
A Taxonomic Review of Clostridium difficile Phages and Proposal of a Novel Genus, “Phimmp04likevirus”

Abstract
:1. Introduction

{kind=link}
{kind=link}
| Morphotype | Sequenced phages of this morphology | Capsid diameter (nm) | Tail length (nm) | Genome size (kbp) |
|---|---|---|---|---|
| MM | ΦCD119, φC2, phiCDHM1, phiCDHM19, | 60–70 | 110–130 | 53–57 |
| SMV | ΦMMP04, phiCDHM11, phiCDHM13, phiCDHM14 | 40–58 | 106 | ~31 |
| LTM | ϕCD27, ΦMMP02 | 60–70 | 150–260 | ~51 |
2. Results
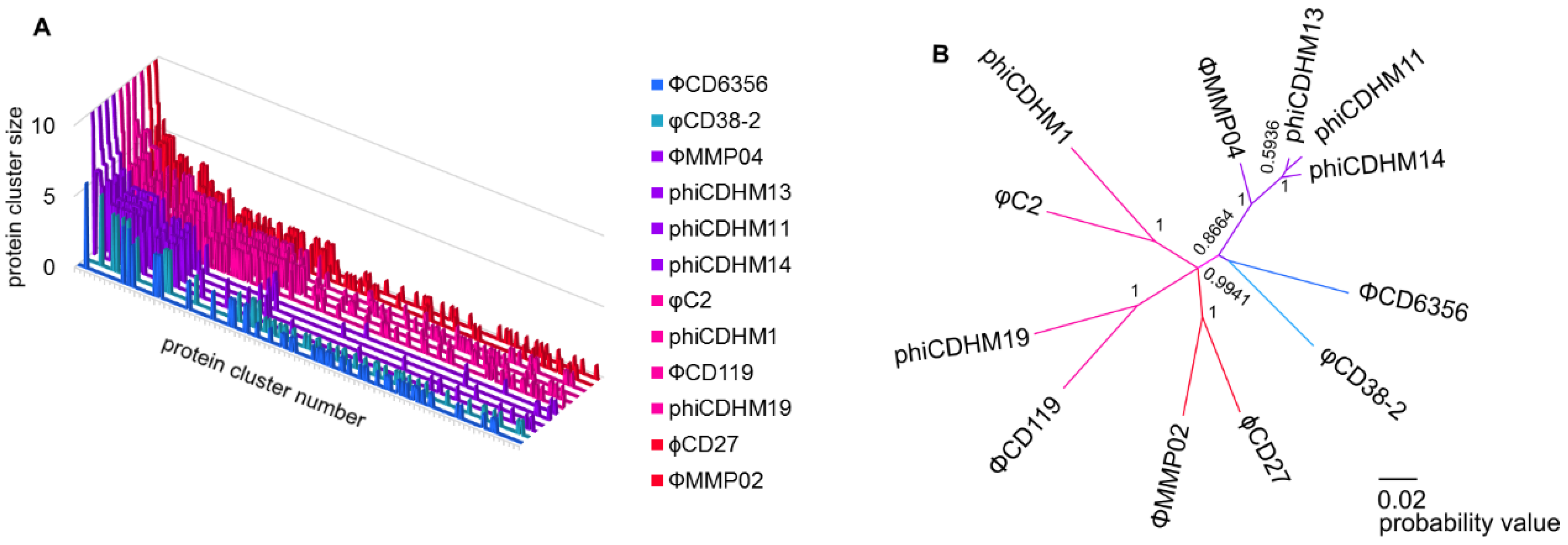
Taxonomic Clusters and Protein Cluster Analysis


