
Recycling Endosomes and Viral Infection


Abstract
:1. Introduction
2. Recycling Pathways Integrate the Endomembrane System
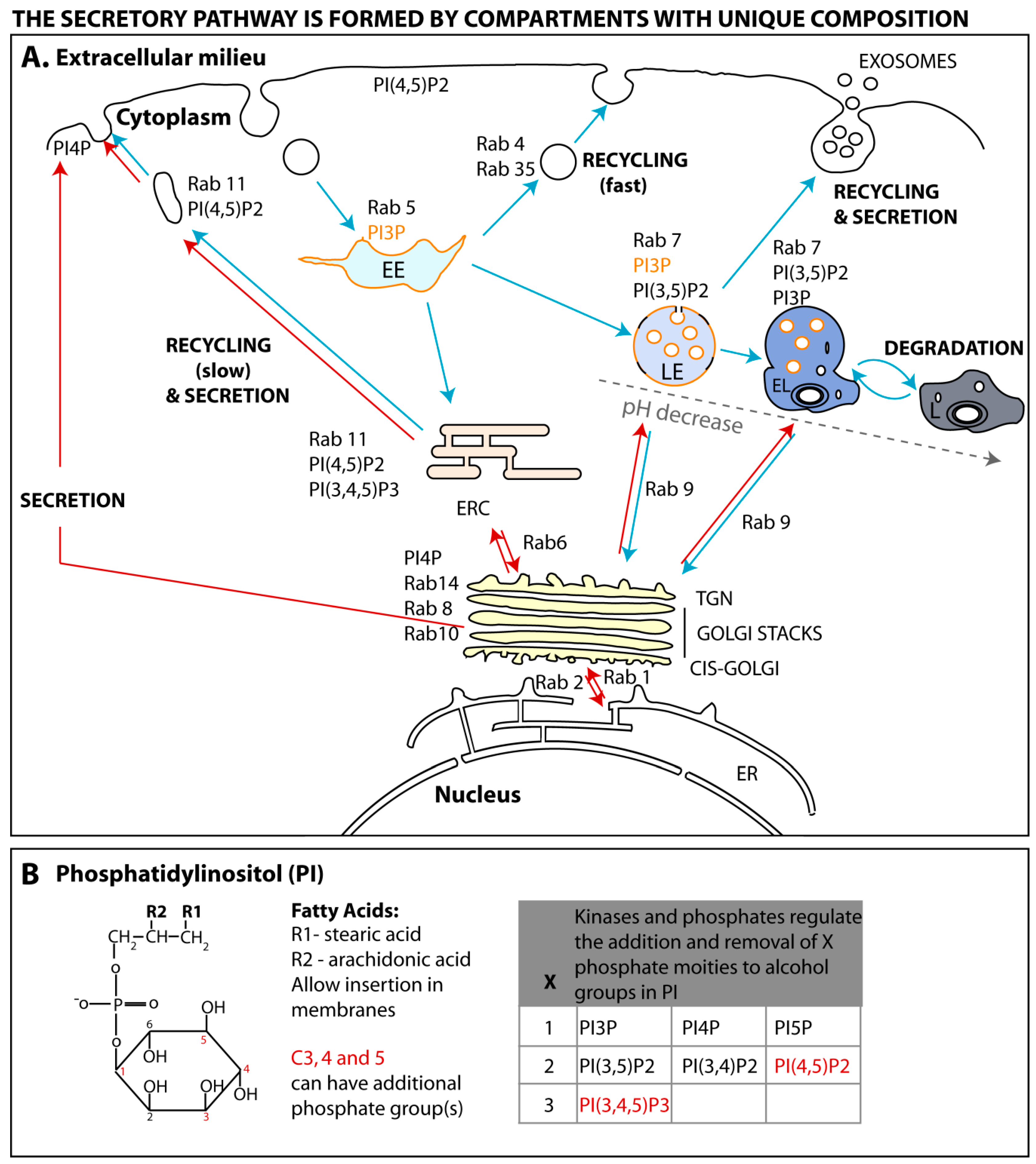
3. The Endomembrane System Is Formed by Compartments of Unique Composition
3.1. Rab GTPases
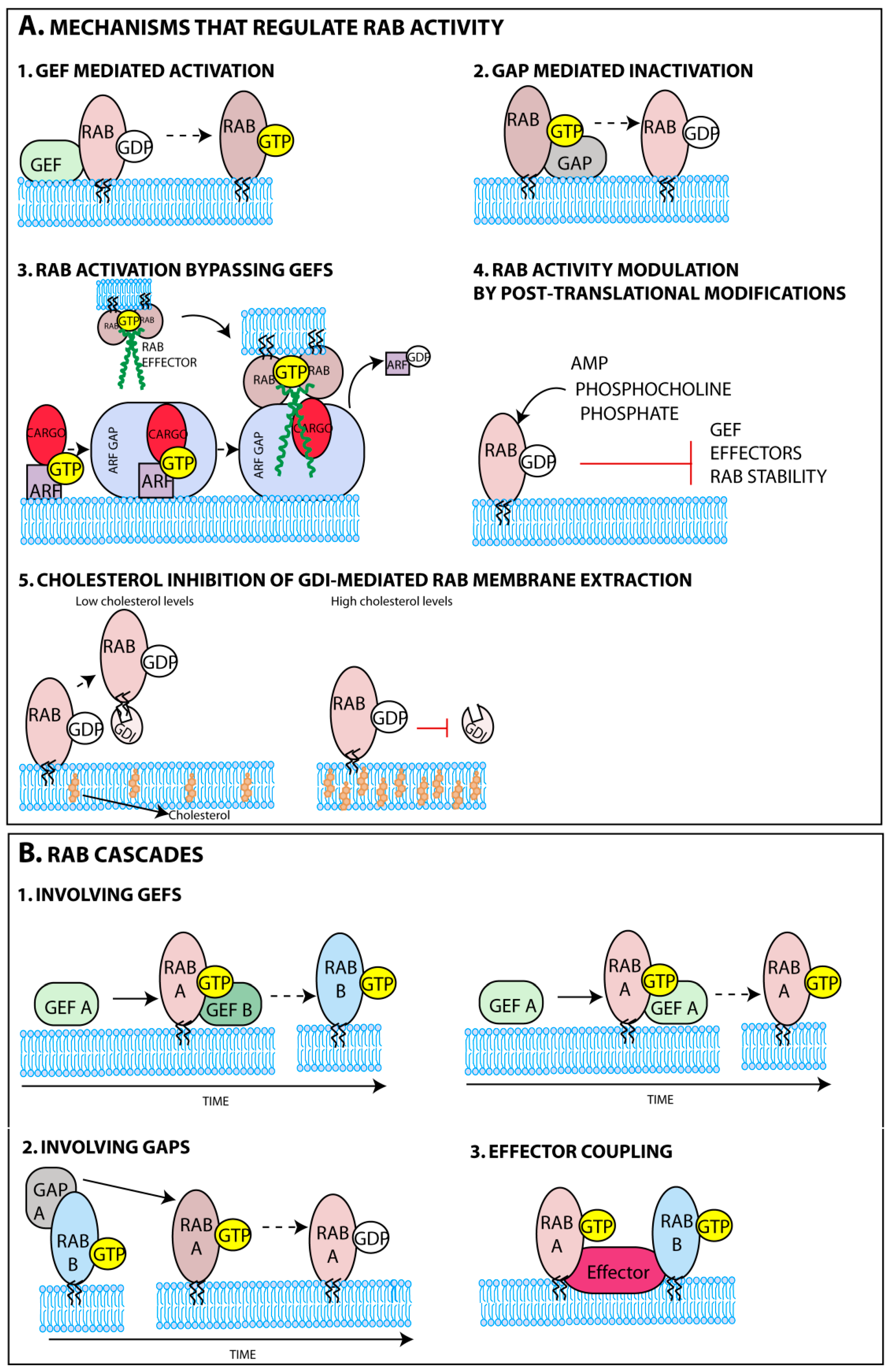
3.1.1. Rab GTPase Spatial Regulation
3.1.2. Rab GTPase Temporal Regulation
- Defining the chronology of Rab cascades;
- Quantifying the number and diversity of Rab-mediated molecular interactions when bound to GDP or GTP;
- Identifying regulatory mechanisms, including GEFs and GAPs;
- Exploring the crosstalk between signaling/metabolic pathways and Rab function;
- Determining the kinetics and composition of sequential protein scaffolds coordinating movement, tethering and fusion events.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
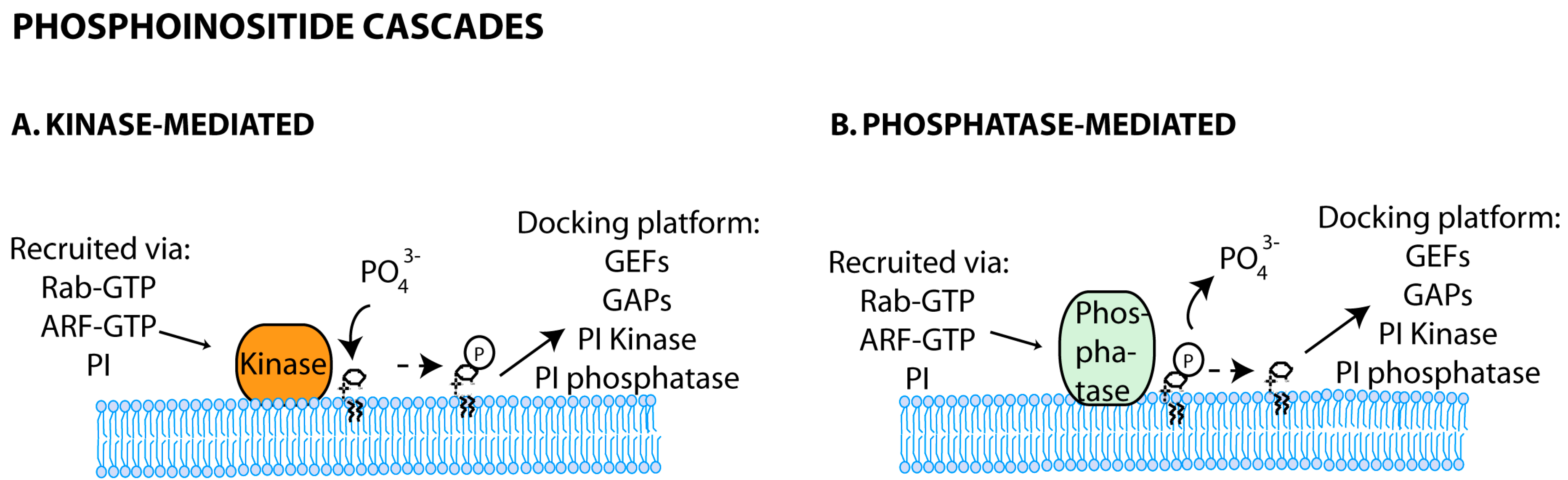
3.2. Phosphoinositide Code
3.2.1. Phosphoinositide Feed-Forward Loops and Cascades
3.2.2. Phosphoinositide and Rab Crosstalk
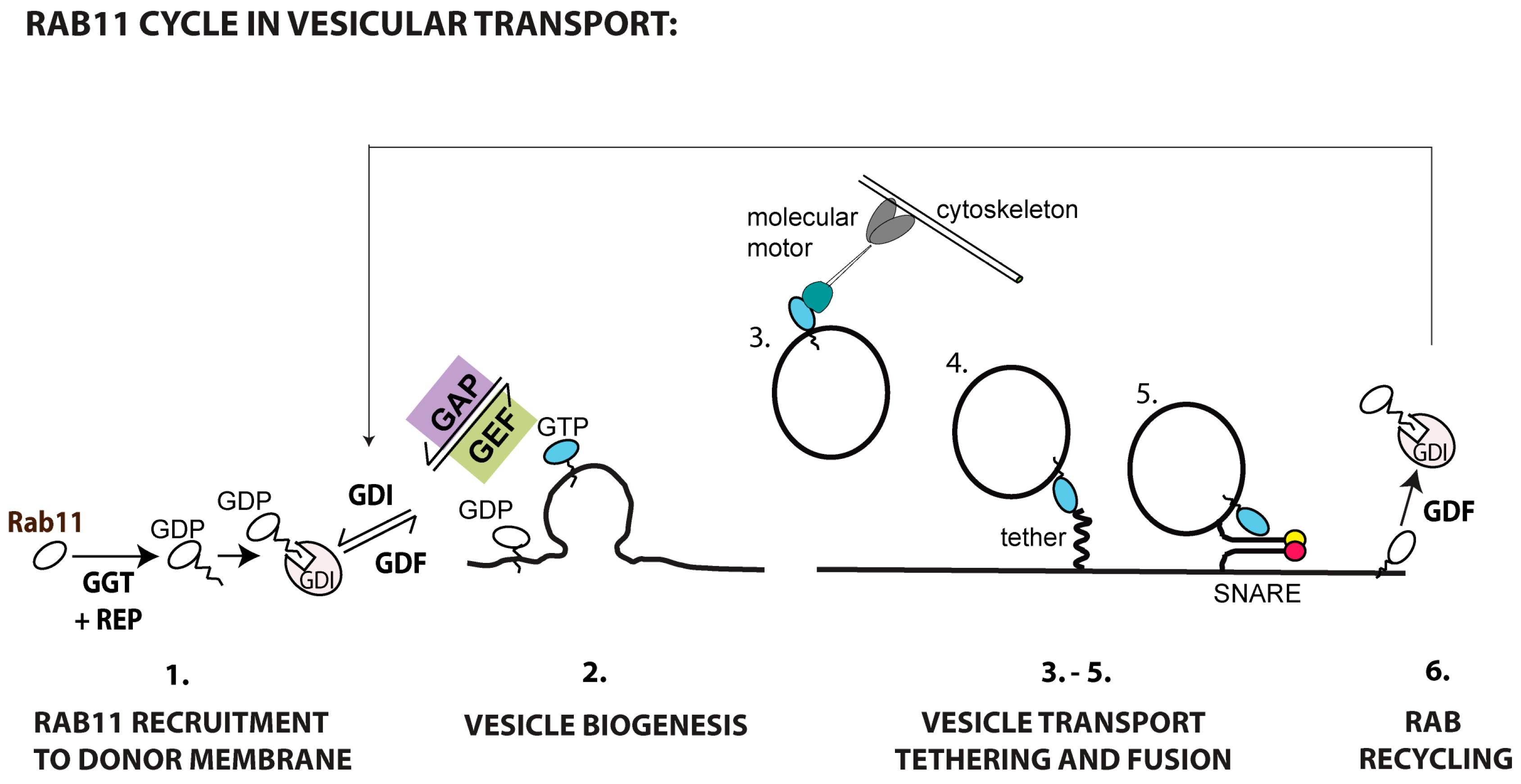
4. The Rab11 Centric View of the ERC
4.1. Rab11 Membrane Recruitment and Activation
4.2. Rab11 Recruitment of Effectors
4.2.1. Rab11 Effectors Involved in Rab Cascades
4.2.2. The Crosstalk between Rab11 and the Cytoskeleton
4.2.3. Rab11 Vesicular Targeting and Fusion with the Target Membrane
4.3. Rab11 Inactivation and Recycling
5. Involvement of ERC in Viral Lifecycles
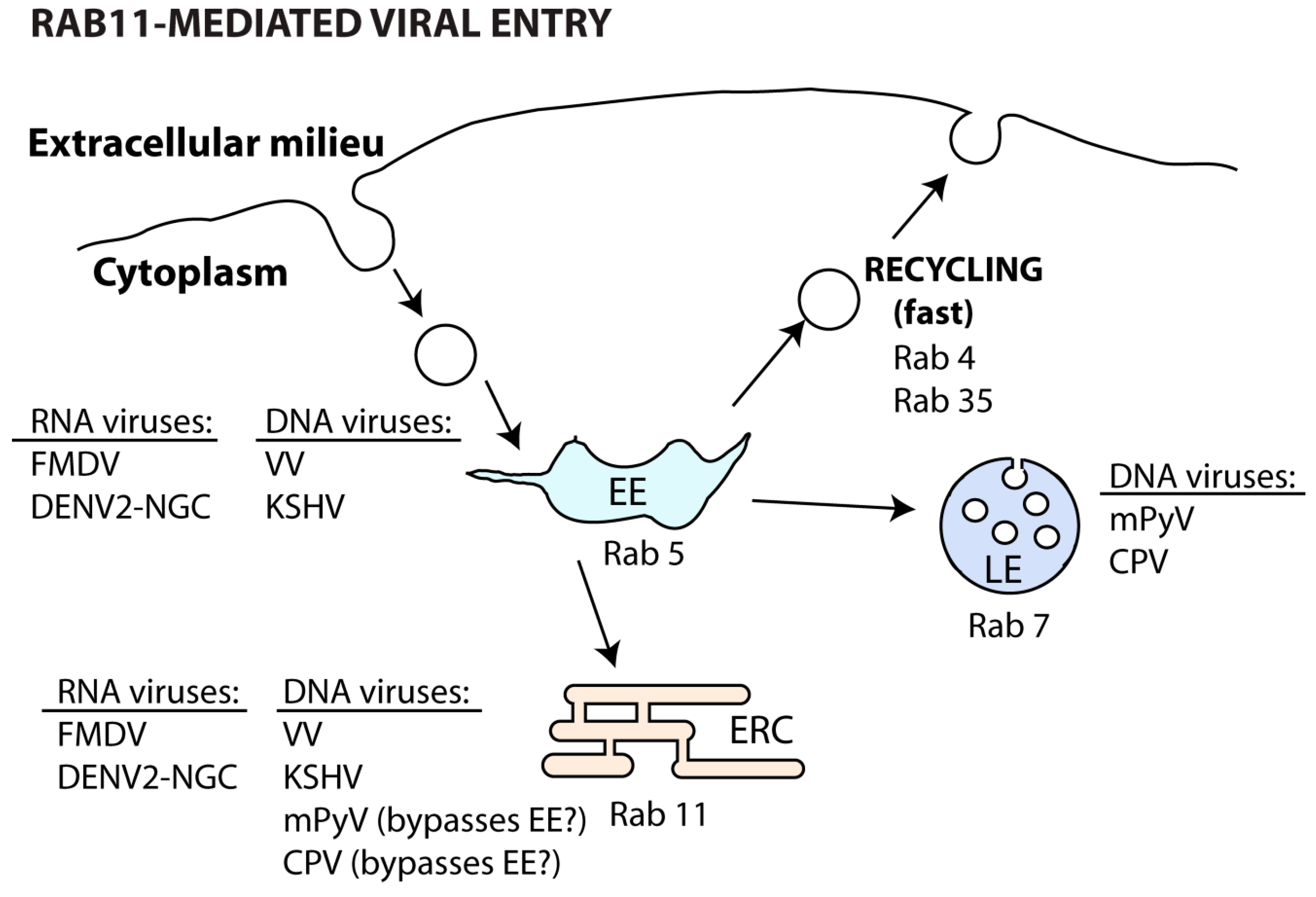
5.1. Viral Entrance
5.2. Viral Assembly
5.3. Viral Budding and Release
6. Cellular Alterations Resulting from the Interplay between the ERC and Viruses
6.1. Cytoskeleton Alterations
6.2. Host Antiviral Response
6.3. Membrane Remodeling
7. Future Perspectives
Acknowledgments
Author Contributions
Conflict of Interests
References
- Harak, C.; Lohmann, V. Ultrastructure of the replication sites of positive-strand RNA viruses. Virology 2015, 479–480, 418–433. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.; Krijnse-Locker, J. Modification of intracellular membrane structures for virus replication. Nat. Rev. Microbiol. 2008, 6, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Reid, C.R.; Airo, A.M.; Hobman, T.C. The virus-host interplay: Biogenesis of +RNA replication complexes. Viruses 2015, 7, 4385–4413. [Google Scholar] [CrossRef] [PubMed]
