
Plant Virus Infection and the Ubiquitin Proteasome Machinery: Arms Race along the Endoplasmic Reticulum


{kind=link}
{kind=link}
Abstract
:1. Introduction
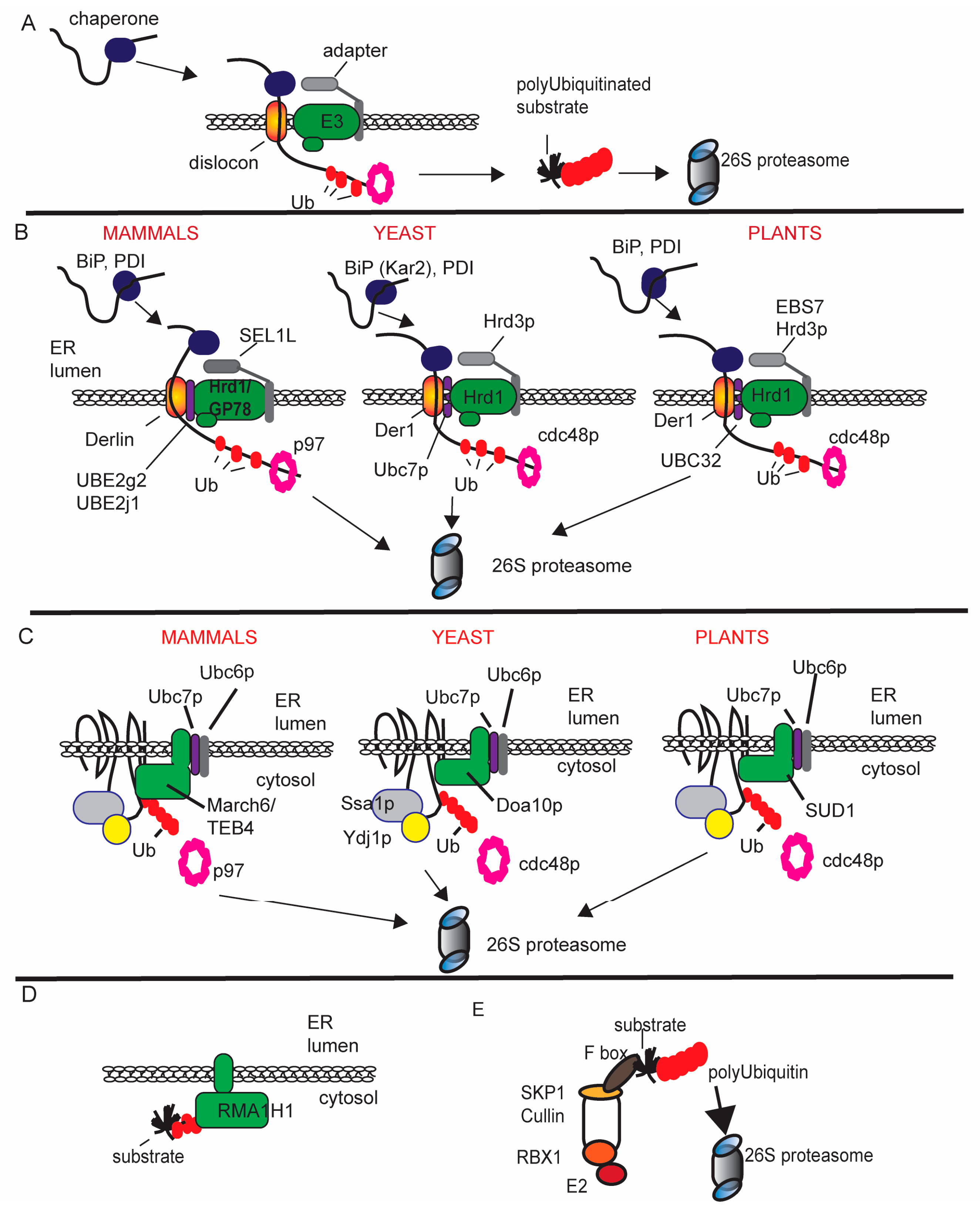
2. Ubiquitin Proteasome System Supporting Virus Replication and Movement
3. Hijacking the Host E3 Ub-Ligase Using Viral-Encoded F Box Proteins
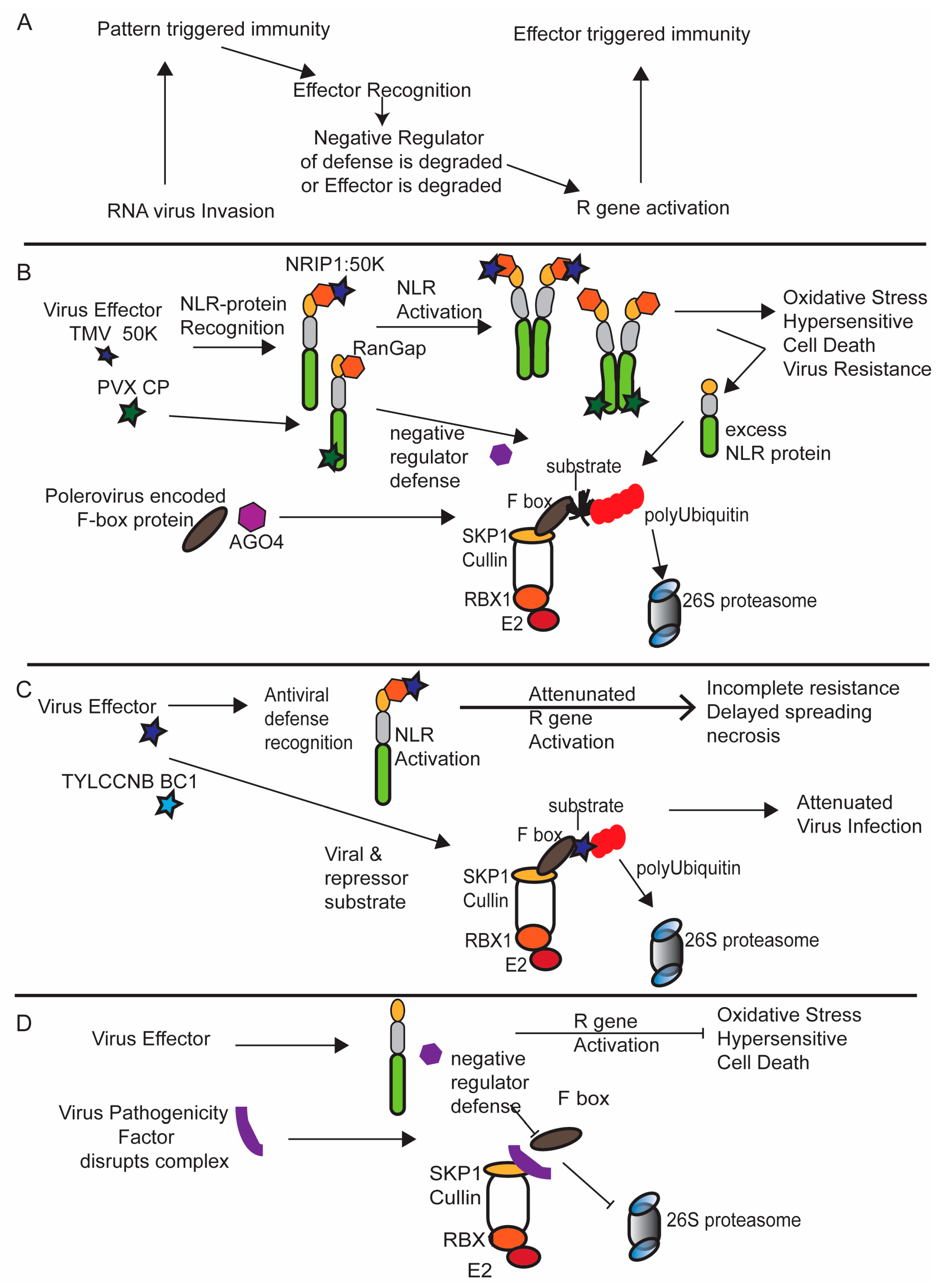
4. The Ubiquitin Ligase 26S Proteasome System and the Molecular Arms Race
5. Conclusion
Acknowledgments
Conflicts of Interest
References
- Duplan, V.; Rivas, S. E3 ubiquitin-ligases and their target proteins during the regulation of plant innate immunity. Front. Plant Sci. 2014, 5, 42. [Google Scholar] [CrossRef] [PubMed]
- Ryu, M.Y.; Cho, S.K.; Kim, W.T. The arabidopsis c3h2c3-type ring e3 ubiquitin ligase atairp1 is a positive regulator of an abscisic acid-dependent response to drought stress. Plant Physiol. 2010, 154, 1983–1997. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Choi, H.W.; Hwang, B.K. The pepper e3 ubiquitin ligase ring1 gene, caring1, is required for cell death and the salicylic acid-dependent defense response. Plant Physiol. 2011, 156, 2011–2025. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.K.; Cho, S.K.; Son, O.; Xu, Z.; Hwang, I.; Kim, W.T. Drought stress-induced rma1h1, a ring membrane-anchor e3 ubiquitin ligase homolog, regulates aquaporin levels via ubiquitination in transgenic arabidopsis plants. Plant Cell 2009, 21, 622–641. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, M.; Shirasu, K. Ubiquitination in plant immunity. Curr. Opin. Plant Biol. 2010, 13, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Metzger, M.B.; Hristova, V.A.; Weissman, A.M. Hect and ring finger families of e3 ubiquitin ligases at a glance. J. Cell Sci. 2012, 125, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.L.; Hauksdottir, H.; Troy, A.; Herschleb, J.; Kraft, E.; Callis, J. Functional analysis of the ring-type ubiquitin ligase family of arabidopsis. Plant Physiol. 2005, 137, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.X.; Howell, S.H. Managing the protein folding demands in the endoplasmic reticulum of plants. New Phytol. 2016, 211, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, J. Endoplasmic reticulum-mediated protein quality control in arabidopsis. Front. Plant Sci. 2014, 5, 162. [Google Scholar] [CrossRef] [PubMed]
