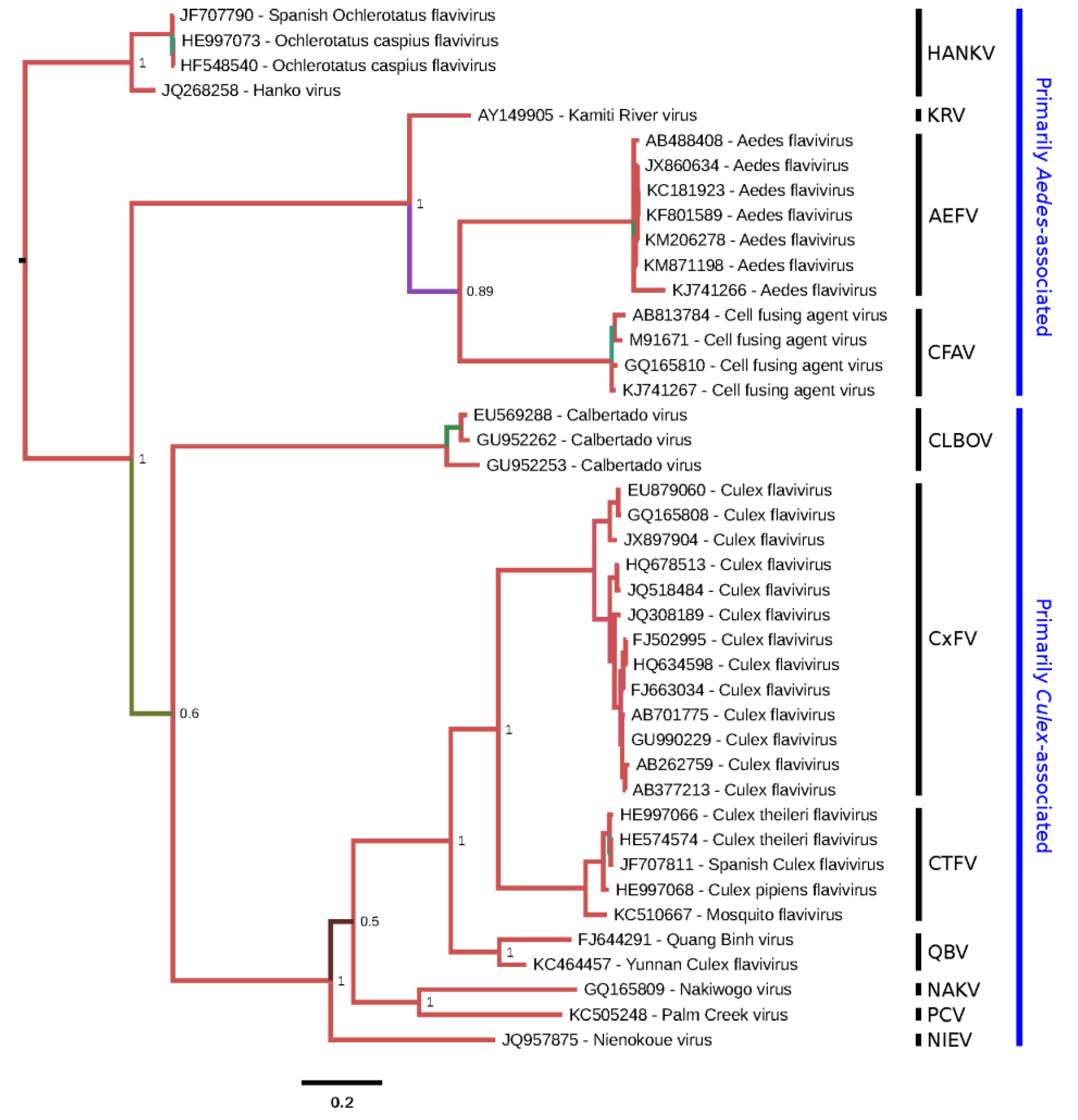
2.5. Genome Sequencing and Phylogeny
| Virus | Sequence Data Available | Length of Genome (nt) | Length of 5’ UTR (nt) | Length of 3’ UTR (nt) | a Genbank Accession No. |
|---|---|---|---|---|---|
| Aedes flavivirus | Genome | 11,064 | 96 | 945 | NC_012932 |
| Aedes galloisi flavivirus | Partial NS5 | b - | - | - | AB639347 |
| Calbertado virus | Partial NS5 | - | - | - | EU569288 |
| Cell fusing agent virus | Genome | 10,695 | 113 | 556 | NC_001564 |
| Culex flavivirus | Genome | 10,834 | 91 | 657 | NC_008604 |
| Culex theileri flavivirus | ORF | - | - | - | HE574574 |
| Hanko virus | ORF | - | - | - | JQ268258 |
| Kamiti River virus | Genome | 11,375 | 96 | 1205 | NC_005064 |
| Nakiwogo virus | ORF | - | - | - | GQ165809 |
| Nienokoue virus | ORF | - | - | - | NC_024299 |
| Palm Creek virus | ORF | - | - | - | KC505248 |
| Quang Binh virus | Genome | 10,865 | 112 | 673 | NC_012671 |

2.6. Ribosomal Frameshifting


| Junction | AEFV | CFAV | CxFV | CTFV | HANKV | KRV | NAKV | NIEV | PCV | QBV | Dual-Host Flaviviruses |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Virion C/Anch | b LEAQR↓SHSPV | c LESRR↓TTGNP | d LEAKR↓SAKNA | LEVRR↓SANNP | LEKER↓SHPRK | e LEKQR↓SGPNL | LEKRR↓GVWSP | LEQRR↓GAQRG | LEKKR↓DGRAA | LENRR↓SANPL | After dibasic residues |
| C/prM | b GLALS↓ETLRY | j VLCGC↓VVIDM | n MMVLG↓AVVID | VLCGC↓VIIDM | IVVTG↓LSIEL | e GLCYG↓EMLRY | VGIFS↓LNVVD | MVTFA↓AVVDV | FGVMG↓VVVID | TLCGT↓MVIDM | Signalase-like cleavage |
| pr/M | b PRKRR↓SSPQR | KREKR↓SREPP | d KRERR↓VASTN | KRVKR↓APETP | ERETR↓QKVDD | e VRRRR↓APQPQ | NRKQR↓SVKDE | RPVRR↓DVTPA | TRAKR↓VAPDG | KRVKR↓ATEQP | Furin |
| prM/E | b NVVRA↓TSIEP | j TTVKG↓EFVEP | d TTVKG↓EFVEP | TTVKG↓EFVEP | NVVKG↓EFVEP | e NVVKA↓SSIEP | TTVRG↓EFMEP | TTVSG↓EYLEP | TTVRG↓EYMEP | STVKG↓EFVEP | Signalase-like cleavage |
| E/NS1 | f RRVAG↓DIGCG | c YYVRA↓DLGCG | d VYTKA↓DVGCG | YFARA↓DVGCG | VYVKA↓DVGCG | e RSVSA↓DVGCG | YTVRA↓DFGCG | YYVRA↓DVGCG | YFVRA↓DFGCG | YYTRA↓DVGCG | Signalase-like cleavage |
| NS1/NS2A | b GKADA↓TADFH | c GKANA↓QSDFR | ° PPVEG↓SYPDF | PGTGA↓FPDFQ | YRVPS↓TNAED | e GKAHA↓CSDFR | PPSGA↓EKLQQ | GGAEA↓TQSFF | PMGET↓AKIQN | PGAEA↓LLQDF | Signalase-like cleavage |
| NS2A/NS2B | g KSSYR↓TSGRS | k RNGYR↓DSGAN | p RSGLR↓ASRRS | KSGLR↓ASKSS | RSGYR↓ALCSS | s KNGYR↓DYGAS | ASGLR↓KPRPH | KSGLR↓SITSW | GDGLR↓APRPH | KSGLR↓ASKRS | After dibasic residues |
| NS2B/NS3 | b NEHCR↓SDDLL | c TASNR↓SDDLL | q VSVFR↓SNEVN | STAYR↓AGVND | TNAFR↓SDELI | e SEQNR↓SDDLL | EFAQR↓SSSEL | STAQR↓SDLLL | AMSQR↓ANSEL | TSNRR↓SGVND | After dibasic residues |
| NS3/NS4A | h YINTR↓SSASL | l YMNCR↓GGPTL | r YLKQR↓SNFNF | FLKQR↓SGANF | YMGTR↓SFLSV | t YLNCR↓SSQTF | FLKQR↓SVLPF | FLKQR↓SLFID | FLKQR↓SLYFD | FLKQR↓SVLNF | After dibasic residues |
| NS4A/2K | AAGNR↓SYLDS | SIGNR↓SYMDS | NNVHR↓AYTTD | NNVHR↓AYTGD | SAGQR↓SYVDI | AIGNR↓SYMDS | GGSQR↓GILDS | ANSQR↓GFAEN | GGSQR↓GVLDS | TNVHR↓AYTGD | After dibasic residues |
| 2K/NS4B | b CSVLA↓WEMRL | c CGVLA↓WEMRM | d MGVVA↓WEMDL | MGVVA↓WELNL | IGVIC↓WELRL | e CGVLA↓WEMRL | IGIAA↓WELQL | SAVVA↓WELNL | IGVTA↓WELEL | MGIVA↓WELEL | Signalase-like cleavage |
| NS4B/NS5 | i FSKFR↓ALEKS | m FNQFR↓ALEKS | dRMALR↓SLVKT | RGGLR↓SLVKT | NITTR↓SLEKS | u FNQFR↓ALEKS | RLSVR↓SLVKS | LDMRR↓SLMKT | RLGVR↓SLVKS | RLATR↓SLVKT | After dibasic residues |
2.7. Predicted Polyprotein Cleavage Sites
3. Dual-Host Affiliated Insect-Specific Flaviviruses
3.1. Discovery, Geographic Distribution and Natural Host Range
| Virus | Isolate Available | Geographic Distribution | Natural Host Range | References |
|---|---|---|---|---|
| Barkedji virus (BJV) | No | Senegal (date not reported), Israel (2011) | Cx. perexiguus | [101], (Genbank Accession No. EU078325) |
| Chaoyang virus (CHAOV) | Yes | China (2008), South Korea (2003) | Ae. vexans, Ae. albopictus, Ae. bekkui, Armigeres subalbatus, Cx. pipiens | [102,103,104,111] |
| Donggang virus (DONV) | a Yes | China (2009) | Aedes spp. | (Genbank Accession No. NC_016997) |
| Ilomantsi virus (ILOV) | Yes | Finland (2007) | Most likely Oc. riparius and/or Anopheles spp. | [105] |
| Lammi virus (LAMV) | Yes | Finland (2004) | Ae. cinereus | [106] |
| Marisma mosquito virus (MMV) | Yes | Spain (2003), Italy (2011) | Ae. caspius | [39,107] |
| Nanay virus (NANV) | Yes | Peru (2009) | Culex (Melanoconion) ocossa | [108] |
| Nhumirim virus (NHUV) | Yes | Brazil (2010) | Cx. chidesteri | [109] |
| Nounané virus (NOUV) | Yes | Côte d'Ivoire (2004) | Uranotaenia mashonaensis | [100] |
3.2. In Vitro and in Vivo Replication Potential in Vertebrates and Arthropods
3.3. Transmission
3.4. Competitive Interaction between dISFs and Dual-Host Flaviviruses
3.5. Genome Sequencing and Phylogeny
| Virus | Sequence Data Available | Length of Genome (nt) | Length of 5’ UTR (nt) | Length of 3’ UTR (nt) | a Genbank Accession No. |