- Howes, D.W.; Melnick, J.L.; Reissig, M. Sequence of morphological changes in epithelial cell cultures infected with poliovirus. J. Exp. Med. 1956, 104, 289–304. [Google Scholar] [PubMed]
- Robbins, F.C.; Enders, J.F.; Weller, T.H. The effect of poliomyelitis virus upon cells in tissue cultures. J. Clin. Investig. 1950, 29, 841. [Google Scholar] [PubMed]
- Condit, R.C.; Moussatche, N.; Traktman, P. In a nutshell: Structure and assembly of the vaccinia virion. Adv. Virus Res. 2006, 66, 31–124. [Google Scholar] [PubMed]
- Den Boon, J.A.; Diaz, A.; Ahlquist, P. Cytoplasmic viral replication complexes. Cell Host Microbe 2010, 8, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Krijnse Locker, J.; Chlanda, P.; Sachsenheimer, T.; Brugger, B. Poxvirus membrane biogenesis: Rupture not disruption. Cell. Microbiol. 2013, 15, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Miyanari, Y.; Atsuzawa, K.; Usuda, N.; Watashi, K.; Hishiki, T.; Zayas, M.; Bartenschlager, R.; Wakita, T.; Hijikata, M.; Shimotohno, K. The lipid droplet is an important organelle for hepatitis C virus production. Nat. Cell Biol. 2007, 9, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Welsch, S.; Miller, S.; Romero-Brey, I.; Merz, A.; Bleck, C.K.; Walther, P.; Fuller, S.D.; Antony, C.; Krijnse-Locker, J.; Bartenschlager, R. Composition and three-dimensional architecture of the dengue virus replication and assembly sites. Cell Host Microbe 2009, 5, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Bruce, E.A.; Stuart, A.; McCaffrey, M.W.; Digard, P. Role of the Rab11 pathway in negative-strand virus assembly. Biochem. Soc. Trans. 2012, 40, 1409–1415. [Google Scholar] [CrossRef] [PubMed]
- Kelly, E.E.; Horgan, C.P.; McCaffrey, M.W. Rab11 proteins in health and disease. Biochem. Soc. Trans. 2012, 40, 1360–1367. [Google Scholar] [CrossRef] [PubMed]
- Guichard, A.; Nizet, V.; Bier, E. Rab11-mediated trafficking in host-pathogen interactions. Nat. Rev. Microbiol. 2014, 12, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Hata, Y.; Slaughter, C.A.; Sudhof, T.C. Synaptic vesicle fusion complex contains unc-18 homologue bound to syntaxin. Nature 1993, 366, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, C.A.; Schekman, R. Distinct sets of SEC genes govern transport vesicle formation and fusion early in the secretory pathway. Cell 1990, 61, 723–733. [Google Scholar] [CrossRef]
