
BnSGS3 Has Differential Effects on the Accumulation of CMV, ORMV and TuMV in Oilseed Rape


Abstract
:1. Introduction
2. Results
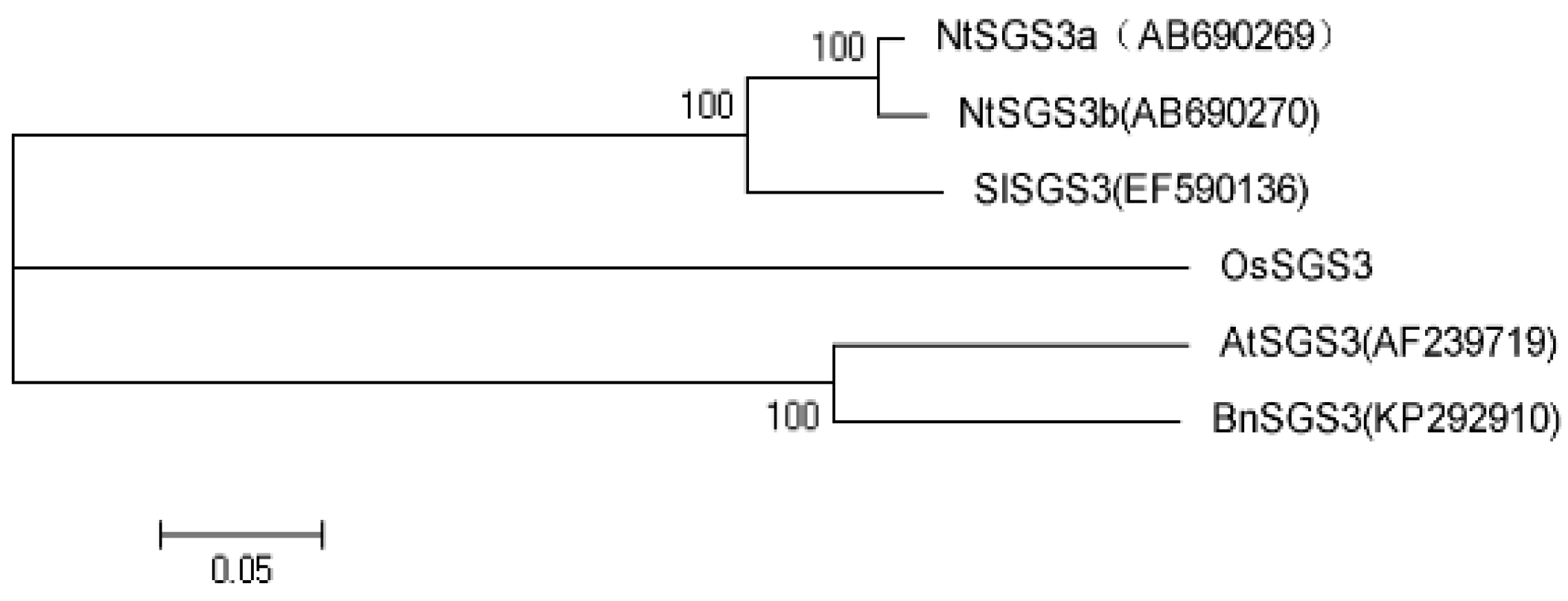
2.1. Sequence Comparison and Phylogenetic Analysis of SGS3 Genes in Different Hosts

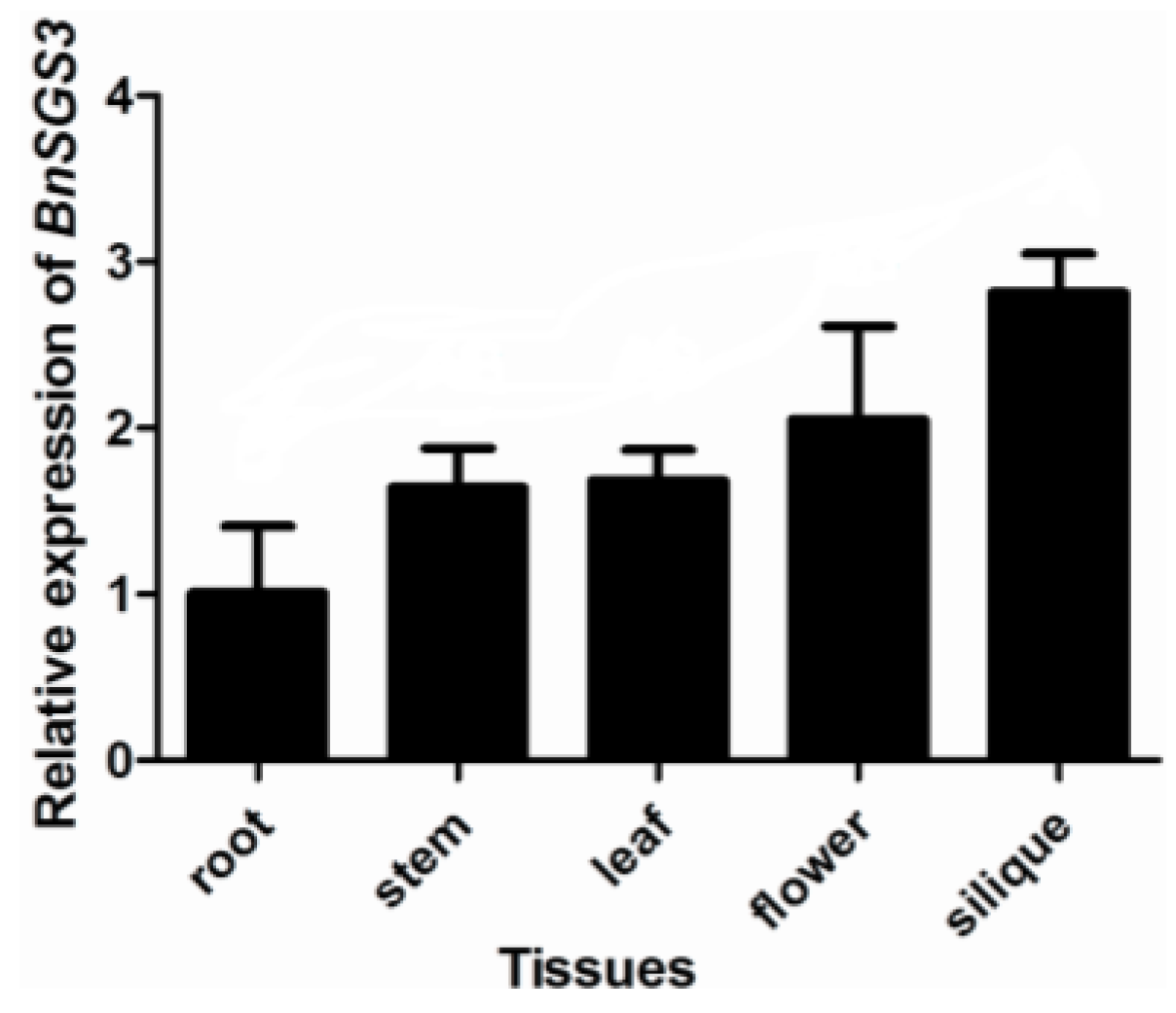
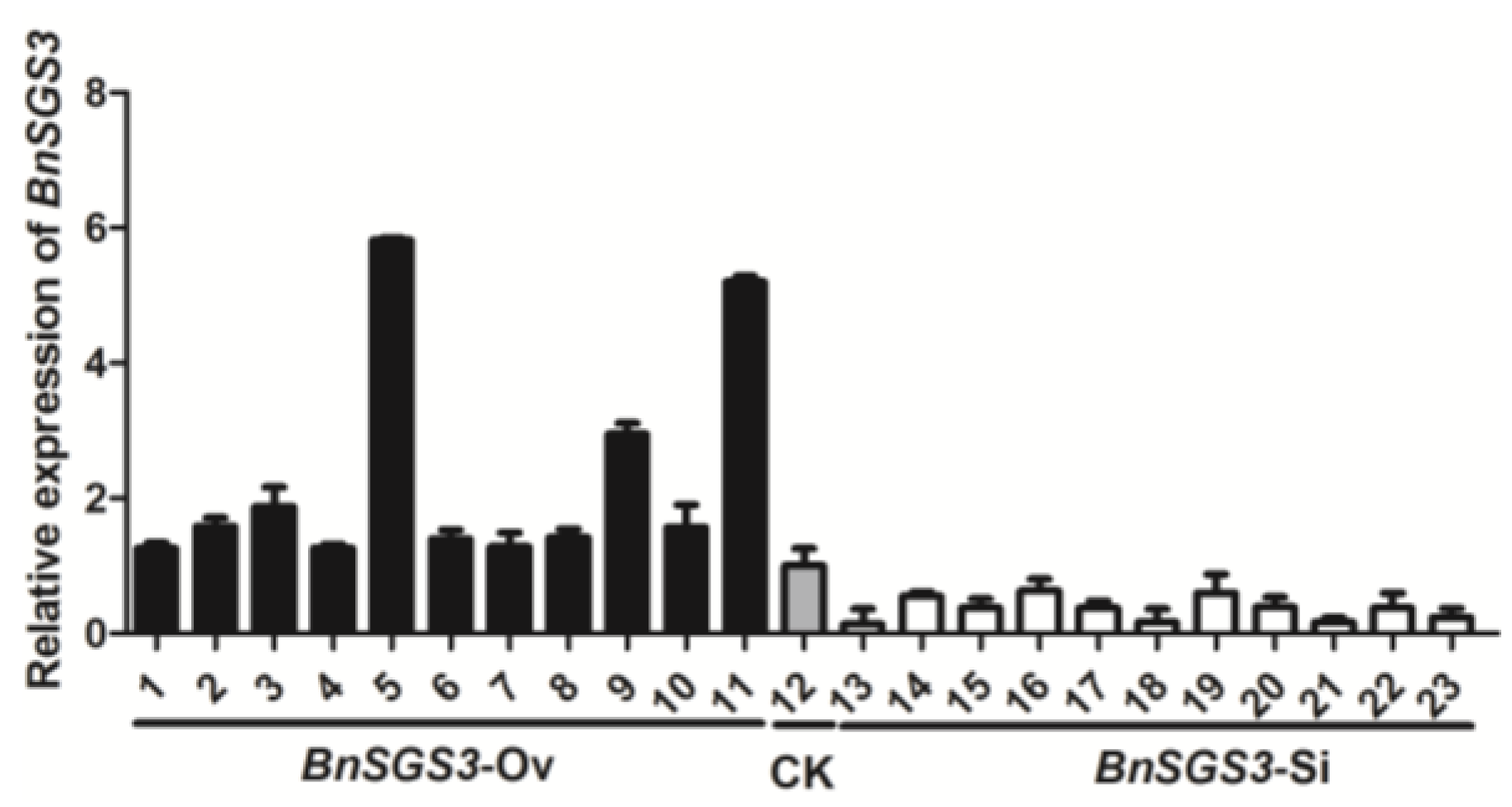
2.2. BnSGS3 Expression in Transgenic and Non-Transgenic Plants of Oilseed Rape


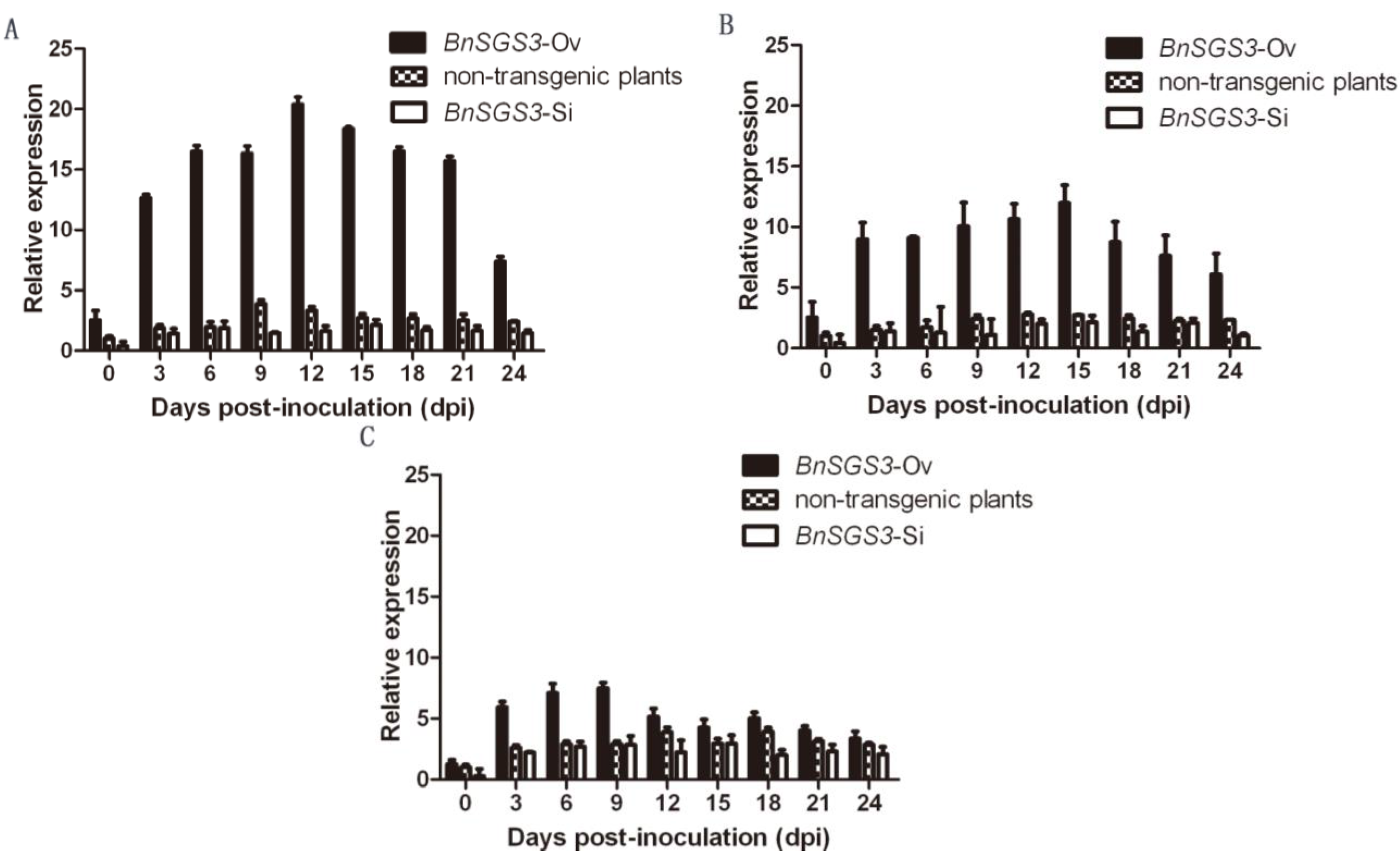
2.3. Dynamics of BnSGS3 Expression in T0 Generation Transgenic and Non-Transgenic Plants of Oilseed Rape after Viral Infection

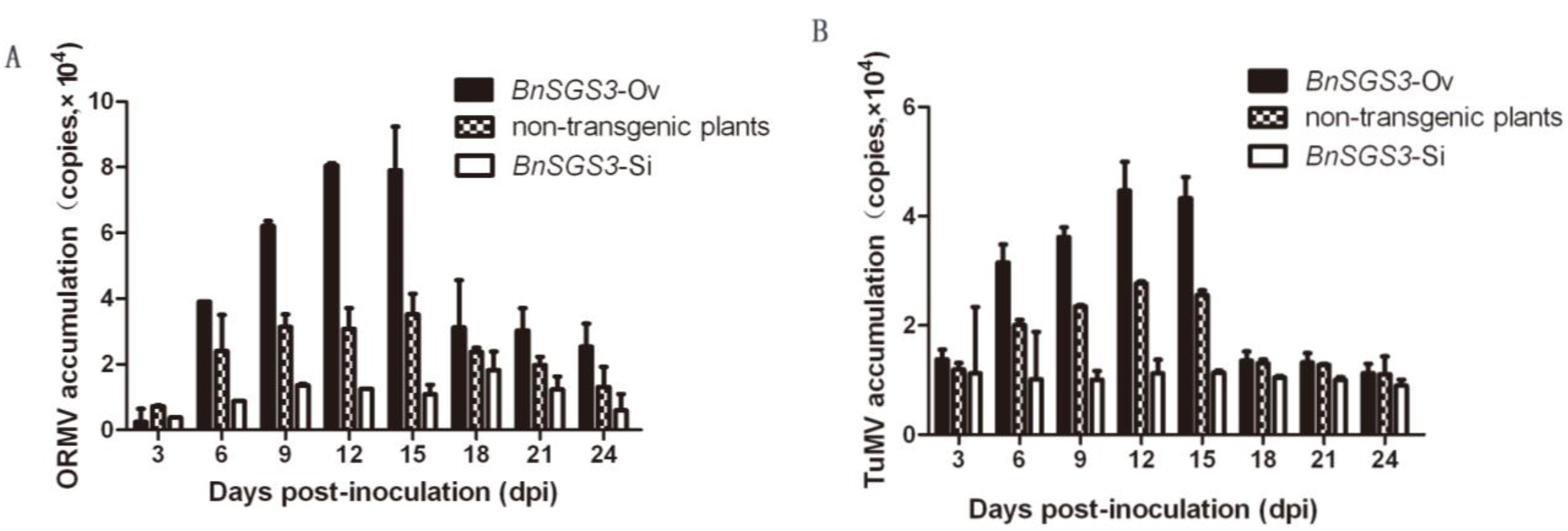
2.4. Effect of BnSGS3 on the Accumulation of ORMV, TuMV and CMV


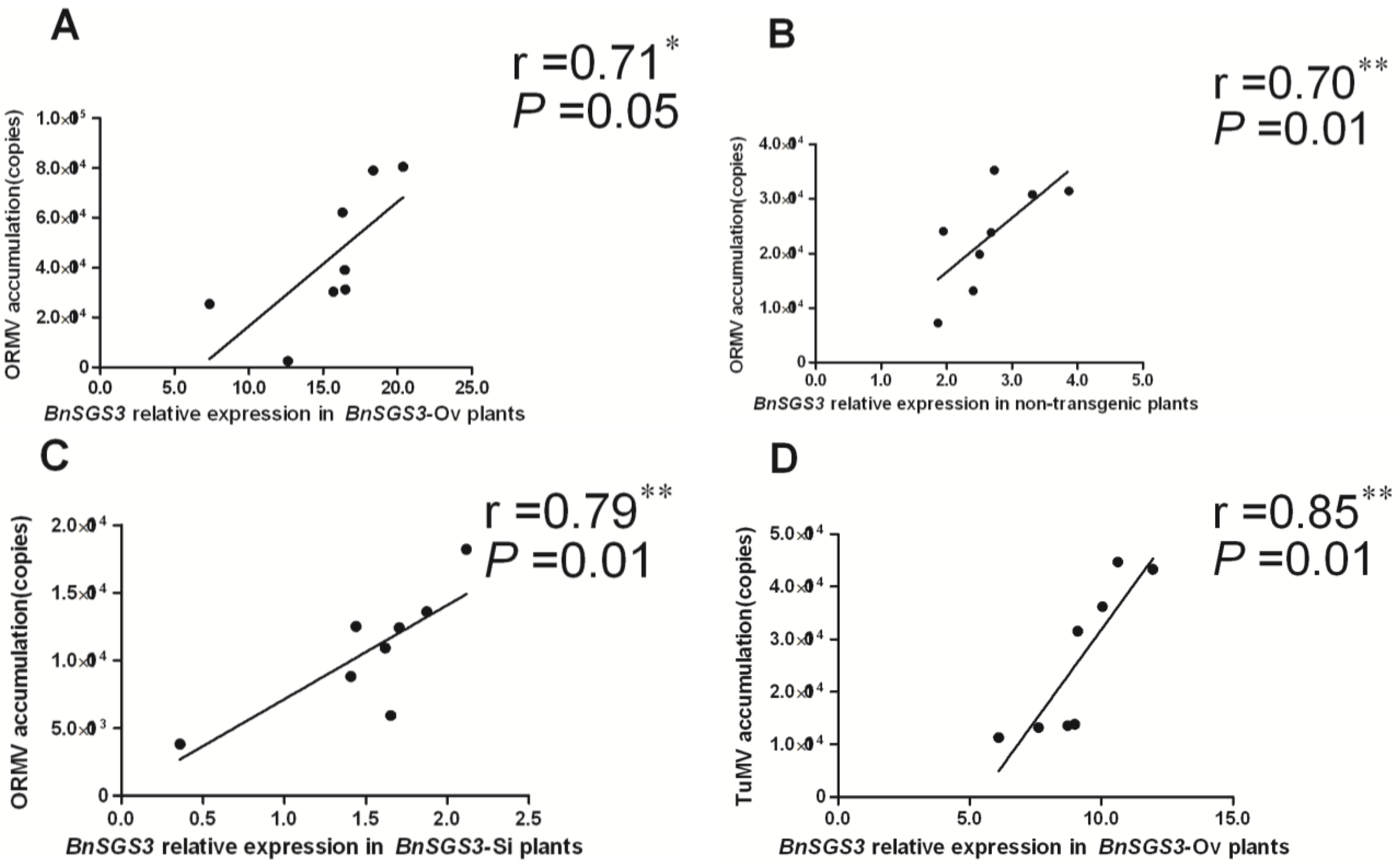
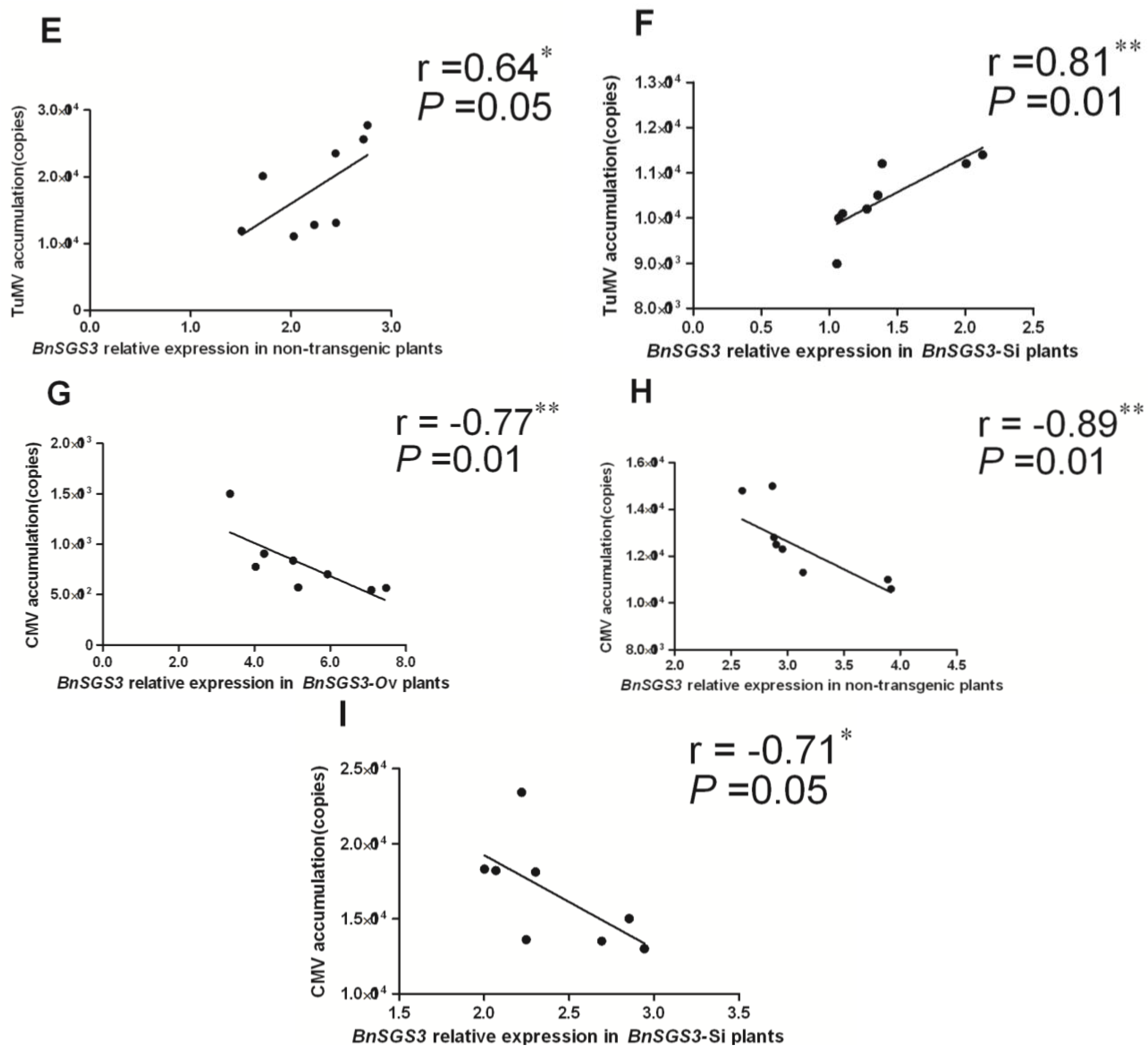
2.5. Correlation between BnSGS3 Expression and Virus Accumulation


3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Sequence Analysis of BnSGS3 and Vector Construction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) | Tm (°C) | Product (bp) |
|---|---|---|---|
| BnSGS3 cDNA amplification | |||
| Bn-F1 | TGAGGTTCTGGACAGGGATC | 55 | 1974 |
| Bn-R1 | GCCGTCTTACTGAAAATGGA | ||
| Vector construction | |||
| Bn-F2 | GTACTCGAGGTTTGCTCTCTGTTTGGTT | 55 | 1848 |
| Bn-R2 | CGTCTCGAGGCTAGTAGTCTTCTGTGTC | ||
| BnRI-C | TATACTCGAGCAGTGGAAGGGTTTGGGTG | 54 | 404 |
| BnRI-KC | GCCCGAATTCTCCCTACTTGACAGTGTTG | ||
| BnRI-A | CGAATCTAGACAGTGGAAGGGTTTGGGTG | 54 | 404 |
| BnRI-E | GCCGAAGCTTTCCCTACTTGACAGTGTTG | ||
| Detection of expression quantity | |||
| Bn-F | TGGAAGGGTTTGGGTGAGGAG | 62 | 181 |
| Bn-R | GTGGACCATAGGAGTGGCGTG | ||
| Actin-F | CTGGAATTGCTGACCGTATGAG | 62 | 145 |
| Actin-R | ATCTGTTGGAAAGTGCTGAGGG | ||
| Bar-F | TGCCAGAAACCCACGTCAT | 55 | 485 |
| Bar-R | CTGCACCATCGTCAACCACTA | ||
| Detection of virus accumulation | |||
| CMV-F | CCTCACCGGTACTGGTTTATC | 62 | 107 |
| CMV-R | CTTTCGCATGTCGCCAATATC | ||
