
Characterization of Humoral Responses Induced by an H7N9 Influenza Virus-Like Particle Vaccine in BALB/C Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
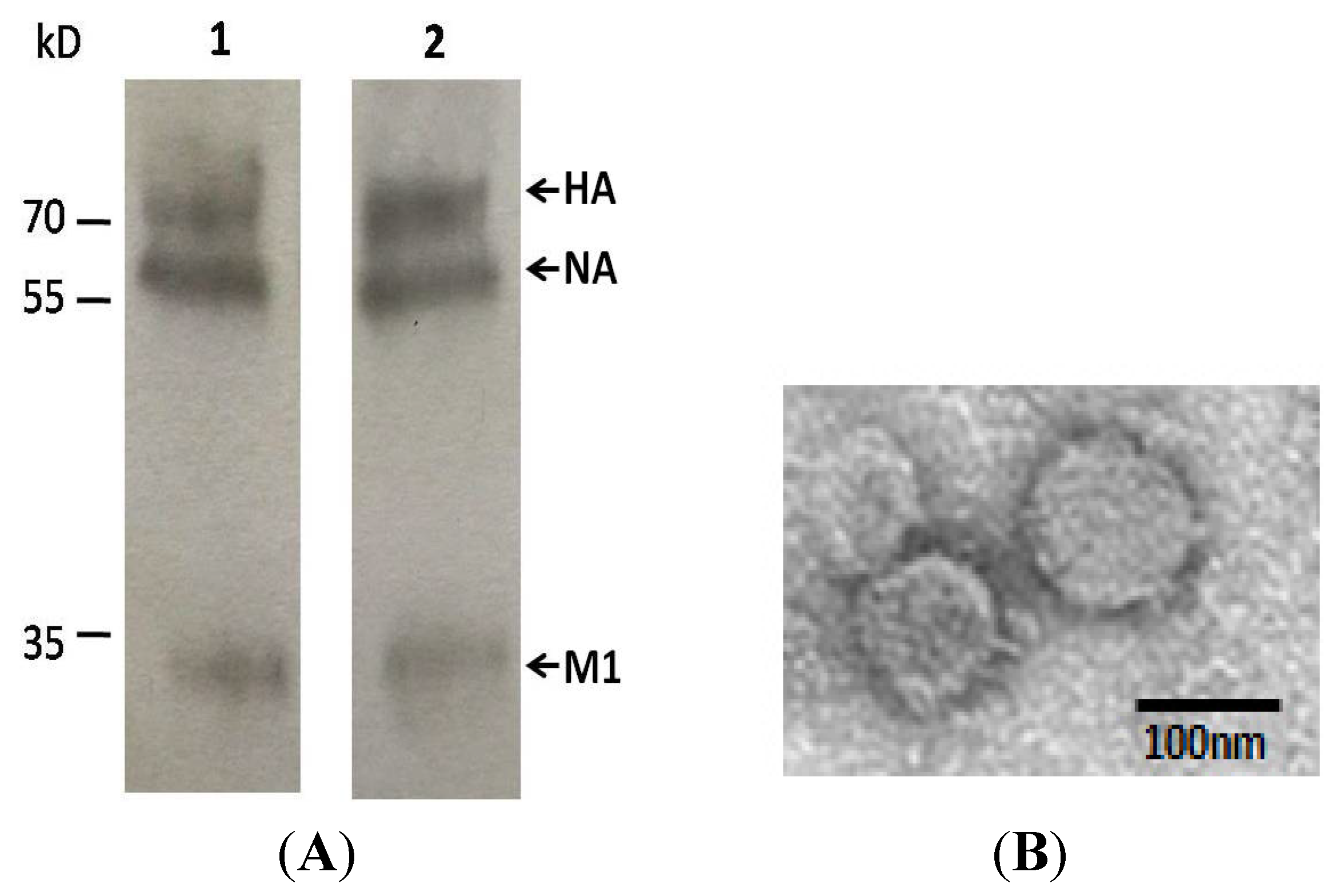
2.1. Production and Characterization of VLPs

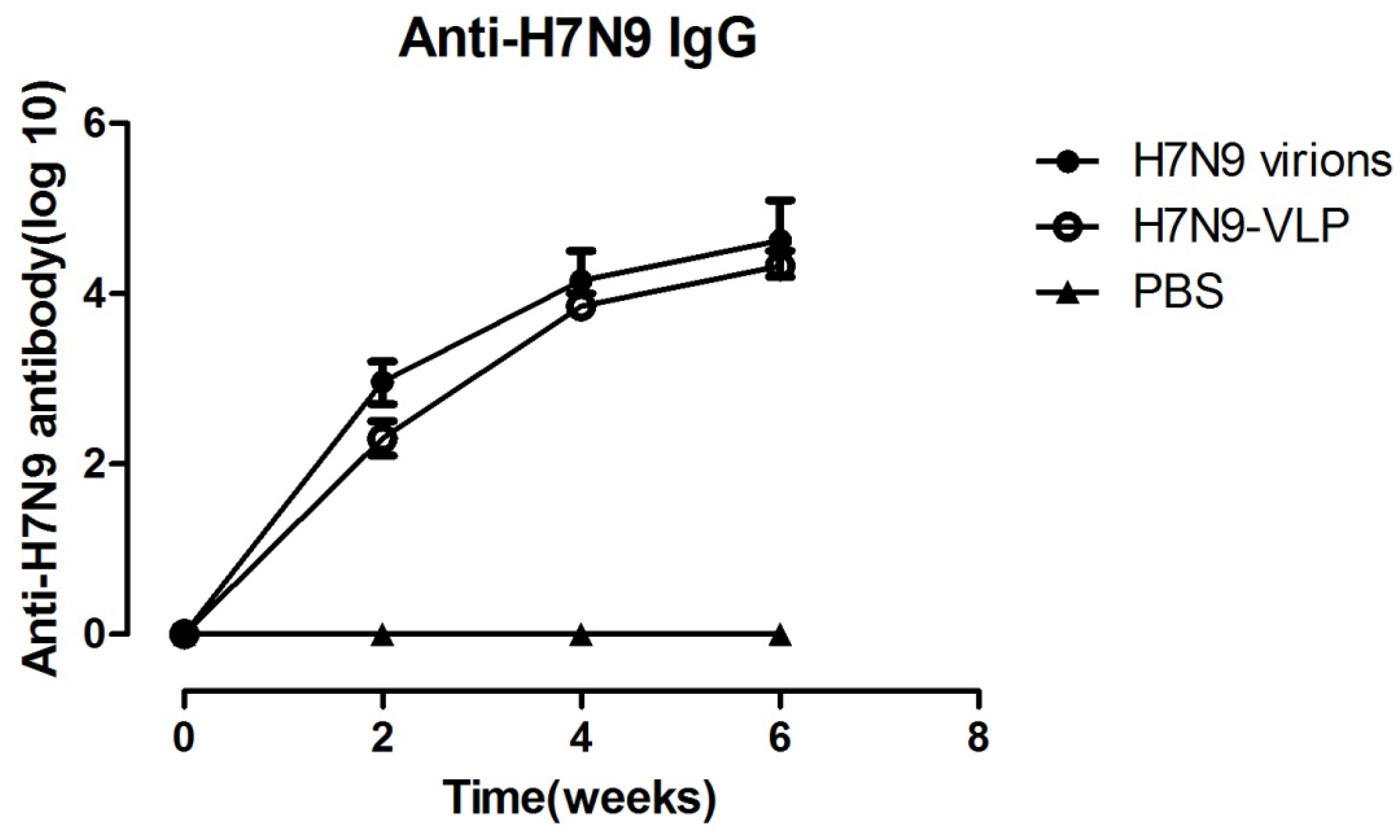
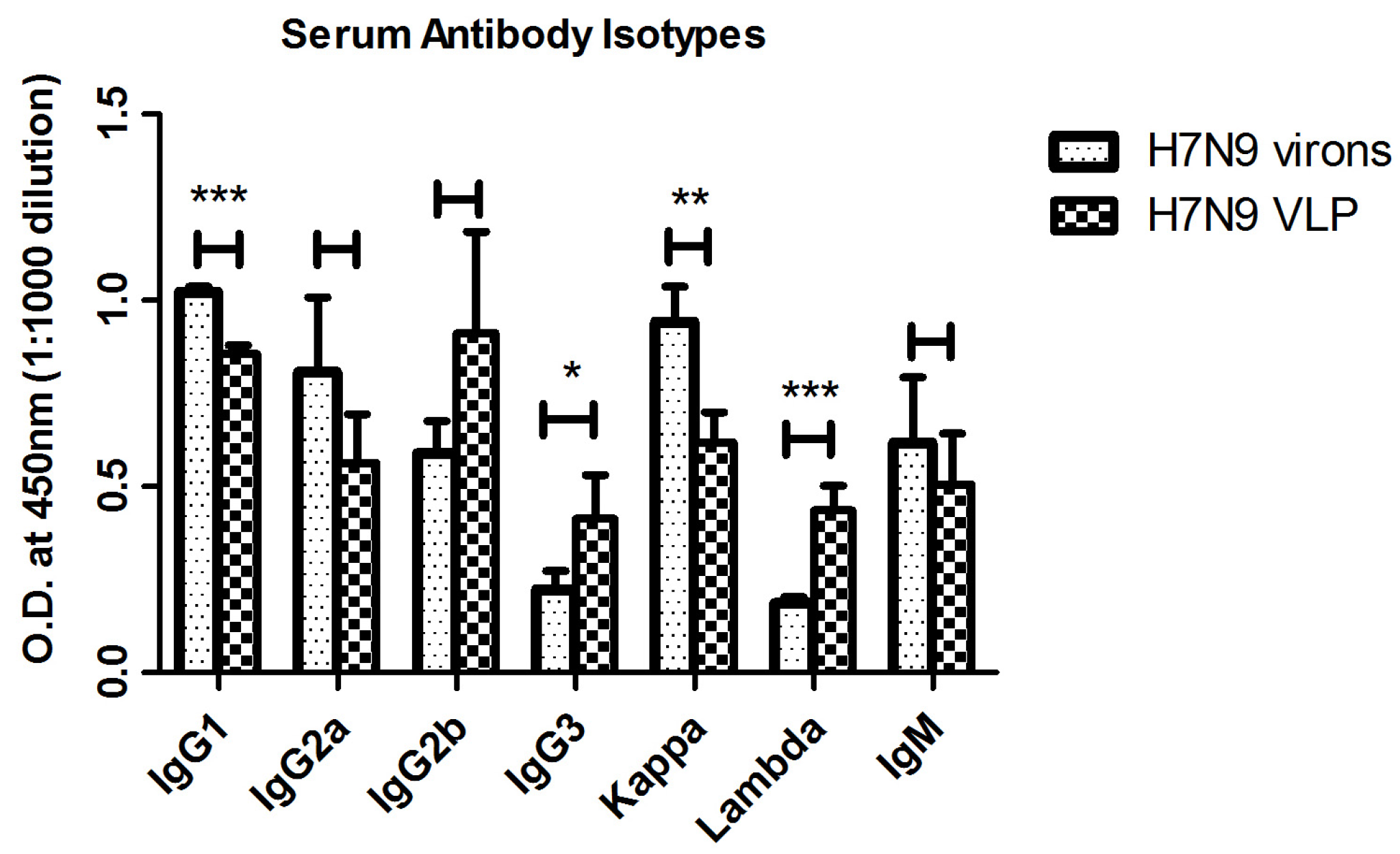
2.2. Antibody Responses Induced by Immunizations


2.3. H7N9 VLPs Induced Virus-Specific T Cell Responses