- Balch, W.E.; Dunphy, W.G.; Braell, W.A.; Rothman, J.E. Reconstitution of the transport of protein between successive compartments of the golgi measured by the coupled incorporation of N-acetylglucosamine. Cell 1984, 39, 405–416. [Google Scholar] [CrossRef]
- Huotari, J.; Helenius, A. Endosome maturation. EMBO J. 2011, 30, 3481–3500. [Google Scholar] [CrossRef] [PubMed]
- Scott, C.C.; Vacca, F.; Gruenberg, J. Endosome maturation, transport and functions. Semin. Cell Dev. Biol. 2014, 31, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Doherty, G.J.; McMahon, H.T. Mechanisms of endocytosis. Annu. Rev. Biochem. 2009, 78, 857–902. [Google Scholar] [CrossRef] [PubMed]
- Grant, B.D.; Donaldson, J.G. Pathways and mechanisms of endocytic recycling. Nat. Rev. Mol. Cell Biol. 2009, 10, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Steinman, R.M.; Mellman, I.S.; Muller, W.A.; Cohn, Z.A. Endocytosis and the recycling of plasma membrane. J. Cell Biol. 1983, 96, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Feng, Y.; Chen, D.; Wandinger-Ness, A. Rab11 is required for trans-golgi network-to-plasma membrane transport and a preferential target for GDP dissociation inhibitor. Mol. Biol. Cell 1998, 9, 3241–3257. [Google Scholar] [CrossRef] [PubMed]
- Ang, A.L.; Taguchi, T.; Francis, S.; Folsch, H.; Murrells, L.J.; Pypaert, M.; Warren, G.; Mellman, I. Recycling endosomes can serve as intermediates during transport from the golgi to the plasma membrane of MDCK cells. J. Cell Biol. 2004, 167, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.Z.; Kay, J.G.; Sangermani, D.G.; Stow, J.L. A role for the phagosome in cytokine secretion. Science 2005, 310, 1492–1495. [Google Scholar] [CrossRef] [PubMed]
- Maxfield, F.R.; McGraw, T.E. Endocytic recycling. Nat. Rev. Mol. Cell Biol. 2004, 5, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Lock, J.G.; Stow, J.L. Rab11 in recycling endosomes regulates the sorting and basolateral transport of e-cadherin. Mol. Biol. Cell 2005, 16, 1744–1755. [Google Scholar] [CrossRef] [PubMed]