| TuMV-F | GGAAGTAAACGCTGGAACCT | 62 | 96 |
| TuMV-R | GCCACTCTTTGCTCGTATCT | ||
| ORMV-F | CTGTGGCCATTAGGAGTCAA | 62 | 108 |
| ORMV-R | GCGCAGTAGTCCAAGGTAATA | ||
4.3. Production of BnSGS3-Ov and BnSGS3-Si Brassica napus Transformants
4.4. Virus Purification and Inoculation
4.5. Sample Collection and RNA Extraction
4.6. Quantitative RT-PCR
4.7. Correlation Analysis of Virus Accumulation vs. BnSGS3 Expression
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflict of Interests
References
- Music, M.S.; Nguyen, H.D.; Cerni, S.; Mamula, E.; Ohshima, K.; Skoric, D. Multilocus sequence analysis of “Candidatus Phytoplasma asteris” strain and the genome analysis of turnip mosaic virus co-infecting oilseed rape. J. Appl. Microbiol. 2014, 117, 774–785. [Google Scholar]
- Cai, L.; Chen, K.R.; Zhang, X.J.; Yan, L.Y.; Hou, M.S.; Xu, Z.Y. Biological and molecular characterization of a crucifer tobamovirus infecting oilseed rape. Biochem. Genet. 2009, 47, 451–461. [Google Scholar]
- Vaucheret, H.; Béclin, C.; Fagard, M. Post-transcriptional gene silencing in plants. J. Cell Sci. 2001, 114, 3083–3091. [Google Scholar]
- Mourrain, P.; Béclin, C.; Elmayan, T.; Feuerbach, F.; Godon, C.; Morel, J.B.; Jouette, D.; Lacombe, A.M.; Nikic, S.; Picault, N.; et al. Arabidopsis SGS2 and SGS3 genes are required for posttranscriptional gene silencing and natural virus resistance. Cell 2000, 101, 533–542. [Google Scholar]
- Pumplin, N.; Voinnet, O. RNA silencing suppression by plant pathogens: Defence, counter-defence and counter-counter-defence. Nat. Rev. Microbiol. 2013, 11, 745–760. [Google Scholar]
- Masson, I.L.; Jauvion, V.; Bouteiller, N.; Rivard, M.; Elmayan, T.; Vaucheret, H. Mutations in the Arabidopsis H3K4me 2/3 demethylase JMJ14 suppress posttranscriptional gene silencing by decreasing transgene transcription. Plant Cell 2012, 24, 3603–3612. [Google Scholar]