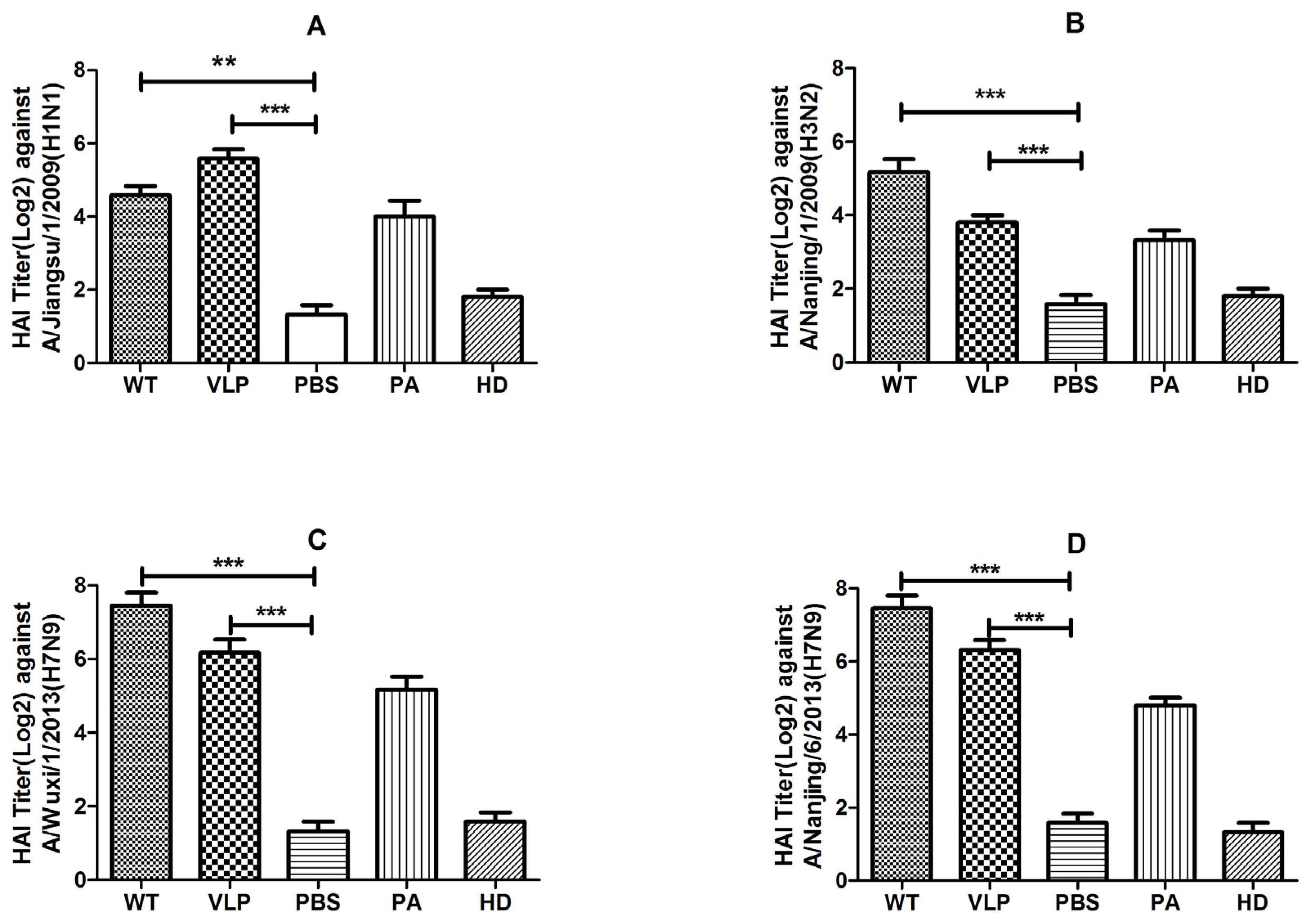
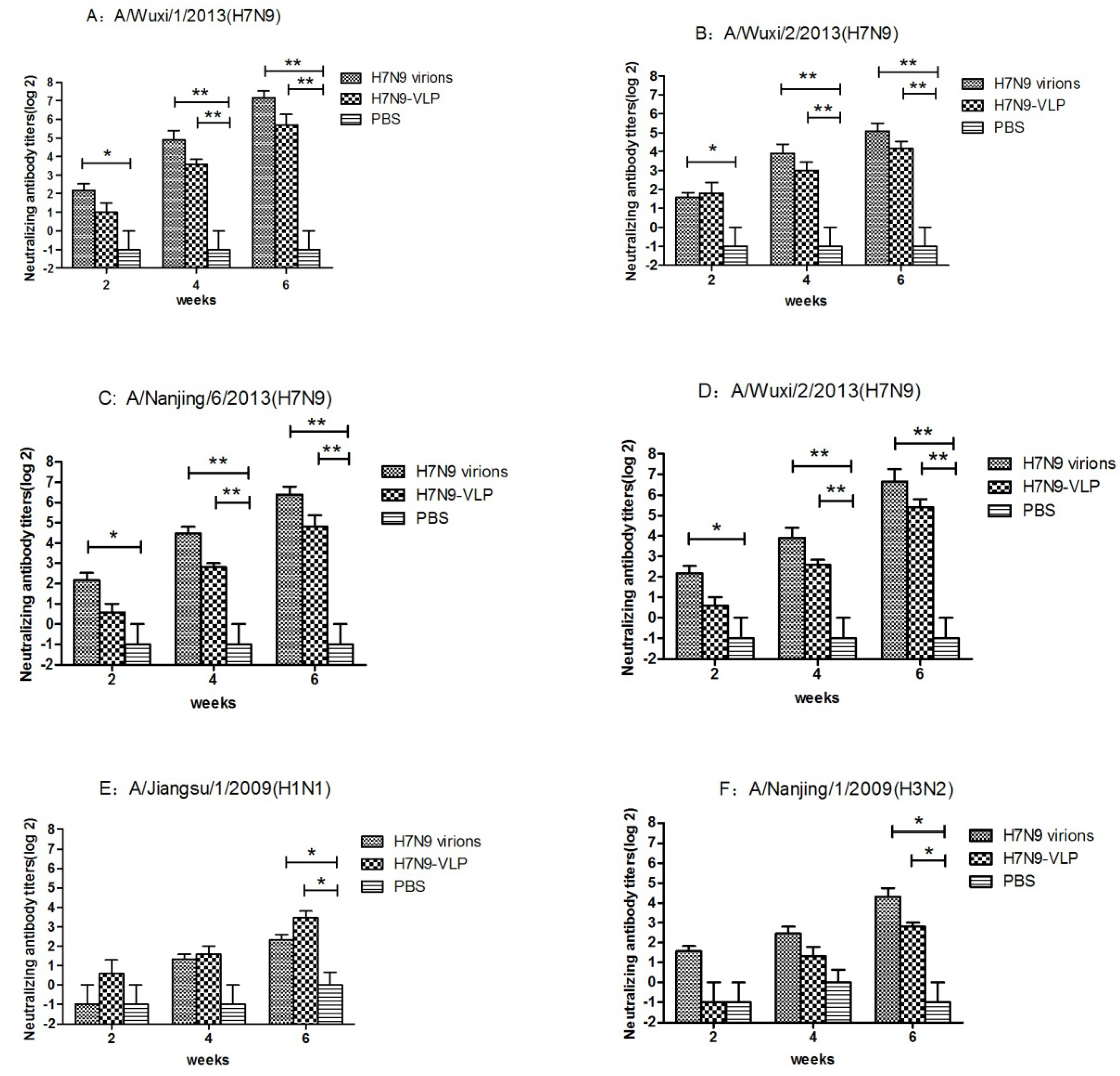
2.4. VLPs Induced Hemagglutination–Inhibition and Neutralization Activity

3. Discussion
4. Materials and Methods
4.1. Cells and Virus
4.2. Construction of H7N9 VLP Expression Plasmids
4.3. Transient Transfection of 293T Cells with H7N9 VLP Expression Plasmids
4.4. Western Blot Analysis
4.5. Electron Microscopy