- Horimoto, S.; Ninagawa, S.; Okada, T.; Koba, H.; Sugimoto, T.; Kamiya, Y.; Kato, K.; Takeda, S.; Mori, K. The unfolded protein response transducer atf6 represents a novel transmembrane-type endoplasmic reticulum-associated degradation substrate requiring both mannose trimming and sel1l protein. J. Biol. Chem. 2013, 288, 31517–31527. [Google Scholar] [CrossRef] [PubMed]
- Doblas, V.G.; Amorim-Silva, V.; Pose, D.; Rosado, A.; Esteban, A.; Arro, M.; Azevedo, H.; Bombarely, A.; Borsani, O.; Valpuesta, V.; et al. The sud1 gene encodes a putative e3 ubiquitin ligase and is a positive regulator of 3-hydroxy-3-methylglutaryl coenzyme a reductase activity in arabidopsis. Plant Cell 2013, 25, 728–743. [Google Scholar] [CrossRef] [PubMed]
- Muller, J.; Piffanelli, P.; Devoto, A.; Miklis, M.; Elliott, C.; Ortmann, B.; Schulze-Lefert, P.; Panstruga, R. Conserved erad-like quality control of a plant polytopic membrane protein. Plant Cell 2005, 17, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, C.; Wang, D.; Su, W.; Liu, L.; Wang, M.; Li, J. Ebs7 is a plant-specific component of a highly conserved endoplasmic reticulum-associated degradation system in arabidopsis. Proc. Natl. Acad. Sci. USA 2015, 112, 12205–12210. [Google Scholar] [CrossRef] [PubMed]
- Van Hoewyk, D. Defects in endoplasmic reticulum-associated degradation (erad) increase selenate sensitivity in arabidopsis. Plant Signal. Behav. 2016. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Fang, S. Live cell imaging of protein dislocation from the endoplasmic reticulum. J. Biol. Chem. 2012, 287, 28057–28066. [Google Scholar] [CrossRef] [PubMed]
- Huttner, S.; Veit, C.; Schoberer, J.; Grass, J.; Strasser, R. Unraveling the function of arabidopsis thaliana os9 in the endoplasmic reticulum-associated degradation of glycoproteins. Plant Mol. Biol. 2012, 79, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Huttner, S.; Veit, C.; Vavra, U.; Schoberer, J.; Liebminger, E.; Maresch, D.; Grass, J.; Altmann, F.; Mach, L.; Strasser, R. Arabidopsis class i alpha-mannosidases mns4 and mns5 are involved in endoplasmic reticulum-associated degradation of misfolded glycoproteins. Plant Cell 2014, 26, 1712–1728. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zhong, Y.; Wu, Y.; Liu, L.; Wang, P.; Liu, R.; Cui, F.; Li, Q.; Yang, X.; Fang, S.; et al. Hrd1-mediated erad tuning of er-bound e2 is conserved between plants and mammals. Nat. Plants 2016, 2, 16094. [Google Scholar] [CrossRef] [PubMed]