- Casanova, J.E.; Wang, X.; Kumar, R.; Bhartur, S.G.; Navarre, J.; Woodrum, J.E.; Altschuler, Y.; Ray, G.S.; Goldenring, J.R. Association of Rab25 and Rab11a with the apical recycling system of polarized madin-darby canine kidney cells. Mol. Biol. Cell 1999, 10, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Goldenring, J.R.; Smith, J.; Vaughan, H.D.; Cameron, P.; Hawkins, W.; Navarre, J. Rab11 is an apically located small GTP-binding protein in epithelial tissues. Am. J. Physiol. 1996, 270, G515–G525. [Google Scholar] [PubMed]
- Sobajima, T.; Yoshimura, S.; Iwano, T.; Kunii, M.; Watanabe, M.; Atik, N.; Mushiake, S.; Morii, E.; Koyama, Y.; Miyoshi, E.; et al. Rab11a is required for apical protein localisation in the intestine. Biol. Open 2014, 4, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Riggs, B.; Rothwell, W.; Mische, S.; Hickson, G.R.; Matheson, J.; Hays, T.S.; Gould, G.W.; Sullivan, W. Actin cytoskeleton remodeling during early drosophila furrow formation requires recycling endosomal components nuclear-fallout and Rab11. J. Cell Biol. 2003, 163, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Skop, A.R.; Bergmann, D.; Mohler, W.A.; White, J.G. Completion of cytokinesis in C. elegans requires a brefeldin a-sensitive membrane accumulation at the cleavage furrow apex. Curr. Biol. 2001, 11, 735–746. [Google Scholar] [CrossRef]
- Park, M.; Penick, E.C.; Edwards, J.G.; Kauer, J.A.; Ehlers, M.D. Recycling endosomes supply AMPA receptors for LTP. Science 2004, 305, 1972–1975. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Edwards, J.G.; Riley, N.; Provance, D.W., Jr.; Karcher, R.; Li, X.D.; Davison, I.G.; Ikebe, M.; Mercer, J.A.; Kauer, J.A.; et al. Myosin Vb mobilizes recycling endosomes and AMPA receptors for postsynaptic plasticity. Cell 2008, 135, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Husebye, H.; Aune, M.H.; Stenvik, J.; Samstad, E.; Skjeldal, F.; Halaas, O.; Nilsen, N.J.; Stenmark, H.; Latz, E.; Lien, E.; et al. The Rab11a gtpase controls Toll-like receptor 4-induced activation of interferon regulatory factor-3 on phagosomes. Immunity 2010, 33, 583–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desclozeaux, M.; Venturato, J.; Wylie, F.G.; Kay, J.G.; Joseph, S.R.; le, H.T.; Stow, J.L. Active Rab11 and functional recycling endosome are required for e-cadherin trafficking and lumen formation during epithelial morphogenesis. Am. J. Physiol. 