4.6. Purification of H7N9-Derived VLPs
4.7. Mice Immunization
4.8. ELISA to Measure Antibody Titer and Antibody Isotypes in Serum
4.9. Hemagglutination Inhibition (HAI) Titer and Neutralization Assay
4.10. Enzyme Linked Immunospot (ELISPOT) Assay
4.11. Statistical Analysis
5. Ethics Statement
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dai, J.; Zhou, X.; Dong, D.; Liu, Y.; Gu, Q.; Zhu, B.; Wu, C.; Cai, H. Human infection with a novel avian-origin influenza A (H7N9) virus: Serial chest radiographic and ct findings. Chin. Med. J. 2014, 127, 2206–2211. [Google Scholar] [PubMed]
- World Health Organization. Human infection with avian influenza A (H7N9) virus in China. World Health Organization 2013 April. Available online: http://www.who.int/csr/don/2013_04_18/en/ (accessed on 18 April 2013).
- Gao, R.; Cao, B.; Hu, Y.; Feng, Z.; Wang, D.; Hu, W.; Chen, J.; Jie, Z.; Qiu, H.; Xu, K.; et al. Human infection with a novel avian-origin influenza A (H7N9) virus. N. Engl. J. Med. 2013, 368, 1888–1897. [Google Scholar] [CrossRef] [PubMed]
- Wikipedia. Influenza A virus subtype H7N9. Wikipedia 2014 July. Available online: http://en.wikipedia.org/wiki/Influenza_A_virus_subtype_H7N9 (accessed on 15 July 2014).