- Gilon, T.; Chomsky, O.; Kulka, R.G. Degradation signals for ubiquitin system proteolysis in saccharomyces cerevisiae. EMBO J. 1998, 17, 2759–2766. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Liu, W.; Zhang, H.; Liu, C.; Shang, Y.; Ye, Y.; Zhang, X.; Li, W. Ube2g2-gp78-mediated herp polyubiquitylation is involved in er stress recovery. J. Cell Sci. 2014, 127, 1417–1427. [Google Scholar] [CrossRef] [PubMed]
- Bachmair, A.; Novatchkova, M.; Potuschak, T.; Eisenhaber, F. Ubiquitylation in plants: A post-genomic look at a post-translational modification. Trends Plant Sci. 2001, 6, 463–470. [Google Scholar] [CrossRef]
- Bernasconi, R.; Galli, C.; Kokame, K.; Molinari, M. Autoadaptive er-associated degradation defines a preemptive unfolded protein response pathway. Mol. Cell 2013. [Google Scholar] [CrossRef] [PubMed]
- Yen, J.L.; Flick, K.; Papagiannis, C.V.; Mathur, R.; Tyrrell, A.; Ouni, I.; Kaake, R.M.; Huang, L.; Kaiser, P. Signal-induced disassembly of the scf ubiquitin ligase complex by cdc48/p97. Mol. Cell 2012, 48, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, S.; Brodsky, J.L.; Nakatsukasa, K. Roles of molecular chaperones in endoplasmic reticulum (er) quality control and er-associated degradation (erad). J. Biochem. 2005, 137, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Copeland, C.; Woloshen, V.; Huang, Y.; Li, X. Atcdc48a is involved in the turnover of an nlr immune receptor. Plant J. 2016. [Google Scholar] [CrossRef] [PubMed]
- Verchot, J. The er quality control and er associated degradation machineries are vital for viral pathogenesis. Front. Plant Sci. 2014, 5, 66. [Google Scholar] [CrossRef] [PubMed]
- Byun, H.; Gou, Y.; Zook, A.; Lozano, M.M.; Dudley, J.P. Erad and how viruses exploit it. Front. Microbiol. 2014, 5, 330. [Google Scholar] [CrossRef] [PubMed]
- Alcaide-Loridan, C.; Jupin, I. Ubiquitin and plant viruses, let’s play together! Plant Physiol. 2012, 160, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Barajas, D.; Li, Z.; Nagy, P.D. The nedd4-type rsp5p ubiquitin ligase inhibits tombusvirus replication by regulating degradation of the p92 replication protein and decreasing the activity of the tombusvirus replicase. J. Virol. 2009, 83, 11751–11764. [Google Scholar] [CrossRef] [PubMed]
- Barajas, D.; Nagy, P.D. Ubiquitination of tombusvirus p33 replication protein plays a role in virus replication and binding to the host vps23p escrt protein. Virology 2010, 397, 358–368. [Google Scholar] [CrossRef] [PubMed]