- Butaye, K.M.; Goderis, I.J.; Wouters, P.F.; Pues, J.M.; Delaure, S.L.; Broekaert, W.F.; Depicker, A.; Cammue, B.P.; de Bolle, M.F. Stable high-level transgene expression in Arabidopsis thaliana using gene silencing mutants and matrix attachment regions. Plant J. 2004, 39, 440–449. [Google Scholar]
- Zheng, Z.M.; Xing, Y.; He, X.J.; Li, W.B.; Hu, Y.L.; Yadav, S.K.; Oh, J.E.; Zhu, J.K. An SGS3-like protein functions in RNA-directed DNA methylation and transcriptional gene silencing in Arabidopsis. Plant J. 2010, 62, 92–99. [Google Scholar] [PubMed]
- Vance, V.; Vaucheret, H. RNA silencing in plants-defense and counterdefense. Science 2001, 292, 2277–2280. [Google Scholar] [PubMed]
- Morel, J.B.; Godon, C.; Mourrain, P.; Béclin, C.; Boutet, S.; Feuerbach, F.; Proux, F.; Vaucheret, H. Fertile hypomorphic ARGONAUTE (ago1) mutants impaired in post-transcriptional gene silencing and virus resistance. Plant Cell 2002, 14, 629–639. [Google Scholar] [PubMed]
- Moreno, A.B.; Alba, A.E.M.; Bardou, F.; Crespi, M.D.; Vaucheret, H.; Maizel, A.; Mallory, A.C. Cytoplasmic and nuclear quality control and turnover of single-stranded RNA modulate post-transcriptional gene silencing in plants. Nucleic Acids Res. 2013, 10, 1–10. [Google Scholar]
- Maine, E.M. A conserved mechanism for post-transcriptional gene silencing? Genome Biol. 2000, 3, 1–4. [Google Scholar]
- Rajamäki, M.L.; Streng, J.; Valkonen, J.P.T. Silencing suppressor protein VPg of a potyvirus interacts with the plant silencing-related protein SGS3. Mol. Plant Microbe. Interact. 2014, 27, 1199–1210. [Google Scholar] [PubMed]
- Chen, H.; Zhang, L.R.; Yu, K.F.; Wang, A. Pathogenesis of Soybean mosaic virus in soybean carrying Rsv1 gene is associated with miRNA and siRNA pathways, and breakdown of AGO1 homeostasis. Virology 2015, 476, 395–404. [Google Scholar] [PubMed]