- Liao, Q.; Ip, D.K.; Tsang, T.K.; Cao, B.; Jiang, H.; Liu, F.; Zheng, J.; Peng, Z.; Wu, P.; Huai, Y.; et al. A clinical prediction rule for diagnosing human infections with avian influenza A (H7N9) in a hospital emergency department setting. BMC Med. 2014, 12, e127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Zhou, L.; Zhou, M.; Chen, Z.; Li, F.; Wu, H.; Xiang, N.; Chen, E.; Tang, F.; Wang, D.; et al. Epidemiology of human infections with avian influenza A (H7N9) virus in china. N. Engl. J. Med. 2014, 370, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Roos, R. Experts offer dim view of potential vaccine response to H7N9. Regents of the University of Minnesota 2013 May. Available online: http://www.cidrap.umn.edu/news-perspective/2013/05/experts-offer-dim-view-potential-vaccine-response-h7n9 (accessed on 5 May 2013).
- Kalthoff, D.; Bogs, J.; Grund, C.; Tauscher, K.; Teifke, J.P.; Starick, E.; Harder, T.; Beer, M. Avian influenza h7n9/13 and h7n7/13: A comparative virulence study in chickens, pigeons, and ferrets. J. Virol. 2014, 88, 9153–9165. [Google Scholar] [CrossRef] [PubMed]
- De Groot, A.S.; Ardito, M.; Terry, F.; Levitz, L.; Ross, T.; Moise, L.; Martin, W. Low immunogenicity predicted for emerging avian-origin H7N9: Implication for influenza vaccine design. Hum. Vaccines Immunother. 2013, 9, 950–956. [Google Scholar]
- Behzadian, F.; Goodarzi, Z.; Fotouhi, F.; Saberfar, E. Baculoviral co-expression of HA, NA and M1 proteins of highly pathogenic H5N1 influenza virus in insect cells. Jundishapur J. Microbiol. 2013, 6, e7665. [Google Scholar] [CrossRef]
- He, F.; Madhan, S.; Kwang, J. Baculovirus vector as a delivery vehicle for influenza vaccines. Expert Rev. Vaccines 2009, 8, 455–467. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Macias, C. Virus-like particle (vlp)-based vaccines for pandemic influenza: Performance of a vlp vaccine during the 2009 influenza pandemic. Hum.Vaccines Immunother. 2012, 8, 411–414. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Bae, S.W.; Park, J.K.; Kwon, J.H.; Yuk, S.S.; Song, J.M.; Kang, S.M.; Kwon, Y.K.; Kim, H.Y.; Song, C.S. Virus-like particle vaccine protects against H3N2 canine influenza virus in dog. Vaccine 2013, 31, 3268–3273. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.C.; Lin, Y.F.; Chong, P.; Wu, S.C. Broader neutralizing antibodies against H5N1 viruses using prime-boost immunization of hyperglycosylated hemagglutinin DNA and virus-like particles. PLoS ONE 2012, 7, e39075. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.C.; Liu, W.C.; Lin, Y.F.; Huang, Y.H.; Liu, J.H.; Wu, S.C. Heterologous prime-boost immunization regimens using adenovirus vector and virus-like particles induce broadly neutralizing antibodies against H5N1 avian influenza viruses. Biotechnol. J. 2013, 8, 1315–1322. [Google Scholar] [CrossRef] [PubMed]
- Park, J.K.; Lee, D.H.; Youn, H.N.; Kim, M.S.; Lee, Y.N.; Yuk, S.S.; Lim, T.H.; Jang, J.H.; Kwon, J.H.; Kim, B.Y.; et al. Protective efficacy of crude virus-like particle vaccine against hpai H5N1 in chickens and its application on diva strategy. Influenza Other Respir. Viruses 2013, 7, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Park, J.K.; Lee, Y.N.; Song, J.M.; Kang, S.M.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Song, C.S. H9N2 avian influenza virus-like particle vaccine provides protective immunity and a strategy for the differentiation of infected from vaccinated animals. Vaccine 2011, 29, 4003–4007. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.E.; Flyer, D.C.; Raghunandan, R.; Liu, Y.; Wei, Z.; Wu, Y.; Kpamegan, E.; Courbron, D.; Fries, L.F., 3rd; Glenn, G.M. Development of influenza H7N9 virus like particle (vlp) vaccine: Homologous a/anhui/1/2013 (h7n9) protection and heterologous a/chicken/jalisco/cpa1/2012 (H7N3) cross-protection in vaccinated mice challenged with H7N9 virus. Vaccine 2013, 31, 4305–4313. [Google Scholar] [CrossRef] [PubMed]