- Imura, Y.; Molho, M.; Chuang, C.; Nagy, P.D. Cellular ubc2/rad6 e2 ubiquitin-conjugating enzyme facilitates tombusvirus replication in yeast and plants. Virology 2015, 484, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Sasvari, Z.; Alatriste Gonzalez, P.; Nagy, P.D. Tombusvirus-yeast interactions identify conserved cell-intrinsic viral restriction factors. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.D. The roles of host factors in tombusvirus rna recombination. Adv. Virus Res. 2011, 81, 63–84. [Google Scholar] [PubMed]
- Sasvari, Z.; Nagy, P.D. Making of viral replication organelles by remodeling interior membranes. Viruses 2010, 2, 2436–2442. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Barajas, D.; Panavas, T.; Herbst, D.A.; Nagy, P.D. Cdc34p ubiquitin-conjugating enzyme is a component of the tombusvirus replicase complex and ubiquitinates p33 replication protein. J. Virol. 2008, 82, 6911–6926. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Liu, L.; Zhao, Q.; Zhang, Z.; Li, Q.; Lin, B.; Wu, Y.; Tang, S.; Xie, Q. Arabidopsis ubiquitin conjugase ubc32 is an erad component that functions in brassinosteroid-mediated salt stress tolerance. Plant Cell 2012, 24, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Barajas, D.; Jiang, Y.; Nagy, P.D. A unique role for the host escrt proteins in replication of tomato bushy stunt virus. PLoS Pathog. 2009, 5, e1000705. [Google Scholar] [CrossRef] [PubMed]
- Prasanth, K.R.; Barajas, D.; Nagy, P.D. The proteasomal rpn11 metalloprotease suppresses tombusvirus rna recombination and promotes viral replication via facilitating assembly of the viral replicase complex. J. Virol. 2015, 89, 2750–2763. [Google Scholar] [CrossRef] [PubMed]
- Feldman, M.; van der Goot, F.G. Novel ubiquitin-dependent quality control in the endoplasmic reticulum. Trends Cell Biol. 2009, 19, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Kovalev, N.; Pogany, J.; Nagy, P.D. A co-opted dead-box rna helicase enhances tombusvirus plus-strand synthesis. PLoS Pathog. 2012, 8, e1002537. [Google Scholar] [CrossRef] [PubMed]
- Linder, P. Yeast rna helicases of the dead-box family involved in translation initiation. Biol. Cell 2003, 95, 157–167. [Google Scholar] [CrossRef]
- Prod’homme, D.; Jakubiec, A.; Tournier, V.; Drugeon, G.; Jupin, I. Targeting of the turnip yellow mosaic virus 66k replication protein to the chloroplast envelope is mediated by the 140k protein. J. Virol. 2003, 77, 9124–9135. [Google Scholar] [CrossRef] [PubMed]
- Camborde, L.; Planchais, S.; Tournier, V.; Jakubiec, A.; Drugeon, G.; Lacassagne, E.; Pflieger, S.; Chenon, M.; Jupin, I. The ubiquitin-proteasome system regulates the accumulation of turnip yellow mosaic virus rna-dependent rna polymerase during viral infection. Plant Cell 2010, 22, 3142–3152. [Google Scholar] [CrossRef] [PubMed]