- Fukunaga, R.; Doudna, J.A. dsRNA with 5′overhangs contrebutes to endogenous and antiviral RNA silencing pathways in plants. EMBO J. 2009, 28, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Kumakura, N.; Takeda, A.; Fujioka, Y.; Motose, H.; Takano, R.; Watanabe, Y. SGS3 and RDR6 interact and colocalize in cytoplasmic SGS3/RDR6-bodies. FEBS Lett. 2009, 583, 1261–1266. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.; Voinnet, O.; Chappell, L.; Baulcombe, D. Two classes of short interfering RNA in RNA silencing. EMBO J. 2002, 21, 4671–4679. [Google Scholar] [CrossRef] [PubMed]
- Boutet, S.; Vazquez, F.; Liu, J.; Béclin, C.; Fagard, M.; Gratias, A.; Morel, J.B.; Crete, P.; Chen, X.M.; Vaucheret, H. Arabidopsis HEN1: A genetic link between endogenous miRNA controlling development and siRNA controlling transgene silencing and virus resistance. Curr. Biol. 2003, 13, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Yang, Z.Y.; Li, J.J.; Minakhina, S.; Yang, M.C.; Padgett, R.W.; Steward, R.; Chen, X.M. Methylation as a crucial step in plant microRNA biogenesis. Science 2005, 307, 932–935. [Google Scholar] [CrossRef] [PubMed]
- Béclin, C.; Waterhouse, P.; Boutet, S.; Vaucheret, H. A branched pathway for transgene-induced RNA silencing in plants. Curr. Biol. 2002, 12, 684–688. [Google Scholar] [CrossRef] [PubMed]
- Muangsan, N.; Béclin, C.; Vaucheret, H.; Robertson, D. Geminivirus VIGs of endogenous genes requires SGS2/SDE1 pathway for silencing in plants. Plant J. 2004, 38, 1004–1014. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Trudeau, V.L. The XS domain of a plant specific SGS3 protein adopts a unique RNA recognition motif (RRM) fold. Cell Cycle 2008, 7, 2268–2270. [Google Scholar] [CrossRef] [PubMed]
- Bateman, A. The SGS3 protein involved in PTGS finds a family. BMC Bioinform. 2002, 3, e21. [Google Scholar] [CrossRef]