3. Discussion
3.1. PhiCD119likevirus Clade
3.2. PhiMMP04likevirus Clade
3.3. Characteristic Genome Features of the New Genus
3.4. C. Difficile Phages Outside of the Established and Proposed Genera
3.5. Phage and Prophage Sequences in NCBI
4. Materials and Methods
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Govind, R.; Fralick, J.; Rolfe, R. Genomic organization and molecular characterization of Clostridium difficile bacteriophage phi cd119. J. Bacteriol. 2006, 188, 2568–2577. [Google Scholar] [CrossRef] [PubMed]
- Goh, S.; Ong, P.; Song, K.; Riley, T.; Chang, B. The complete genome sequence of Clostridium difficile phage phiC2 and comparisons to phiCD119 and inducible prophages of CD630. Microbiology 2007, 153, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.; Narbad, A.; Gasson, M. Molecular characterization of a Clostridium difficile bacteriophage and its cloned biologically active endolysin. J. Bacteriol. 2008, 190, 6734–6740. [Google Scholar] [CrossRef] [PubMed]
- Horgan, M.; O’Sullivan, O.; Coffey, A.; Fitzgerald, G.; van Sinderen, D.; McAuliffe, O.; Ross, R. Genome analysis of the Clostridium difficile phage phiCD6356, a temperate phage of the siphoviridae family. Gene 2010, 462, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Sekulovic, O.; Meessen-Pinard, M.; Fortier, L. Prophage-stimulated toxin production in Clostridium difficile NAP1/027 lysogens. J. Bacteriol. 2011, 193, 2726–2734. [Google Scholar] [CrossRef] [PubMed]
- Meessen-Pinard, M.; Sekulovic, O.; Fortier, L. Evidence of in vivo prophage induction during Clostridium difficile infection. Appl. Environ. Microbiol. 2012, 78, 7662–7670. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, K.R.; Kropinski, A.M.; Clokie, M.R.J. What does the talking? Quorum sensing signalling genes discovered in a bacteriophage genome. PLoS ONE 2014, 9, e85131. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, K.R.; Flores, C.O.; Lawley, T.D.; Clokie, M.R.J. Abundant and diverse clustered regularly interspaced short palindromic repeat spacers in Clostridium difficile strains and prophages target multiple phage types within this pathogen. MBio 2014, 5, e01045–e01113. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, K.R.; Clokie, M.R.J. Clostridium difficile phages: Still difficult? Front. Microbiol. 2014, 5, e184. [Google Scholar] [CrossRef]
- Lavigne, R.; Darius, P.; Summer, E.J.; Seto, D.; Mahadevan, P.; Nilsson, A.S.; Ackermann, H.W.; Kropinski, A.M. Classification of myoviridae bacteriophages using protein sequence similarity. BMC Microbiol. 2009, 9, e224. [Google Scholar] [CrossRef]
- Goh, S.; Chang, B.; Riley, T. Effect of phage infection on toxin production by Clostridium difficile. J. Med. Microbiol. 2005, 54, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Meader, E.; Mayer, M.; Gasson, M.; Steverding, D.; Carding, S.; Narbad, A. Bacteriophage treatment significantly reduces viable Clostridium difficile and prevents toxin production in an in vitro model system. Anaerobe 2010, 16, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Goh, S.; Hussain, H.; Chang, B.J.; Emmett, W.; Riley, T.V.; Mullany, P. Phage C2 mediates transduction of tn6215, encoding erythromycin resistance, between Clostridium difficile strains. MBio 2013, 4, e00840-13. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.; Garefalaki, V.; Spoerl, R.; Narbad, A.; Meijers, R. Structure-based modification of a Clostridium difficile-targeting endolysin affects activity and host range. J. Bacteriol. 2011, 193, 5477–5486. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating the human-ape split by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Sekulovic, O.; Garneau, J.R.; Néron, A.; Fortier, L.-C. Characterization of temperate phages infecting Clostridium difficile isolates from human and animal origin. Appl. Environ. Microbiol 2014, 80, 2555–2563. [Google Scholar] [CrossRef] [PubMed]
- Bignell, C.; Thomas, C.M. The bacterial para-parb partitioning proteins. J. Biotechnol. 2001, 91, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, K.; Parkhill, J.; Crook, J.; Horsnell, T.; Rice, P.; Rajandream, M.A.; Barrell, B. Artemis: Sequence visualization and annotation. Bioinformatics 2000, 16, 944–945. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. Cd-hit: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef] [PubMed]
- FigTree v.1.4.2. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 17 May 2015).
- Inkscape v0.91. Available online: https://inkscape.org/en/download/ (accessed on 17 May 2015).
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. Mega6: Molecular evolutionary genetics analysis version 6.0. J. Mol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hargreaves, K.R.; Clokie, M.R.J. A Taxonomic Review of Clostridium difficile Phages and Proposal of a Novel Genus, “Phimmp04likevirus”. Viruses 2015, 7, 2534-2541. https://doi.org/10.3390/v7052534
Hargreaves KR, Clokie MRJ. A Taxonomic Review of Clostridium difficile Phages and Proposal of a Novel Genus, “Phimmp04likevirus”. Viruses. 2015; 7(5):2534-2541. https://doi.org/10.3390/v7052534
Chicago/Turabian StyleHargreaves, Katherine R., and Martha R. J. Clokie. 2015. "A Taxonomic Review of Clostridium difficile Phages and Proposal of a Novel Genus, “Phimmp04likevirus”" Viruses 7, no. 5: 2534-2541. https://doi.org/10.3390/v7052534
APA StyleHargreaves, K. R., & Clokie, M. R. J. (2015). A Taxonomic Review of Clostridium difficile Phages and Proposal of a Novel Genus, “Phimmp04likevirus”. Viruses, 7(5), 2534-2541. https://doi.org/10.3390/v7052534