- Drugeon, G.; Jupin, I. Stability in vitro of the 69k movement protein of turnip yellow mosaic virus is regulated by the ubiquitin-mediated proteasome pathway. J. Gen. Virol. 2002, 83, 3187–3197. [Google Scholar] [CrossRef] [PubMed]
- Dunigan, D.D.; Dietzgen, R.G.; Schoelz, J.E.; Zaitlin, M. Tobacco mosaic virus particles contain ubiquitinated coat protein subunits. Virology 1988, 165, 310–312. [Google Scholar] [CrossRef]
- Jockusch, H.; Wiegand, C. Misfolded plant virus proteins: Elicitors and targets of ubiquitylation. FEBS Lett. 2003, 545, 229–232. [Google Scholar] [CrossRef]
- Reichel, C.; Beachy, R.N. Degradation of Tobacco mosaic virus movement protein by the 26s proteasome. J. Virol. 2000, 74, 3330–3337. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Dickman, M.B.; Whitham, S.A.; Payton, M.; Verchot, J. The unfolded protein response is triggered by a plant viral movement protein. Plant Physiol. 2011, 156, 741–755. [Google Scholar] [CrossRef] [PubMed]
- Ju, H.J.; Ye, C.M.; Verchot-Lubicz, J. Mutational analysis of pvx tgbp3 links subcellular accumulation and protein turnover. Virology 2008, 375, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.M.; Chen, S.; Payton, M.; Dickman, M.B.; Verchot, J. Tgbp3 triggers the unfolded protein response and skp1-dependent programmed cell death. Mol. Plant Pathol. 2013, 14, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Jang, J. F-box proteins in arabidopsis. Trends Plant Sci. 2000, 5, 454–457. [Google Scholar] [CrossRef]
- Yu, H.; Moss, B.L.; Jang, S.S.; Prigge, M.; Klavins, E.; Nemhauser, J.L.; Estelle, M. Mutations in the tir1 auxin receptor that increase affinity for auxin/indole-3-acetic acid proteins result in auxin hypersensitivity. Plant Physiol. 2013, 162, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Neve, J.; Hirose, M.; Kuboki, A.; Shimada, Y.; Kepinski, S.; Nozaki, H. Rational design of an auxin antagonist of the scf(tir1) auxin receptor complex. ACS Chem. Biol. 2012, 7, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Maraschin Fdos, S.; Memelink, J.; Offringa, R. Auxin-induced, scf(tir1)-mediated poly-ubiquitination marks aux/iaa proteins for degradation. Plant J. 2009, 59, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Zheng, N. Hormone signaling through protein destruction: A lesson from plants. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E223–E227. [Google Scholar] [CrossRef] [PubMed]