|---|---|---|---|---|---|
| Barkedji virus | Almost entire ORF | b - | - | - | KC496020 |
| Chaoyang virus | Genome | 10,733 | 99 | 326 | NC_017086 |
| Donggang virus | Genome | 10,791 | 113 | 343 | NC_016997 |
| Ilomantsi virus | ORF | - | - | - | NC_024805 |
| Lammi virus | ORF | - | - | - | KC692068 |
| Marisma mosquito virus | Partial NS5 | - | - | - | JN603190 |
| Nanay virus | Partial E and NS5 | - | - | - | JX627335 |
| Nhumirim virus | Genome | 10,891 | 102 | 451 | NC_024017 |
| Nounané virus | ORF | - | - | - | EU159426 |

3.6. Ribosomal Frameshifting
| Junction | BJV | CHAOV | DONV | ILOV | LAMV | NHUV | NOUV | Dual-Host Flaviviruses |
|---|---|---|---|---|---|---|---|---|
| Virion C/Anch | b KTSKR↓GLQQS | RKAKR↓SVTTP | RPNRR↓SAGSN | QKTRR↓SVDTV | KNGKR↓SKTEI | c RRARR↓GMGIP | d VSKRR↓GSASL | After dibasic residues |
| C/prM | b TMAAC↓ATLGM | CMAYG↓ATRFT | GTAMA↓ATSMT | VAVIA↓TTVTT | GTAMA↓ASMFT | b TMVAC↓VTVGT | d GVASA↓VTFTT | Signalase-like cleavage |
| pr/M | b RRSKR↓SVAIA | RRSRR↓SVALA | RRSRR↓SIMIP | RRSRR↓SIALA | RRGKR↓SVALA | b RRSRR↓SVALS | d QRSRR↓SVGIS | Furin |
| prM/E | b APAYS↓LHCSR | GPAYS↓LQCID | APVYG↓SQCSG | APVYG↓HHCSG | GPAYS↓LQCVD | b APAYS↓THCVR | d IPAYS↓MKCIG | Signalase-like cleavage |
| E/NS1 | b TTVAG↓DVGCN | TVGVS↓EIGCS | TNAVS↓EVGCS | SAAAS↓EVGCS | TVALS↓EVGCS | b TSAHA↓EVGCS | c TSVSA↓ELGCS | Signalase-like cleavage |
| NS1/NS2A | b SWTTA↓GNATG | SKVSA↓GTFQG | ARVSA↓GAVHG | ARVSA↓GLVAG | SKVSA↓GTFQG | b SWVTA↓GQMTG | e SWVSA↓GEPMV | Signalase-like cleavage |
| NS2A/NS2B | b GSGKR↓SVSMG | SSGKR↓SWPAG | KHGKR↓SWPAG | RNGRR↓SWPAG | TSGKR↓SWPAG | b KSGKR↓SVSMG | d KTTKR↓SVPQS | After dibasic residues |
| NS2B/NS3 | b KGTQK↓AGAMW | KSGRR↓GTVLW | KHDRR↓GGVLW | RTAKR↓GGVLW | KSGRR↓GTVLW | b SATQR↓AGAMW | d ENRKR↓SNDTP | After dibasic residues |
| NS3/NS4A | b AEGRR↓GASDI | AEGRR↓SYVPI | AEGRR↓SYMPI | AEGKR↓SAVQL | AEGRR↓SYVPL | b AEGRR↓GAMDL | d AGGKR↓SAVDL | After dibasic residues |
| NS4A/2K | AEKQR↓SAIDN | PGSQR↓SVQDN | PGNQR↓SIQDN | AGGQR↓SIADN | PGSQR↓SVQDN | AEKQR↓SALDN | d EGKQR↓SMVDN | After dibasic residues |
| 2K/NS4B | b LAVTA↓NEKGL | ALIAA↓NETGL | GGIAA↓NEMGM | SLIAA↓NETGL | ALIAA↓NETGL | b LMIAA↓NEKGL | d GAVAA↓NEYGM | Signalase-like cleavage |
| NS4B/NS5 | b KSARK↓GTPGG | GVPRR↓GVTIS | QPSRR↓GKKVE | TTPRR↓GRRVN | GVPRR↓GMTIC | b KSARR↓GTPGG | f VVTRK↓GTAGG | After dibasic residues |
3.7. Predicted Polyprotein Cleavage Sites
4. Closing Remarks and Future Research
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lindenbach, B.D.; Thiel, H.-J.; Rice, C.M. Flaviviridae: The viruses and their replication. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2007; pp. 1101–1152. [Google Scholar]
- Markoff, L. 5'- and 3'-noncoding regions in flavivirus RNA. Adv. Virus Res. 2003, 59, 177–228. [Google Scholar] [PubMed]
- Rice, C.M.; Lenches, E.M.; Eddy, S.R.; Shin, S.J.; Sheets, R.L.; Strauss, J.H. Nucleotide sequence of yellow fever virus: implications for flavivirus gene expression and evolution. Science 1985, 229, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Gubler, D.J.; Kuno, G.; Markoff, L. Flaviviruses. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2007; pp. 1153–1252. [Google Scholar]
- Johnson, H.N. Ecological implications of antigenically related mammalian viruses for which arthropod vectors are unknown and avian associated soft tick viruses. Jpn. J. Med. Sci. Biol. 1967, 20 (Suppl), 160–166. [Google Scholar] [PubMed]
- Burns, K.F.; Farinacci, C.J. Virus of bats antigenically related to St. Louis encephalitis. Science 1956, 123, 227. [Google Scholar] [CrossRef] [PubMed]
- Fairbrother, A.; Yuill, T.M. Experimental infection and horizontal transmission of Modoc virus in deer mice (Peromyscus maniculatus). J. Wildl. Dis. 1987, 23, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Adams, A.P.; Travassos da Rosa, A.P.; Nunes, M.R.; Xiao, S.Y.; Tesh, R.B. Pathogenesis of Modoc virus (Flaviviridae; Flavivirus) in persistently infected hamsters. Am. J. Trop. Med. Hyg. 2013, 88, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.F.; Thomas, L.A. A New Virus, "Mml", Enzootic in Bats (Myotis Lucifugus) of Montana. Am. J. Trop. Med. Hyg. 1964, 13, 607–612. [Google Scholar] [PubMed]
- Constantine, D.G.; Woodall, D.F. Latent Infection of Rio Bravo Virus in Salivary Glands of Bats. Public Health Rep. 1964, 79, 1033–1039. [Google Scholar] [CrossRef] [PubMed]
- Crochu, S.; Cook, S.; Attoui, H.; Charrel, R.N.; de Chesse, R.; Belhouchet, M.; Lemasson, J.J.; de Micco, P.; de Lamballerie, X. Sequences of flavivirus-related RNA viruses persist in DNA form integrated in the genome of Aedes spp. mosquitoes. J. Gen. Virol. 2004, 85, 1971–1980. [Google Scholar] [CrossRef] [PubMed]