- Klausberger, M.; Wilde, M.; Palmberger, D.; Hai, R.; Albrecht, R.A.; Margine, I.; Hirsh, A.; Garcia-Sastre, A.; Grabherr, R.; Krammer, F. One-shot vaccination with an insect cell-derived low-dose influenza a h7 virus-like particle preparation protects mice against H7N9 challenge. Vaccine 2014, 32, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Bellone, S.; El-Sahwi, K.; Cocco, E.; Casagrande, F.; Cargnelutti, M.; Palmieri, M.; Bignotti, E.; Romani, C.; Silasi, D.A.; Azodi, M.; et al. Human papillomavirus type 16 (HPV-16) virus-like particle l1-specific cd8+ cytotoxic T lymphocytes (ctls) are equally effective as e7-specific cd8+ ctls in killing autologous HPV-16-positive tumor cells in cervical cancer patients: Implications for l1 dendritic cell-based therapeutic vaccines. J. Virol. 2009, 83, 6779–6789. [Google Scholar] [PubMed]
- Huret, C.; Desjardins, D.; Miyalou, M.; Levacher, B.; Amadoudji Zin, M.; Bonduelle, O.; Combadiere, B.; Dalba, C.; Klatzmann, D.; Bellier, B. Recombinant retrovirus-derived virus-like particle-based vaccines induce hepatitis C virus-specific cellular and neutralizing immune responses in mice. Vaccine 2013, 31, 1540–1547. [Google Scholar] [CrossRef] [PubMed]
- Quan, F.S.; Kim, Y.C.; Song, J.M.; Hwang, H.S.; Compans, R.W.; Prausnitz, M.R.; Kang, S.M. Long-term protective immunity from an influenza virus-like particle vaccine administered with a microneedle patch. Clin. Vaccine Immunol. 2013, 20, 1433–1439. [Google Scholar] [CrossRef] [PubMed]
- Sistigu, A.; Bracci, L.; Valentini, M.; Proietti, E.; Bona, R.; Negri, D.R.; Ciccaglione, A.R.; Tritarelli, E.; Nisini, R.; Equestre, M.; et al. Strong cd8+ T cell antigenicity and immunogenicity of large foreign proteins incorporated in HIV-1 vlps able to induce a nef-dependent activation/maturation of dendritic cells. Vaccine 2011, 29, 3465–3475. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Guo, H.C.; Wei, Y.Q.; Dong, H.; Han, S.C.; Ao, D.; Sun, D.H.; Wang, H.M.; Cao, S.Z.; Sun, S.Q. Self-assembly of virus-like particles of canine parvovirus capsid protein expressed from Escherichia coli and application as virus-like particle vaccine. Appl. Microbiol. Biotechnol. 2014, 98, 3529–3538. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Limas, W.A.; Pastor, A.R.; Esquivel-Soto, E.; Esquivel-Guadarrama, F.; Ramirez, O.T.; Palomares, L.A. Immunogenicity and protective efficacy of yeast extracts containing rotavirus-like particles: A potential veterinary vaccine. Vaccine 2014, 32, 2794–2798. [Google Scholar] [CrossRef] [PubMed]
- Cervera, L.; Gutierrez-Granados, S.; Martinez, M.; Blanco, J.; Godia, F.; Segura, M.M. Generation of HIV-1 gag vlps by transient transfection of hek 293 suspension cell cultures using an optimized animal-derived component free medium. J. Biotechnol. 2013, 166, 152–165. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Peng, H.; Tao, Q.; Zhao, X.; Tang, H.; Tang, Z.; Wang, Y.; Wang, Y.; Zhao, P.; Qi, Z. Serologic assay for avian-origin influenza A (H7N9) virus in adults of Shanghai, Guangzhou and Yunnan, china. J. Clin. Virol.: Off. Publ. Pan Am. Soc. Clin. Virol. 2014, 60, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Steinhauer, D.A. Role of hemagglutinin cleavage for the pathogenicity of influenza virus. Virology 1999, 258, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Air, G.M. Sequence relationships among the hemagglutinin genes of 12 subtypes of influenza A virus. Proc. Natl. Acad. Sci. USA 1981, 78, 7639–7643. [Google Scholar] [CrossRef] [PubMed]