- Elmayan, T.; Adenot, X.; Gissot, L.; Lauressergues, D.; Gy, I.; Vaucheret, H. A neomorphic sgs3 allele stabilizing miRNA cleavage products reveals that SGS3 acts as a homodimer. FEBS J. 2009, 276, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Glick, E.; Zrachya, A.; Levy, Y.; Mett, A.; Gidoni, D.; Belausov, E.; Citovsky, V.; Gafni, Y. Interaction with host SGS3 is required for suppression of RNA silencing by tomato yellow leaf curl virus V2 protein. Proc. Natl. Acad. Sci. USA 2007, 105, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.G.; Xiao, D.L.; Wu, J.G.; Jia, D.S.; Yuan, Z.J.; Liu, Y.; Hu, L.Y.; Han, Z.; Wei, T.Y.; Lin, Q.Y.; et al. p2 of Rice stripe virus (RSV) interacts with OsSGS3 and is a silencing suppressor. Mol. Plant Pathol. 2011, 12, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Iki, T.; Tsutsui, Y.; Miyashita, K.; Poethig, R.S.; Habu, Y.; Ishikawa, M. 3′ fragment of miR173-programmed RISC-cleaved RNA is protected from degradation in a complex with RISC and SGS3. Proc. Natl. Acad. Sci. USA 2013, 110, 4117–4122. [Google Scholar] [CrossRef] [PubMed]
- Dalmay, T.; Hamilton, A.; Rudd, S.; Angell, S.; Baulcombe, D.C. An RNA-dependent RNA polymerase gene in Arabidopsis is required for posttranscriptional gene silencing mediated by a transgene but not by a virus. Cell 2000, 101, 543–553. [Google Scholar] [CrossRef] [PubMed]
- Anandalakshmi, R.; Pruss, G.J.; Ge, X.; Marathe, R.; Mallory, A.C.; Smith, T.H.; Vance, V.B. A viral suppressor of gene silencing in plants. Proc. Natl. Acad. Sci. USA 1998, 95, 13079–13084. [Google Scholar] [CrossRef] [PubMed]
- Béclin, C.; Berthome, R.; Palauqui, J.C.; Tepfer, M.; Vaucheret, H. Infection of tobacco or Arabidopsis plants by CMV counteracts systemic post-transcriptional silencing of nonviral (trans)genes. Virology 1998, 252, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Brigneti, G.; Voinnet, O.; Li, W.X.; Ji, L.H.; Ding, S.W.; Baulcombe, D.C. Viral pathogenicity determinants are suppressors of transgene silencing in Nicotiana benthamiana. EMBO J. 1998, 17, 6739–6746. [Google Scholar] [CrossRef] [PubMed]