- Roiz, D.; Vazquez, A.; Rosso, F.; Arnoldi, D.; Girardi, M.; Cuevas, L.; Perez-Pastrana, E.; Sanchez-Seco, M.P.; Tenorio, A.; Rizzoli, A. Detection of a new insect flavivirus and isolation of Aedes flavivirus in Northern Italy. Parasit. Vectors. 2012, 5, e223. [Google Scholar] [CrossRef]
- Katzourakis, A.; Gifford, R.J. Endogenous viral elements in animal genomes. PLOS Genet. 2010, 6, e1001191. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: a multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, e113. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Stollar, V.; Thomas, V.L. An agent in the Aedes aegypti cell line (Peleg) which causes fusion of Aedes albopictus cells. Virology 1975, 64, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Cammisa-Parks, H.; Cisar, L.A.; Kane, A.; Stollar, V. The complete nucleotide sequence of cell fusing agent (CFA): Homology between the nonstructural proteins encoded by CFA and the nonstructural proteins encoded by arthropod-borne flaviviruses. Virology 1992, 189, 511–524. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, K.; Isawa, H.; Tsuda, Y.; Sawabe, K.; Kobayashi, M. Isolation and characterization of a new insect flavivirus from Aedes albopictus and Aedes flavopictus mosquitoes in Japan. Virology 2009, 391, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Espinoza-Gomez, F.; Lopez-Lemus, A.U.; Rodriguez-Sanchez, I.P.; Martinez-Fierro, M.L.; Newton-Sanchez, O.A.; Chavez-Flores, E.; Delgado-Enciso, I. Detection of sequences from a potentially novel strain of cell fusing agent virus in Mexican Stegomyia (Aedes) aegypti mosquitoes. Arch. Virol. 2011, 156, 1263–1267. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.; Bennett, S.N.; Holmes, E.C.; de Chesse, R.; Moureau, G.; de Lamballerie, X. Isolation of a new strain of the flavivirus cell fusing agent virus in a natural mosquito population from Puerto Rico. J. Gen. Virol. 2006, 87, 735–748. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, A.; Thongrungkiat, S.; Ramasoota, P.; Konishi, E. Genetic and evolutionary analysis of cell-fusing agent virus based on Thai strains isolated in 2008 and 2012. Infect. Genet. Evol. 2013, 19, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Kihara, Y.; Satho, T.; Eshita, Y.; Sakai, K.; Kotaki, A.; Takasaki, T.; Rongsriyam, Y.; Komalamisra, N.; Srisawat, R.; Lapcharoen, P.; et al. Rapid determination of viral RNA sequences in mosquitoes collected in the field. J. Virol. Methods. 2007, 146, 372–374. [Google Scholar] [CrossRef] [PubMed]
- Bolling, B.G.; Vasilakis, N.; Guzman, H.; Widen, S.G.; Wood, T.G.; Popov, V.L.; Thangamani, S.; Tesh, R.B. Insect-Specific Viruses Detected in Laboratory Mosquito Colonies and Their Potential Implications for Experiments Evaluating Arbovirus Vector Competence. Am. J. Trop. Med. Hyg. 2014. [Google Scholar] [CrossRef]
- Calzolari, M.; Ze-Ze, L.; Ruzek, D.; Vazquez, A.; Jeffries, C.; Defilippo, F.; Osorio, H.C.; Kilian, P.; Ruiz, S.; Fooks, A.R.; et al. Detection of mosquito-only flaviviruses in Europe. J. Gen. Virol. 2012, 93, 1215–1225. [Google Scholar] [CrossRef] [PubMed]
- Haddow, A.D.; Guzman, H.; Popov, V.L.; Wood, T.G.; Widen, S.G.; Haddow, A.D.; Tesh, R.B.; Weaver, S.C. First isolation of Aedes flavivirus in the Western Hemisphere and evidence of vertical transmission in the mosquito Aedes (Stegomyia) albopictus (Diptera: Culicidae). Virology 2013, 440, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Grisenti, M.; Vazquez, A.; Herrero, L.; Cuevas, L.; Perez-Pastrana, E.; Arnoldi, D.; Rosa, R.; Capelli, G.; Tenorio, A.; Sanchez-Seco, M.P.; Rizzoli, A. Wide detection of Aedes flavivirus in north-eastern Italy, a European hot-spot of emerging mosquito-borne diseases. J. Gen. Virol. 2015, 96, 420–430. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, K.; Takahashi-Nakaguchi, A.; Isawa, H.; Sasaki, T.; Higa, Y.; Kasai, S.; Tsuda, Y.; Sawabe, K.; Kobayashi, M. Entomological surveillance for flaviviruses at migratory bird stopover sites in Hokkaido, Japan, and a new insect flavivirus detected in Aedes galloisi (Diptera: Culicidae). J. Med. Entomol. 2012, 49, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Bolling, B.G.; Eisen, L.; Moore, C.G.; Blair, C.D. Insect-specific flaviviruses from Culex mosquitoes in Colorado, with evidence of vertical transmission. Am. J. Trop. Med. Hyg. 2011, 85, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Tyler, S.; Bolling, B.G.; Blair, C.D.; Brault, A.C.; Pabbaraju, K.; Armijos, M.V.; Clark, D.C.; Calisher, C.H.; Drebot, M.A. Distribution and phylogenetic comparisons of a novel mosquito flavivirus sequence present in Culex tarsalis Mosquitoes from western Canada with viruses isolated in California and Colorado. Am. J. Trop. Med. Hyg. 2011, 85, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, K.; Isawa, H.; Tsuda, Y.; Yano, K.; Sasaki, T.; Yuda, M.; Takasaki, T.; Kobayashi, M.; Sawabe, K. Genetic characterization of a new insect flavivirus isolated from Culex pipiens mosquito in Japan. Virology 2007, 359, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Goenaga, S.; Fabbri, C.M.; Garcia, J.B.; Rondan, J.C.; Gardenal, N.; Calderon, G.E.; Enria, D.A.; Levis, S.M. New strains of Culex flavivirus isolated in Argentina. J. Med. Entomol. 