2008, 295, C545–C556. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, O.; Reinsch, S.; Urbe, S.; Zerial, M.; Parton, R.G. Rab11 regulates recycling through the pericentriolar recycling endosome. J. Cell Biol. 1996, 135, 913–924. [Google Scholar] [CrossRef] [PubMed]
- Hutagalung, A.H.; Novick, P.J. Role of Rab GTPases in membrane traffic and cell physiology. Physiol. Rev. 2011, 91, 119–149. [Google Scholar] [CrossRef] [PubMed]
- Jean, S.; Kiger, A.A. Coordination between Rab GTPase and phosphoinositide regulation and functions. Nat. Rev. 2012, 13, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Novick, P.; Field, C.; Schekman, R. Identification of 23 complementation groups required for post-translational events in the yeast secretory pathway. Cell 1980, 21, 205–215. [Google Scholar] [CrossRef]
- Salminen, A.; Novick, P.J. A Ras-like protein is required for a post-golgi event in yeast secretion. Cell 1987, 49, 527–538. [Google Scholar] [CrossRef]
- Chavrier, P.; Parton, R.G.; Hauri, H.P.; Simons, K.; Zerial, M. Localization of low molecular weight GTP binding proteins to exocytic and endocytic compartments. Cell 1990, 62, 317–329. [Google Scholar] [CrossRef]
- Pereira-Leal, J.B.; Seabra, M.C. Evolution of the Rab family of small GTP-binding proteins. J. Mol. Biol. 2001, 313, 889–901. [Google Scholar] [CrossRef] [PubMed]
- Alexandrov, K.; Horiuchi, H.; Steele-Mortimer, O.; Seabra, M.C.; Zerial, M. Rab escort protein-1 is a multifunctional protein that accompanies newly prenylated rab proteins to their target membranes. EMBO J. 1994, 13, 5262–5273. [Google Scholar] [PubMed]
- Andres, D.A.; Seabra, M.C.; Brown, M.S.; Armstrong, S.A.; Smeland, T.E.; Cremers, F.P.; Goldstein, J.L. cDNA cloning of component a of Rab geranylgeranyl transferase and demonstration of its role as a Rab escort protein. Cell 1993, 73, 1091–1099. [Google Scholar] [CrossRef]
- Armstrong, S.A.; Seabra, M.C.; Sudhof, T.C.; Goldstein, J.L.; Brown, M.S. cDNA cloning and expression of the alpha and beta subunits of Rat Rab geranylgeranyl transferase. J. Biol. Chem. 1993, 268, 12221–12229. [Google Scholar] [PubMed]