- Bright, R.A.; Carter, D.M.; Daniluk, S.; Toapanta, F.R.; Ahmad, A.; Gavrilov, V.; Massare, M.; Pushko, P.; Mytle, N.; Rowe, T.; et al. Influenza virus-like particles elicit broader immune responses than whole virion inactivated influenza virus or recombinant hemagglutinin. Vaccine 2007, 25, 3871–3878. [Google Scholar] [CrossRef] [PubMed]
- Haredy, A.M.; Yamada, H.; Sakoda, Y.; Okamatsu, M.; Yamamoto, N.; Omasa, T.; Mori, Y.; Kida, H.; Okamoto, S.; Okuno, Y.; et al. Neuraminidase gene homology contributes to the protective activity of influenza vaccines prepared from the influenza virus library. J. Gen. Virol. 2014, 95, 2365–2371. [Google Scholar]
- Kovacsovics-Bankowski, M.; Rock, K.L. Presentation of exogenous antigens by macrophages: Analysis of major histocompatibility complex class i and ii presentation and regulation by cytokines. Eur. J. Immunol. 1994, 24, 2421–2428. [Google Scholar] [PubMed]
- Garcia-Sastre, A. Inhibition of interferon-mediated antiviral responses by influenza a viruses and other negative-strand RNA viruses. Virology 2001, 279, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Buonaguro, L.; Tornesello, M.L.; Tagliamonte, M.; Gallo, R.C.; Wang, L.X.; Kamin-Lewis, R.; Abdelwahab, S.; Lewis, G.K.; Buonaguro, F.M. Baculovirus-derived human immunodeficiency virus type 1 virus-like particles activate dendritic cells and induce ex vivo T-cell responses. J. Virol. 2006, 80, 9134–9143. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Avalos, R.T.; Ponimaskin, E.; Nayak, D.P. Influenza virus assembly: Effect of influenza virus glycoproteins on the membrane association of M1 protein. J. Virol. 2000, 74, 8709–8719. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.Y.; Chang, C.Y.; Ma, H.H.; Wang, C.W.; Chen, Y.T.; Hsiao, P.W.; Chang, C.C.; Chan, C.H.; Liu, C.C.; Chen, J.R. Squalene-adjuvanted H7N9 virus vaccine induces robust humoral immune response against H7N9 and H7N7 viruses. Vaccine 2014, 32, 4485–4494. [Google Scholar] [CrossRef] [PubMed]
- Quan, F.S.; Compans, R.W.; Cho, Y.K.; Kang, S.M. Ginseng and salviae herbs play a role as immune activators and modulate immune responses during influenza virus infection. Vaccine 2007, 25, 272–282. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Serological detection of avian influenza A (H7N9) virus infections by turkey haemagglutination-inhibition assay. World Health Organization 2013 May. Available online: http://www.who.int/influenza/gisrs_laboratory/cnic_serological_diagnosis_hai_a_h7n9.pdf (accessed on 15 May 2013).
- World Health Organization. Serological detection of avian influenza A (H7N9) infections by microneutralization assay. World Health Organization 2013 May. Available online: http://www.who.int/influenza/gisrs_laboratory/cnic_serological_diagnosis_microneutralization_a_h7n9.pdf (accessed on 1 December 2013).
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Lu, J.; Chen, Y.; Shi, F.; Yu, H.; Huang, C.; Cui, L.; Shi, Z.; Jiao, Y.; Hu, Y. Characterization of Humoral Responses Induced by an H7N9 Influenza Virus-Like Particle Vaccine in BALB/C Mice. Viruses 2015, 7, 4369-4384. https://doi.org/10.3390/v7082821
Zhang L, Lu J, Chen Y, Shi F, Yu H, Huang C, Cui L, Shi Z, Jiao Y, Hu Y. Characterization of Humoral Responses Induced by an H7N9 Influenza Virus-Like Particle Vaccine in BALB/C Mice. Viruses. 2015; 7(8):4369-4384. https://doi.org/10.3390/v7082821
Chicago/Turabian StyleZhang, Li, Jing Lu, Yin Chen, Fengjuan Shi, Huiyan Yu, Chao Huang, Lunbiao Cui, Zhiyang Shi, Yongjun Jiao, and Yuemei Hu. 2015. "Characterization of Humoral Responses Induced by an H7N9 Influenza Virus-Like Particle Vaccine in BALB/C Mice" Viruses 7, no. 8: 4369-4384. https://doi.org/10.3390/v7082821