- Kasschau, K.D.; Carrington, J.C. A counterdefensive strategy of plant viruses: Suppression of posttranscriptional gene silencing. Cell 1998, 95, 461–470. [Google Scholar] [CrossRef]
- Voinnet, O.; Pinto, Y.M.; Baulcombe, D.C. Suppression of gene silencing: A general strategy used by diverse DNA and RNA viruses of plants. Proc. Natl. Acad. Sci. USA 1999, 96, 14147–14152. [Google Scholar] [CrossRef] [PubMed]
- Roth, B.M.; Pruss, G.J.; Vance, V.B. Plant viral suppressors of RNA silencing. Virus Res. 2004, 102, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Qu, F.; Morris, T.J. Suppressors of RNA silencing encoded by plant viruses and their role in viral infections. FEBS Lett. 2005, 579, 5958–5964. [Google Scholar] [CrossRef] [PubMed]
- Mallory, A.C.; Ely, L.; Smith, T.H.; Marathe, R.; Anandalakshmi, R.; Fagard, M.; Vaucheret, H.; Pruss, G.; Bowman, L.; Vancea, V.B. HC-Pro suppression of transgene silencing eliminates the small RNAs but not transgene methylation or the mobile signal. Plant Cell 2011, 13, 571–583. [Google Scholar] [CrossRef]
- Kubota, K.; Tsuda, S.; Tamai, A.; Meshi, T. Tomato mosaic virus replication protein suppresses virus-targeted posttranscriptional gene silencing. J. Virol. 2003, 77, 11016–11026. [Google Scholar] [CrossRef] [PubMed]
- Morel, J.B.; Mourrain, P.; Béclin, C.; Vaucheret, H. DNA methylation and chromatin structure affect transcriptional and post-transcriptional transgene silencing in Arabidopsis. Curr. Biol. 2000, 10, 1591–1594. [Google Scholar] [PubMed]
- Voinnet, O. Use, tolerance and avoidance of amplified RNA silencing by plants. Trends Plant Sci. 2008, 13, 1360–1385. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.B.; Jovel, J.; Udomporn, P.; Wang, Y.; Wu, Q.F.; Li, W.X.; Gasciolli, V.; Vaucheret, H.; Ding, S.W. The 21-nucleotide, but not 22-nucleotide, viral secondary small interfering RNAs direct potent antiviral defense by two cooperative argonautes in Arabidopsis thaliana. Plant Cell 2011, 23, 1625–1638. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.S.; Ding, S.W. A viral protein inhibits the long range signaling activity of the gene silencing signal. EMBO J. 2002, 21, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Liao, Q.; Zhu, L.; Du, Z.; Zeng, R.; Feng, J.; Chen, J. Satellite RNA-mediated reduction of cucumber mosaic virus genomic RNAs accumulation in Nicotiana tabacum. Acta Biochim. Biophys. Sin. 2007, 39, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.R.; Yuan, Y.R.; Pei, Y.; Lin, S.S.; Tuschl, T.; Patel, D.J.; Chua, N.H. Cucumber mosaic virus-encoded 2b suppressor inhibits Arabidopsis Argonaute1 cleavage activity to counter plant defense. Gene Dev. 2006, 20, 3255–3268. [Google Scholar] [CrossRef] [PubMed]
- González, I.; Martínez, L.; Rakitina, D.V.; Lewsey, M.G.; Atencio, F.A.; Llave, C.; Kalinina, N.O.; Carr, J.P.; Palukaitis, P.; Canto, T. Cucumber mosaic virus 2b protein subcellular targets and interactions: Their significance to RNA silencing suppressor activity. Mol. Plant-Microbe Interact. 2010, 23, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Vain, P.; Afolabi, A.S.; Worland, B.; Snape, J.W. Transgene behaviour in populations of rice plants transformed using a new dual binary vector system: PGreen/pSoup. Theor. Appl. Genet. 2003, 107, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Hellens, R.P.; Edwards, E.A.; Leyland, N.R.; Bean, S.; Mullineaux, P.M. pGreen: A versatile and flexible binary Ti vector for Agrobacterium-mediated plant transformation. Plant Mol. Biol. 2000, 42, 819–832. [Google Scholar]
- Liu, F.; Xiong, X.J.; Wu, L.; Fu, D.H.; Hayward, A.; Zeng, X.H.; Cao, Y.L.; Wu, Y.H.; Li, Y.J.; Wu, G. BraLTP1, a lipid transfer protein gene involved in epicuticular wax deposition, cell proliferation and flower development in Brassica napus. PLoS ONE 2014, 9, e110272. [Google Scholar] [CrossRef] [PubMed]