- Ullrich, O.; Stenmark, H.; Alexandrov, K.; Huber, L.A.; Kaibuchi, K.; Sasaki, T.; Takai, Y.; Zerial, M. Rab GDP dissociation inhibitor as a general regulator for the membrane association of Rab proteins. J. Biol. Chem. 1993, 268, 18143–18150. [Google Scholar] [PubMed]
- Collins, R.N. “Getting it on”—GDI displacement and small GTPase membrane recruitment. Mol. Cell 2003, 12, 1064–1066. [Google Scholar] [CrossRef]
- Dirac-Svejstrup, A.B.; Sumizawa, T.; Pfeffer, S.R. Identification of a GDI displacement factor that releases endosomal Rab GTPases from Rab-GDI. EMBO J. 1997, 16, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Christoforidis, S.; Miaczynska, M.; Ashman, K.; Wilm, M.; Zhao, L.; Yip, S.C.; Waterfield, M.D.; Backer, J.M.; Zerial, M. Phosphatidylinositol-3-OH kinases are Rab5 effectors. Nat. Cell Biol. 1999, 1, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Carlton, J.G.; Cullen, P.J. Coincidence detection in phosphoinositide signaling. Trends Cell Biol. 2005, 15, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Davies, B.A.; Carney, D.S.; Horazdovsky, B.F. Ubiquitin regulation of the Rab5 family GEF Vps9p. Methods Enzymol. 2005, 403, 561–583. [Google Scholar] [PubMed]
- Blumer, J.; Rey, J.; Dehmelt, L.; Mazel, T.; Wu, Y.W.; Bastiaens, P.; Goody, R.S.; Itzen, A. RabGEFs are a major determinant for specific Rab membrane targeting. J. Cell Biol. 2013, 200, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Gerondopoulos, A.; Langemeyer, L.; Liang, J.R.; Linford, A.; Barr, F.A. Bloc-3 mutated in hermansky-pudlak syndrome is a Rab32/38 guanine nucleotide exchange factor. Curr. Biology 2012, 22, 2135–2139. [Google Scholar] [CrossRef] [PubMed]
- Barr, F.; Lambright, D.G. Rab GEFs and GAPs. Curr. Opin. Cell Biol. 2010, 22, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Barr, F.A. Review series: Rab GTPases and membrane identity: Causal or inconsequential? J. Cell Biol. 2013, 202, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Mazelova, J.; Astuto-Gribble, L.; Inoue, H.; Tam, B.M.; Schonteich, E.; Prekeris, R.; Moritz, O.L.; Randazzo, P.A.; Deretic, D. Ciliary targeting motif VxPx directs assembly of a trafficking module through Arf4. EMBO J. 2009, 28, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, J.G.; Jackson, C.L. Arf family G proteins and their regulators: Roles in membrane transport, development and disease. Nat. Rev. Mol. Cell Biol. 2011, 12, 362–375. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Morita, Y.; Mazelova, J.; Deretic, D. The Arf GAP ASAP1 provides a platform to regulate Arf4- and Rab11-Rab8-mediated ciliary receptor targeting. EMBO J. 2012, 31, 4057–4071. [Google Scholar] [CrossRef] [PubMed]
- Schonteich, E.; Pilli, M.; Simon, G.C.; Matern, H.T.; Junutula, J.R.; Sentz, D.; Holmes, R.K.; Prekeris, R. Molecular characterization of Rab11-FIP3 binding to Arf GTPases. Eur. J. Cell Biol. 2007, 86, 417–431. [Google Scholar] [CrossRef] [PubMed]
- Inoue, H.; Ha, V.L.; Prekeris, R.; Randazzo, P.A. Arf GTPase-activating protein ASAP1 interacts with Rab11 effector FIP3 and regulates pericentrosomal localization of transferrin receptor-positive recycling endosome. Mol. Biol. Cell 2008, 19, 4224–4237. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, A.; Hirohama, M.; Harada, Y.; Osari, S.; Nagata, K. Influenza virus induces cholesterol-enriched endocytic recycling compartments for budozone formation via cell cycle-independent centrosome maturation. PLoS Pathog. 2015, 11, e1005284. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.; Newman, C.; Liu, F.; Segev, N. The TRAPP complex is a nucleotide exchanger for YPT1 and YPT31/32. Mol. Biol. Cell 2000, 11, 4403–4411. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Sacher, M.; Ferro-Novick, S. TRAPP stimulates guanine nucleotide exchange on YPT1p. J. Cell Biol. 2000, 151, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Gorvel, J.P.; Chavrier, P.; Zerial, M.; Gruenberg, J. Rab5 controls early endosome fusion in vitro. Cell 1991, 64, 915–925. [Google Scholar] [CrossRef]
- Goud, B.; Salminen, A.; Walworth, N.C.; Novick, P.J. A GTP-binding protein required for secretion rapidly associates with secretory vesicles and the plasma membrane in yeast. Cell 1988, 53, 753–768. [Google Scholar] [CrossRef]
- Lian, J.P.; Stone, S.; Jiang, Y.; Lyons, P.; Ferro-Novick, S. YPT1p implicated in v-SNARE activation. Nature 1994, 372, 698–701. [Google Scholar] [CrossRef] [PubMed]