2014, 51, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Huanyu, W.; Haiyan, W.; Shihong, F.; Guifang, L.; Hong, L.; Xiaoyan, G.; Lizhi, S.; Rayner, S.; Aiqiang, X.; Guodong, L. Isolation and identification of a distinct strain of Culex Flavivirus from mosquitoes collected in Mainland China. Virol. J. 2012, 9, e73. [Google Scholar] [CrossRef]
- Morales-Betoulle, M.E.; Monzon Pineda, M.L.; Sosa, S.M.; Panella, N.; Lopez, M.R.; Cordon-Rosales, C.; Komar, N.; Powers, A.; Johnson, B.W. Culex flavivirus isolates from mosquitoes in Guatemala. J. Med. Entomol. 2008, 45, 1187–1190. [Google Scholar] [CrossRef] [PubMed]
- Farfan-Ale, J.A.; Lorono-Pino, M.A.; Garcia-Rejon, J.E.; Hovav, E.; Powers, A.M.; Lin, M.; Dorman, K.S.; Platt, K.B.; Bartholomay, L.C.; Soto, V.; Beaty, B.J.; Lanciotti, R.S.; Blitvich, B.J. Detection of RNA from a novel West Nile-like virus and high prevalence of an insect-specific flavivirus in mosquitoes in the Yucatan Peninsula of Mexico. Am. J. Trop. Med. Hyg. 2009, 80, 85–95. [Google Scholar] [PubMed]
- Saiyasombat, R.; Dorman, K.S.; Garcia-Rejon, J.E.; Lorono-Pino, M.A.; Farfan-Ale, J.A.; Blitvich, B.J. Isolation and sequence analysis of Culex flavivirus from Culex interrogator and Culex quinquefasciatus in the Yucatan Peninsula of Mexico. Arch. Virol. 2010, 155, 983–986. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Guzman, H.; Bueno, R., Jr.; Dennett, J.A.; Auguste, A.J.; Carrington, C.V.; Popov, V.L.; Weaver, S.C.; Beasley, D.W.; Tesh, R.B. Characterization of Culex Flavivirus (Flaviviridae) strains isolated from mosquitoes in the United States and Trinidad. Virology 2009, 386, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.; Moureau, G.; Harbach, R.E.; Mukwaya, L.; Goodger, K.; Ssenfuka, F.; Gould, E.; Holmes, E.C.; de Lamballerie, X. Isolation of a novel species of flavivirus and a new strain of Culex flavivirus (Flaviviridae) from a natural mosquito population in Uganda. J. Gen. Virol. 2009, 90, 2669–2678. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, A.; Sanchez-Seco, M.P.; Palacios, G.; Molero, F.; Reyes, N.; Ruiz, S.; Aranda, C.; Marques, E.; Escosa, R.; Moreno, J.; et al. Novel flaviviruses detected in different species of mosquitoes in Spain. Vector Borne Zoonotic Dis. 2012, 12, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Parreira, R.; Cook, S.; Lopes, A.; de Matos, A.P.; de Almeida, A.P.; Piedade, J.; Esteves, A. Genetic characterization of an insect-specific flavivirus isolated from Culex theileri mosquitoes collected in southern Portugal. Virus Res. 2012, 167, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Papa, A.; Papadopoulou, E.; Kalaitzopoulou, S.; Tsioka, K.; Mourelatos, S. Detection of West Nile virus and insect-specific flavivirus RNA in Culex mosquitoes, central Macedonia, Greece. Trans. R. Soc. Trop. Med. Hyg. 2014, 108, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Huhtamo, E.; Moureau, G.; Cook, S.; Julkunen, O.; Putkuri, N.; Kurkela, S.; Uzcategui, N.Y.; Harbach, R.E.; Gould, E.A.; Vapalahti, O.; de Lamballerie, X. Novel insect-specific flavivirus isolated from northern Europe. Virology 2012, 433, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, D.D.; Cook, S.; Lopes, A.; de Matos, A.P.; Esteves, A.; Abecasis, A.; de Almeida, A.P.; Piedade, J.; Parreira, R. Characterization of an insect-specific flavivirus (OCFVPT) co-isolated from Ochlerotatus caspius collected in southern Portugal along with a putative new Negev-like virus. Virus Genes 2013, 47, 532–545. [Google Scholar] [CrossRef] [PubMed]
- Crabtree, M.B.; Sang, R.C.; Stollar, V.; Dunster, L.M.; Miller, B.R. Genetic and phenotypic characterization of the newly described insect flavivirus, Kamiti River virus. Arch. Virol. 2003, 148, 1095–1118. [Google Scholar] [CrossRef] [PubMed]
- Sang, R.C.; Gichogo, A.; Gachoya, J.; Dunster, M.D.; Ofula, V.; Hunt, A.R.; Crabtree, M.B.; Miller, B.R.; Dunster, L.M. Isolation of a new flavivirus related to cell fusing agent virus (CFAV) from field-collected flood-water Aedes mosquitoes sampled from a dambo in central Kenya. Arch. Virol. 2003, 148, 1085–1093. [Google Scholar] [CrossRef] [PubMed]
- Hobson-Peters, J.; Yam, A.W.; Lu, J.W.; Setoh, Y.X.; May, F.J.; Kurucz, N.; Walsh, S.; Prow, N.A.; Davis, S.S.; Weir, R.; et al. A new insect-specific flavivirus from northern Australia suppresses replication of West Nile virus and Murray Valley encephalitis virus in co-infected mosquito cells. PLOS ONE 2013, 8, e56534. [Google Scholar] [CrossRef] [PubMed]