- Peltier, C.; Hleibieh, K.; Thiel, H.; Klein, E.; Bragard, C.; Gilmer, D. Molecular biology of the beet necrotic yellow vein virus. Plant Viruses 2008, 2, 14–24. [Google Scholar]
- Peltier, C.; Schmidlin, L.; Klein, E.; Taconnat, L.; Prinsen, E.; Erhardt, M.; Heintz, D.; Weyens, G.; Lefebvre, M.; Renou, J.P.; et al. Expression of the beet necrotic yellow vein virus p25 protein induces hormonal changes and a root branching phenotype in arabidopsis thaliana. Transgenic Res. 2011, 20, 443–466. [Google Scholar] [CrossRef] [PubMed]
- Thiel, H.; Hleibieh, K.; Gilmer, D.; Varrelmann, M. The p25 pathogenicity factor of beet necrotic yellow vein virus targets the sugar beet 26s proteasome involved in the induction of a hypersensitive resistance response via interaction with an f-box protein. Mol. Plant Microbe Interact. 2012, 25, 1058–1072. [Google Scholar] [CrossRef] [PubMed]
- Fusaro, A.F.; Correa, R.L.; Nakasugi, K.; Jackson, C.; Kawchuk, L.; Vaslin, M.F.; Waterhouse, P.M. The enamovirus p0 protein is a silencing suppressor which inhibits local and systemic rna silencing through ago1 degradation. Virology 2012, 426, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Pazhouhandeh, M.; Dieterle, M.; Marrocco, K.; Lechner, E.; Berry, B.; Brault, V.; Hemmer, O.; Kretsch, T.; Richards, K.E.; Genschik, P.; et al. F-box-like domain in the polerovirus protein p0 is required for silencing suppressor function. Proc. Natl. Acad. Sci. USA 2006, 103, 1994–1999. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Tao, T.; Han, Y.; Chen, X.; Fan, Z.; Li, D.; Yu, J.; Han, C. Nonstructural protein p7–2 encoded by rice black-streaked dwarf virus interacts with skp1, a core subunit of scf ubiquitin ligase. Virol. J. 2013, 10, 325. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, E.; Zilber-Rosenberg, I. Microbes drive evolution of animals and plants: The hologenome concept. mBio 2016, 7, e01395-15. [Google Scholar] [CrossRef] [PubMed]
- Dielen, A.S.; Badaoui, S.; Candresse, T.; German-Retana, S. The ubiquitin/26s proteasome system in plant–pathogen interactions: A never-ending hide-and-seek game. Mol. Plant Pathol. 2010, 11, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Bonardi, V.; Dangl, J.L. How complex are intracellular immune receptor signaling complexes? Front. Plant Sci. 2012, 3, 237. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, J.M. Plant immunity triggered by microbial molecular signatures. Mol. Plant 2010, 3, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Lecellier, C.H.; Voinnet, O. Rna silencing: No mercy for viruses? Immunol. Rev. 2004, 198, 285–303. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. Rna silencing as a plant immune system against viruses. Trends Genet. 2001, 17, 449–459. [Google Scholar] [CrossRef]