- Jenner, C.E.; Walsh, J.A. Pathotypic variation in turnip mosaic virus with special reference to European isolates. Plant Pathol. 1996, 45, 848–856. [Google Scholar] [CrossRef]
- Aguilar, I.; Sánchez, F.; Martín, A.M.; Martínez-Herrera, D.; Ponz, F. Nucleotide sequence of Chinese rape mosaic virus (oilseed rape mosaic virus), a crucifer tobamovirus infectious on Arabidopsis thaliana. Plant Mol. Biol. 1996, 30, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Chen, C.W.; Doddapaneni, H.; Duan, Y.P.; Civerolo, E.L.; Bai, X.J.; Zhao, X.L. A new diagnostic system for ultra-sensitive and specific detection and quantification of Candidatus Liberibacter asiaticus, the bacterium associated with citrus Huanglongbing. J. Microbiol. Methods 2010, 81, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Mason, G.; Provero, P.; Vaira, A.M.; Accotto, G.P. Estimating the number of integrations in transformed plants by quantitative real-time PCR. BMC Biotechnol. 2002, 2, e20. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Q.; Wang, J.; Hou, M.; Liu, S.; Huang, J.; Cai, L. BnSGS3 Has Differential Effects on the Accumulation of CMV, ORMV and TuMV in Oilseed Rape. Viruses 2015, 7, 4169-4185. https://doi.org/10.3390/v7082815
Chen Q, Wang J, Hou M, Liu S, Huang J, Cai L. BnSGS3 Has Differential Effects on the Accumulation of CMV, ORMV and TuMV in Oilseed Rape. Viruses. 2015; 7(8):4169-4185. https://doi.org/10.3390/v7082815
Chicago/Turabian StyleChen, Quan, Jie Wang, Mingsheng Hou, Shengyi Liu, Junyan Huang, and Li Cai. 2015. "BnSGS3 Has Differential Effects on the Accumulation of CMV, ORMV and TuMV in Oilseed Rape" Viruses 7, no. 8: 4169-4185. https://doi.org/10.3390/v7082815
APA StyleChen, Q., Wang, J., Hou, M., Liu, S., Huang, J., & Cai, L. (2015). BnSGS3 Has Differential Effects on the Accumulation of CMV, ORMV and TuMV in Oilseed Rape. Viruses, 7(8), 4169-4185. https://doi.org/10.3390/v7082815