- Crabtree, M.B.; Nga, P.T.; Miller, B.R. Isolation and characterization of a new mosquito flavivirus, Quang Binh virus, from Vietnam. Arch. Virol. 2009, 154, 857–860. [Google Scholar] [CrossRef] [PubMed]
- Zuo, S.; Zhao, Q.; Guo, X.; Zhou, H.; Cao, W.; Zhang, J. Detection of Quang Binh virus from mosquitoes in China. Virus Res. 2014, 180, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Li, H.B.; Zhu, J.; Zhang, Y.Z.; Yang, W.H.; Fan, J.H.; Liang, G.D.; Zhang, H.L. First isolation of quang binh-like virus from mosquitoes in China. Bing Du Xue Bao 2014, 30, 57–61. [Google Scholar] [PubMed]
- Machado, D.C.; Mondini, A.; dos Santos Santana, V.; Yonamine, P.T.; Chiaravalloti Neto, F.; Zanotto, P.M.; Nogueira, M.L. First identification of Culex flavivirus (Flaviviridae) in Brazil. Intervirology 2012, 55, 475–483. [Google Scholar] [CrossRef] [PubMed]
- An, S.Y.; Liu, J.S.; Ren, Y.; Wang, Z.S.; Han, Y.; Ding, J.; Guo, J.Q. Isolation of the Culex flavivirus from mosquitoes in Liaoning Province, China. Bing Du Xue Bao 2012, 28, 511–516. [Google Scholar] [PubMed]
- Farfan-Ale, J.A.; Lorono-Pino, M.A.; Garcia-Rejon, J.E.; Soto, V.; Lin, M.; Staley, M.; Dorman, K.S.; Bartholomay, L.C.; Hovav, E.; Blitvich, B.J. Detection of flaviviruses and orthobunyaviruses in mosquitoes in the Yucatan Peninsula of Mexico in 2008. Vector Borne Zoonotic Dis. 2010, 10, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Lin, J.W.; Fan, Y.C.; Tu, W.C.; Chang, G.J.; Chiou, S.S. First detection of the Africa/Caribbean/Latin American subtype of Culex flavivirus in Asian country, Taiwan. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Blitvich, B.J.; Lin, M.; Dorman, K.S.; Soto, V.; Hovav, E.; Tucker, B.J.; Staley, M.; Platt, K.B.; Bartholomay, L.C. Genomic sequence and phylogenetic analysis of Culex flavivirus, an insect-specific flavivirus, isolated from Culex pipiens (Diptera: Culicidae) in Iowa. J. Med. Entomol. 2009, 46, 934–941. [Google Scholar] [CrossRef] [PubMed]
- Crockett, R.K.; Burkhalter, K.; Mead, D.; Kelly, R.; Brown, J.; Varnado, W.; Roy, A.; Horiuchi, K.; Biggerstaff, B.J.; Miller, B.; et al. Culex flavivirus and West Nile virus in Culex quinquefasciatus populations in the southeastern United States. J. Med. Entomol. 2012, 49, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Newman, C.M.; Cerutti, F.; Anderson, T.K.; Hamer, G.L.; Walker, E.D.; Kitron, U.D.; Ruiz, M.O.; Brawn, J.D.; Goldberg, T.L. Culex flavivirus and West Nile virus mosquito coinfection and positive ecological association in Chicago, United States. Vector Borne Zoonotic Dis. 2011, 11, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Calzolari, M.; Bonilauri, P.; Bellini, R.; Caimi, M.; Defilippo, F.; Maioli, G.; Albieri, A.; Medici, A.; Veronesi, R.; Pilani, R.; et al. Arboviral survey of mosquitoes in two northern Italian regions in 2007 and 2008. Vector Borne Zoonotic Dis. 2010, 10, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Pabbaraju, K.; Ho, K.C.; Wong, S.; Fox, J.D.; Kaplen, B.; Tyler, S.; Drebot, M.; Tilley, P.A. Surveillance of mosquito-borne viruses in Alberta using reverse transcription polymerase chain reaction with generic primers. J. Med. Entomol. 2009, 46, 640–648. [Google Scholar] [CrossRef] [PubMed]
- Kuno, G.; Chang, G.J.; Tsuchiya, K.R.; Karabatsos, N.; Cropp, C.B. Phylogeny of the genus Flavivirus. J. Virol. 1998, 72, 73–83. [Google Scholar] [PubMed]
- Moureau, G.; Ninove, L.; Izri, A.; Cook, S.; de Lamballerie, X.; Charrel, R.N. Flavivirus RNA in phlebotomine sandflies. Vector Borne Zoonotic Dis. 2010, 10, 195–197. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Seco, M.P.; Vazquez, A.; Collao, X.; Hernandez, L.; Aranda, C.; Ruiz, S.; Escosa, R.; Marques, E.; Bustillo, M.A.; Molero, F.; et al. Surveillance of arboviruses in Spanish wetlands: detection of new flavi- and phleboviruses. Vector Borne Zoonotic Dis. 2010, 10, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.; Chung, B.Y.; Bass, D.; Moureau, G.; Tang, S.; McAlister, E.; Culverwell, C.L.; Glucksman, E.; Wang, H.; Brown, T.D.; et al. Novel virus discovery and genome reconstruction from field RNA samples reveals highly divergent viruses in dipteran hosts. PLOS One 2013, 8, e80720. [Google Scholar] [CrossRef] [PubMed]
- Kent, R.J.; Crabtree, M.B.; Miller, B.R. Transmission of West Nile virus by Culex quinquefasciatus say infected with Culex Flavivirus Izabal. PLOS Negl. Trop. Dis. 2010, 4, e671. [Google Scholar] [CrossRef] [PubMed]