- Pfeffer, S.; Dunoyer, P.; Heim, F.; Richards, K.E.; Jonard, G.; Ziegler-Graff, V. P0 of beet western yellows virus is a suppressor of posttranscriptional gene silencing. J. Virol. 2002, 76, 6815–6824. [Google Scholar] [CrossRef] [PubMed]
- Bortolamiol, D.; Pazhouhandeh, M.; Marrocco, K.; Genschik, P.; Ziegler-Graff, V. The polerovirus f box protein p0 targets argonaute1 to suppress rna silencing. Curr. Biol. 2007, 17, 1615–1621. [Google Scholar] [CrossRef] [PubMed]
- Kozlowska-Makulska, A.; Guilley, H.; Szyndel, M.S.; Beuve, M.; Lemaire, O.; Herrbach, E.; Bouzoubaa, S. P0 proteins of european beet-infecting poleroviruses display variable rna silencing suppression activity. J. Gen. Virol. 2010, 91, 1082–1091. [Google Scholar] [CrossRef] [PubMed]
- Baumberger, N.; Tsai, C.H.; Lie, M.; Havecker, E.; Baulcombe, D.C. The polerovirus silencing suppressor p0 targets argonaute proteins for degradation. Curr. Biol. 2007, 17, 1609–1614. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Ma, D.; Dong, J.; Jin, J.; Li, D.; Deng, C.; Wang, T. Hc-pro protein of potato virus y can interact with three arabidopsis 20s proteasome subunits in planta. J. Virol. 2007, 81, 12881–12888. [Google Scholar] [CrossRef] [PubMed]
- Sahana, N.; Kaur, H.; Basavaraj; Tena, F.; Jain, R.K.; Palukaitis, P.; Canto, T.; Praveen, S. Inhibition of the host proteasome facilitates papaya ringspot virus accumulation and proteosomal catalytic activity is modulated by viral factor hcpro. PLoS ONE 2012, 7, e52546. [Google Scholar] [CrossRef] [PubMed]
- Dielen, A.S.; Sassaki, F.T.; Walter, J.; Michon, T.; Menard, G.; Pagny, G.; Krause-Sakate, R.; Maia Ide, G.; Badaoui, S.; Le Gall, O.; et al. The 20s proteasome alpha5 subunit of arabidopsis thaliana carries an rnase activity and interacts in planta with the lettuce mosaic potyvirus hcpro protein. Mol. Plant Pathol. 2011, 12, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Ballut, L.; Drucker, M.; Pugniere, M.; Cambon, F.; Blanc, S.; Roquet, F.; Candresse, T.; Schmid, H.P.; Nicolas, P.; Gall, O.L.; et al. Hcpro, a multifunctional protein encoded by a plant rna virus, targets the 20s proteasome and affects its enzymic activities. J. Gen. Virol. 2005, 86, 2595–2603. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Schiff, M.; Serino, G.; Deng, X.W.; Dinesh-Kumar, S.P. Role of scf ubiquitin-ligase and the cop9 signalosome in the n gene-mediated resistance response to Tobacco mosaic virus. Plant Cell 2002, 14, 1483–1496. [Google Scholar] [CrossRef] [PubMed]