- Saiyasombat, R.; Bolling, B.G.; Brault, A.C.; Bartholomay, L.C.; Blitvich, B.J. Evidence of efficient transovarial transmission of Culex flavivirus by Culex pipiens (Diptera: Culicidae). J. Med. Entomol. 2011, 48, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Bolling, B.G.; Olea-Popelka, F.J.; Eisen, L.; Moore, C.G.; Blair, C.D. Transmission dynamics of an insect-specific flavivirus in a naturally infected Culex pipiens laboratory colony and effects of co-infection on vector competence for West Nile virus. Virology 2012, 427, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.R.P. Cell Cultures Derived from Larvae of Aedes albopictus (Skuse) and Aedes aegypti (L). Curr. Sci. India 1967, 36, 506. [Google Scholar]
- Rosen, L.; Tesh, R.B.; Lien, J.C.; Cross, J.H. Transovarial transmission of Japanese encephalitis virus by mosquitoes. Science 1978, 199, 909–911. [Google Scholar] [CrossRef] [PubMed]
- Aitken, T.H.; Tesh, R.B.; Beaty, B.J.; Rosen, L. Transovarial transmission of yellow fever virus by mosquitoes (Aedes aegypti). Am. J. Trop. Med. Hyg. 1979, 28, 119–121. [Google Scholar] [PubMed]
- Tesh, R.B. Experimental studies on the transovarial transmission of Kunjin and San Angelo viruses in mosquitoes. Am. J. Trop. Med. Hyg. 1980, 29, 657–666. [Google Scholar] [PubMed]
- Lutomiah, J.J.; Mwandawiro, C.; Magambo, J.; Sang, R.C. Infection and vertical transmission of Kamiti river virus in laboratory bred Aedes aegypti mosquitoes. J. Insect Sci. 2007, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Nasar, F.; Haddow, A.D.; Tesh, R.B.; Weaver, S.C. Eilat virus displays a narrow mosquito vector range. Parasit. Vectors 2014, 7, e595. [Google Scholar] [CrossRef]
- Barbanti-Brodano, G.; Swetly, P.; Koprowski, H. Superinfection of simian virus 40-transformed permissive cells with simian virus 40. J. Virol. 1970, 6, 644–651. [Google Scholar] [PubMed]
- Burivong, P.; Pattanakitsakul, S.N.; Thongrungkiat, S.; Malasit, P.; Flegel, T.W. Markedly reduced severity of Dengue virus infection in mosquito cell cultures persistently infected with Aedes albopictus densovirus (AalDNV). Virology 2004, 329, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Pepin, K.M.; Lambeth, K.; Hanley, K.A. Asymmetric competitive suppression between strains of dengue virus. BMC Microbiol. 2008, 8, e28. [Google Scholar] [CrossRef]
- Sundin, D.R.; Beaty, B.J. Interference to oral superinfection of Aedes triseriatus infected with La Crosse virus. Am. J. Trop Med. Hyg. 1988, 38, 428–432. [Google Scholar] [PubMed]
- Tscherne, D.M.; Evans, M.J.; von Hahn, T.; Jones, C.T.; Stamataki, Z.; McKeating, J.A.; Lindenbach, B.D.; Rice, C.M. Superinfection exclusion in cells infected with hepatitis C virus. J. Virol. 2007, 81, 3693–3703. [Google Scholar] [CrossRef] [PubMed]
- Kuwata, R.; Isawa, H.; Hoshino, K.; Sasaki, T.; Kobayashi, M.; Maeda, K.; Sawabe, K. Analysis of mosquito-borne flavivirus superinfection in Culex tritaeniorhynchus (Diptera: Culicidae) cells persistently infected with Culex flavivirus (Flaviviridae). J. Med. Entomol. 2015. [Google Scholar] [CrossRef]
- Gritsun, T.S.; Gould, E.A. The 3' untranslated regions of Kamiti River virus and Cell fusing agent virus originated by self-duplication. J. Gen. Virol. 2006, 87, 2615–2619. [Google Scholar] [CrossRef] [PubMed]
- Kenney, J.L.; Solberg, O.D.; Langevin, S.A.; Brault, A.C. Characterization of a novel insect-specific flavivirus from Brazil: potential for inhibition of infection of arthropod cells with medically important flaviviruses. J. Gen. Virol. 2014, 95, 2796–2808. [Google Scholar] [CrossRef] [PubMed]
- Lobo, F.P.; Mota, B.E.; Pena, S.D.; Azevedo, V.; Macedo, A.M.; Tauch, A.; Machado, C.R.; Franco, G.R. Virus-Host coevolution: Common patterns of nucleotide motif usage in Flaviviridae and their hosts. PLOS ONE 2009, 4, e6282. [Google Scholar] [CrossRef] [PubMed]
- Greenbaum, B.D.; Levine, A.J.; Bhanot, G.; Rabadan, R. Patterns of evolution and host gene mimicry in influenza and other RNA viruses. PLOS Pathog. 2008, 4, e1000079. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Zhou, T.; Ma, J.; Sun, X.; Lu, Z. Analysis of synonymous codon usage in SARS Coronavirus and other viruses in the Nidovirales. Virus Res. 2004, 101, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, G.M.; Pagel, M.; Gould, E.A.; de Andrade Zanotto, P.M.; Holmes, E.C. Evolution of base composition and codon usage bias in the genus Flavivirus. J. Mol. Evol. 2001, 52, 383–390. [Google Scholar] [PubMed]
- Tulloch, F.; Atkinson, N.J.; Evans, D.J.; Ryan, M.D.; Simmonds, P. RNA virus attenuation by codon pair deoptimisation is an artefact of increases in CpG/UpA dinucleotide frequencies. eLife 2014, 4, e04531. [Google Scholar]
- Simmen, M.W. Genome-scale relationships between cytosine methylation and dinucleotide abundances in animals. Genomics 2008, 92, 33–40. [Google Scholar] [CrossRef] [PubMed]
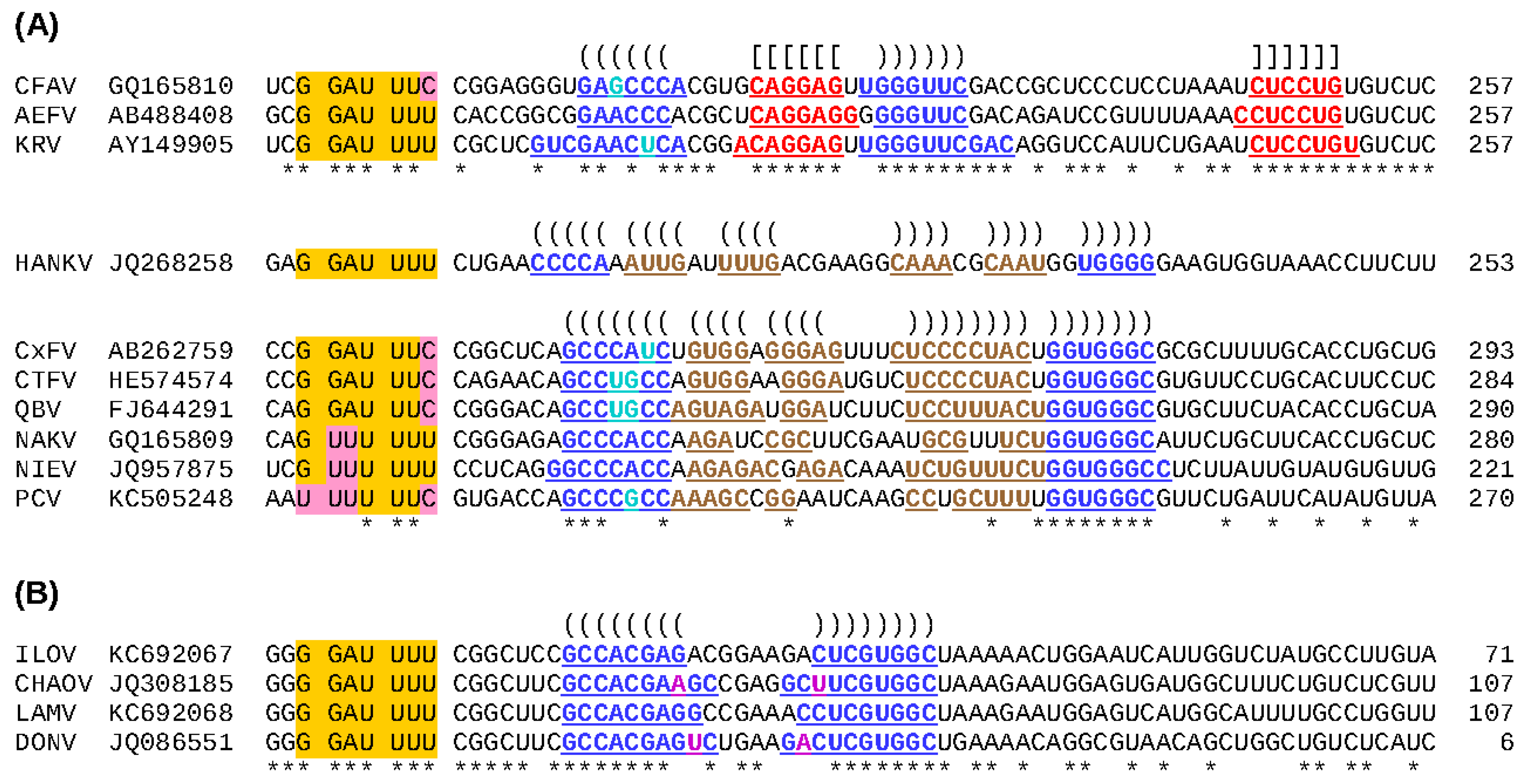
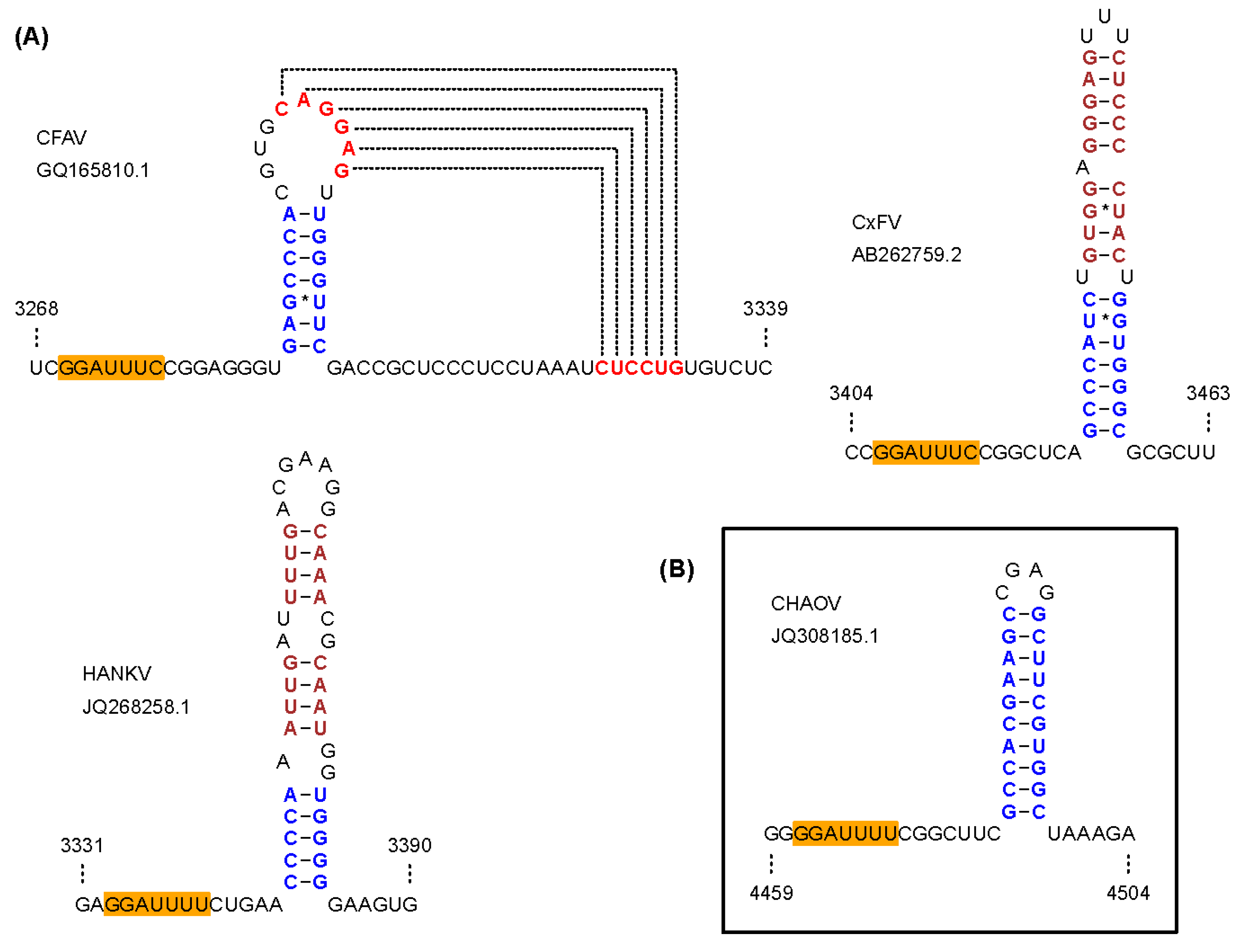
- Firth, A.E.; Brierley, I. Non-canonical translation in RNA viruses. J. Gen. Virol. 2012, 93, 1385–1409. [Google Scholar] [CrossRef] [PubMed]
- Firth, A.E.; Blitvich, B.J.; Wills, N.M.; Miller, C.L.; Atkins, J.F. Evidence for ribosomal frameshifting and a novel overlapping gene in the genomes of insect-specific flaviviruses. Virology 2010, 399, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Firth, A.E.; Atkins, J.F. A conserved predicted pseudoknot in the NS2A-encoding sequence of West Nile and Japanese encephalitis flaviviruses suggests NS1' may derive from ribosomal frameshifting. Virol. J. 2009, 6, e14. [Google Scholar] [CrossRef]
- Melian, E.B.; Hinzman, E.; Nagasaki, T.; Firth, A.E.; Wills, N.M.; Nouwens, A.S.; Blitvich, B.J.; Leung, J.; Funk, A.; Atkins, J.F.; et al. NS1' of flaviviruses in the Japanese encephalitis virus serogroup is a product of ribosomal frameshifting and plays a role in viral neuroinvasiveness. J. Virol. 2010, 84, 1641–1647. [Google Scholar] [CrossRef] [PubMed]
- Melian, E.B.; Hall-Mendelin, S.; Du, F.; Owens, N.; Bosco-Lauth, A.M.; Nagasaki, T.; Rudd, S.; Brault, A.C.; Bowen, R.A.; Hall, R.A.; et al. Programmed ribosomal frameshift alters expression of west nile virus genes and facilitates virus replication in birds and mosquitoes. PLOS Pathog. 2014, 10, e1004447. [Google Scholar] [CrossRef] [PubMed]
- Young, L.B.; Melian, E.B.; Setoh, Y.X.; Young, P.R.; Khromykh, A. Last twenty amino acids of the West Nile Virus NS1' protein are responsible for its retention in cells and formation of unique heat-stable dimers. J. Gen. Virol. 2015. [Google Scholar] [CrossRef]
- Moureau, G.; Cook, S.; Lemey, P.; Nougairede, A.; Forrester, N.L.; Khasnatinov, M.; Charrel, R.N.; Firth, A.E.; Gould, E.A.; de Lamballerie, X. New insights into flavivirus evolution, taxonomy and biogeographic history, extended by analysis of canonical and alternative coding sequences. PLOS ONE 2015, in press. [Google Scholar]
- Chambers, T.J.; Hahn, C.S.; Galler, R.; Rice, C.M. Flavivirus genome organization, expression, and replication. Ann. Rev. Microbiol. 1990, 44, 649–688. [Google Scholar] [CrossRef]
- Falgout, B.; Markoff, L. Evidence that flavivirus NS1-NS2A cleavage is mediated by a membrane-bound host protease in the endoplasmic reticulum. J. Virol. 1995, 69, 7232–7243. [Google Scholar] [PubMed]
- Choo, K.H.; Ranganathan, S. Flanking signal and mature peptide residues influence signal peptide cleavage. BMC Bioinform. 2008, 9 (Suppl 12), eS15. [Google Scholar]
- Stadler, K.; Allison, S.L.; Schalich, J.; Heinz, F.X. Proteolytic activation of tick-borne encephalitis virus by furin. J. Virol. 1997, 71, 8475–8481. [Google Scholar] [PubMed]
- Molloy, S.S.; Bresnahan, P.A.; Leppla, S.H.; Klimpel, K.R.; Thomas, G. Human furin is a calcium-dependent serine endoprotease that recognizes the sequence Arg-X-X-Arg and efficiently cleaves anthrax toxin protective antigen. J. Biol. Chem. 1992, 267, 16396–16402. [Google Scholar] [PubMed]
- Chambers, T.J.; Weir, R.C.; Grakoui, A.; McCourt, D.W.; Bazan, J.F.; Fletterick, R.J.; Rice, C.M. Evidence that the N-terminal domain of nonstructural protein NS3 from yellow fever virus is a serine protease responsible for site-specific cleavages in the viral polyprotein. Proc. Natl. Acad. Sci. USA 1990, 87, 8898–8902. [Google Scholar] [CrossRef] [PubMed]
- Chambers, T.J.; Grakoui, A.; Rice, C.M. Processing of the yellow fever virus nonstructural polyprotein: A catalytically active NS3 proteinase domain and NS2B are required for cleavages at dibasic sites. J. Virol. 1991, 65, 6042–6050. [Google Scholar] [PubMed]
- Junglen, S.; Kopp, A.; Kurth, A.; Pauli, G.; Ellerbrok, H.; Leendertz, F.H. A new flavivirus and a new vector: Characterization of a novel flavivirus isolated from uranotaenia mosquitoes from a tropical rain forest. J. Virol. 2009, 83, 4462–4468. [Google Scholar] [CrossRef] [PubMed]
- Kolodziejek, J.; Pachler, K.; Bin, H.; Mendelson, E.; Shulman, L.; Orshan, L.; Nowotny, N. Barkedji virus, a novel mosquito-borne flavivirus identified in Culex perexiguus mosquitoes, Israel, 2011. J. Gen. Virol. 2013, 94, 2449–2457. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Grubaugh, N.D.; Kondig, J.P.; Turell, M.J.; Kim, H.C.; Klein, T.A.; O'Guinn, M.L. Isolation and genomic characterization of Chaoyang virus strain ROK144 from Aedes vexans nipponii from the Republic of Korea. Virology 2013, 435, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Takhampunya, R.; Kim, H.C.; Tippayachai, B.; Lee, D.K.; Lee, W.J.; Chong, S.T.; Kim, M.S.; Lee, J.S.; Klein, T.A. Distribution and mosquito hosts of Chaoyang virus, a newly reported flavivirus from the Republic of Korea, 2008–2011. J. Med. Entomol. 2014, 51, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.S.; An, S.Y.; Wang, Y.; Han, Y.; Guo, J.Q. A new virus of Flavivirus: Chaoyang virus isolated in Liaoning province. Chin. Pub. Health 2009, 25, 769–772. (In Chinese) [Google Scholar]