- Mestre, P.; Baulcombe, D.C. Elicitor-mediated oligomerization of the tobacco n disease resistance protein. Plant Cell 2006, 18, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Schiff, M.; Marathe, R.; Dinesh-Kumar, S.P. Tobacco rar1, eds1 and npr1/nim1 like genes are required for n-mediated resistance to Tobacco mosaic virus. Plant J. 2002, 30, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Duran, R.; Rosas-Diaz, T.; Gusmaroli, G.; Luna, A.P.; Taconnat, L.; Deng, X.W.; Bejarano, E.R. Geminiviruses subvert ubiquitination by altering csn-mediated derubylation of scf e3 ligase complexes and inhibit jasmonate signaling in arabidopsis thaliana. Plant Cell 2011, 23, 1014–1032. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Duran, R.; Bejarano, E.R. Geminivirus c2 protein might be the key player for geminiviral co-option of scf-mediated ubiquitination. Plant Signal. Behav. 2011, 6, 999–1001. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Hu, T.; Bao, M.; Cao, L.; Zhang, H.; Song, F.; Xie, Q.; Zhou, X. Tobacco ring e3 ligase ntrfp1 mediates ubiquitination and proteasomal degradation of a geminivirus-encoded betac1. Mol. Plant 2016, 9, 911–925. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Bao, M.; Zhou, X. A plant kinase plays roles in defense response against geminivirus by phosphorylation of a viral pathogenesis protein. Plant Signal. Behav. 2012, 7, 888–892. [Google Scholar] [CrossRef] [PubMed]


© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verchot, J. Plant Virus Infection and the Ubiquitin Proteasome Machinery: Arms Race along the Endoplasmic Reticulum. Viruses 2016, 8, 314. https://doi.org/10.3390/v8110314
Verchot J. Plant Virus Infection and the Ubiquitin Proteasome Machinery: Arms Race along the Endoplasmic Reticulum. Viruses. 2016; 8(11):314. https://doi.org/10.3390/v8110314
Chicago/Turabian StyleVerchot, Jeanmarie. 2016. "Plant Virus Infection and the Ubiquitin Proteasome Machinery: Arms Race along the Endoplasmic Reticulum" Viruses 8, no. 11: 314. https://doi.org/10.3390/v8110314
APA StyleVerchot, J. (2016). Plant Virus Infection and the Ubiquitin Proteasome Machinery: Arms Race along the Endoplasmic Reticulum. Viruses, 8(11), 314. https://doi.org/10.3390/v8110314