- Huhtamo, E.; Cook, S.; Moureau, G.; Uzcategui, N.Y.; Sironen, T.; Kuivanen, S.; Putkuri, N.; Kurkela, S.; Harbach, R.E.; Firth, A.E.; et al. Novel flaviviruses from mosquitoes: Mosquito-specific evolutionary lineages within the phylogenetic group of mosquito-borne flaviviruses. Virology 2014, 464–465, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Huhtamo, E.; Putkuri, N.; Kurkela, S.; Manni, T.; Vaheri, A.; Vapalahti, O.; Uzcategui, N.Y. Characterization of a novel flavivirus from mosquitoes in northern Europe that is related to mosquito-borne flaviviruses of the tropics. J. Virol. 2009, 83, 9532–9540. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, F.; Cerutti, F.; Ballardini, M.; Mosca, A.; Vitale, N.; Radaelli, M.C.; Desiato, R.; Prearo, M.; Pautasso, A.; Casalone, C.; et al. Molecular characterization of flaviviruses from field-collected mosquitoes in northwestern Italy, 2011–2012. Parasit. Vectors 2014, 7, e395. [Google Scholar] [CrossRef]
- Evangelista, J.; Cruz, C.; Guevara, C.; Astete, H.; Carey, C.; Kochel, T.J.; Morrison, A.C.; Williams, M.; Halsey, E.S.; Forshey, B.M. Characterization of a novel flavivirus isolated from Culex (Melanoconion) ocossa mosquitoes from Iquitos, Peru. J. Gen. Virol. 2013, 94, 1266–1272. [Google Scholar] [CrossRef] [PubMed]
- Pauvolid-Correa, A.; Solberg, O.; Couto-Lima, D.; Kenney, J.; Serra-Freire, N.; Brault, A.; Nogueira, R.; Langevin, S.; Komar, N. Nhumirim virus, a novel flavivirus isolated from mosquitoes from the Pantanal, Brazil. Arch. Virol. 2015, 160, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Ochieng, C.; Lutomiah, J.; Makio, A.; Koka, H.; Chepkorir, E.; Yalwala, S.; Mutisya, J.; Musila, L.; Khamadi, S.; Richardson, J.; et al. Mosquito-borne arbovirus surveillance at selected sites in diverse ecological zones of Kenya; 2007–2012. Virol. J. 2013, 10, e140. [Google Scholar] [CrossRef]
- Liu, H.; Gao, X.; Liang, G. Newly recognized mosquito-associated viruses in mainland China, in the last two decades. Virol. J. 2011, 8, e68. [Google Scholar] [CrossRef]
- Charlier, N.; Molenkamp, R.; Leyssen, P.; Paeshuyse, J.; Drosten, C.; Panning, M.; de Clercq, E.; Bredenbeek, P.J.; Neyts, J. Exchanging the yellow fever virus envelope proteins with Modoc virus prM and E proteins results in a chimeric virus that is neuroinvasive in SCID mice. J. Virol. 2004, 78, 7418–7426. [Google Scholar] [CrossRef] [PubMed]
- Charlier, N.; Davidson, A.; Dallmeier, K.; Molenkamp, R.; de Clercq, E.; Neyts, J. Replication of not-known-vector flaviviruses in mosquito cells is restricted by intracellular host factors rather than by the viral envelope proteins. J. Gen. Virol. 2010, 91, 1693–1697. [Google Scholar] [CrossRef] [PubMed]
- Tumban, E.; Maes, N.E.; Schirtzinger, E.E.; Young, K.I.; Hanson, C.T.; Whitehead, S.S.; Hanley, K.A. Replacement of conserved or variable sequences of the mosquito-borne dengue virus 3' UTR with homologous sequences from Modoc virus does not change infectivity for mosquitoes. J. Gen. Virol. 2013, 94, 783–788. [Google Scholar] [CrossRef] [PubMed]
- Saiyasombat, R.; Carrillo-Tripp, J.; Miller, W.A.; Bredenbeek, P.J.; Blitvich, B.J. Substitution of the premembrane and envelope protein genes of Modoc virus with the homologous sequences of West Nile virus generates a chimeric virus that replicates in vertebrate but not mosquito cells. Virol. J. 2014, 11, e150. [Google Scholar] [CrossRef]
- Isawa, H.; Kuwata, R.; Tajima, S.; Hoshino, K.; Sasaki, T.; Takasaki, T.; Kobayashi, M.; Sawabe, K. Construction of an infectious cDNA clone of Culex flavivirus, an insect-specific flavivirus from Culex mosquitoes. Arch. Virol. 2012, 157, 975–979. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blitvich, B.J.; Firth, A.E. Insect-Specific Flaviviruses: A Systematic Review of Their Discovery, Host Range, Mode of Transmission, Superinfection Exclusion Potential and Genomic Organization. Viruses 2015, 7, 1927-1959. https://doi.org/10.3390/v7041927
Blitvich BJ, Firth AE. Insect-Specific Flaviviruses: A Systematic Review of Their Discovery, Host Range, Mode of Transmission, Superinfection Exclusion Potential and Genomic Organization. Viruses. 2015; 7(4):1927-1959. https://doi.org/10.3390/v7041927
Chicago/Turabian StyleBlitvich, Bradley J., and Andrew E. Firth. 2015. "Insect-Specific Flaviviruses: A Systematic Review of Their Discovery, Host Range, Mode of Transmission, Superinfection Exclusion Potential and Genomic Organization" Viruses 7, no. 4: 1927-1959. https://doi.org/10.3390/v7041927
APA StyleBlitvich, B. J., & Firth, A. E. (2015). Insect-Specific Flaviviruses: A Systematic Review of Their Discovery, Host Range, Mode of Transmission, Superinfection Exclusion Potential and Genomic Organization. Viruses, 7(4), 1927-1959. https://doi.org/10.3390/v7041927