- Segev, N. Mediation of the attachment or fusion step in vesicular transport by the GTP-binding YPT1 protein. Science 1991, 252, 1553–1556. [Google Scholar] [CrossRef] [PubMed]
- Sogaard, M.; Tani, K.; Ye, R.R.; Geromanos, S.; Tempst, P.; Kirchhausen, T.; Rothman, J.E.; Sollner, T. A Rab protein is required for the assembly of SNARE complexes in the docking of transport vesicles. Cell 1994, 78, 937–948. [Google Scholar] [CrossRef]
- Granger, E.; McNee, G.; Allan, V.; Woodman, P. The role of the cytoskeleton and molecular motors in endosomal dynamics. Semin. Cell Dev. Biol. 2014, 31, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, S.R. Rab GTPases: Specifying and deciphering organelle identity and function. Trends Cell Biol. 2001, 11, 487–491. [Google Scholar] [CrossRef]
- Zerial, M.; McBride, H. Rab proteins as membrane organizers. Nat. Rev. Mol. Cell Biol. 2001, 2, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Roth, D.; Walch-Solimena, C.; Novick, P. The exocyst is an effector for Sec4p, targeting secretory vesicles to sites of exocytosis. EMBO J. 1999, 18, 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, D.; Medkova, M.; Walch-Solimena, C.; Novick, P. YPT32 recruits the Sec4p guanine nucleotide exchange factor, Sec2p, to secretory vesicles; evidence for a Rab cascade in yeast. J. Cell Biol. 2002, 157, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Knodler, A.; Feng, S.; Zhang, J.; Zhang, X.; Das, A.; Peranen, J.; Guo, W. Coordination of Rab8 and Rab11 in primary ciliogenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 6346–6351. [Google Scholar] [CrossRef] [PubMed]
- Pusapati, G.V.; Luchetti, G.; Pfeffer, S.R. Ric1-Rgp1 complex is a guanine nucleotide exchange factor for the late golgi Rab6a GTPase and an effector of the medial golgi Rab33b GTPase. J. Biol. Chem. 2012, 287, 42129–42137. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Ohbayashi, N.; Maruta, Y.; Kanno, E.; Itoh, T.; Fukuda, M. Varp is a novel Rab32/38-binding protein that regulates TYRP1 trafficking in melanocytes. Mol. Biol. Cell 2009, 20, 2900–2908. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; He, X.; Fu, X.Y.; Chang, Z. Varp is a Rab21 guanine nucleotide exchange factor and regulates endosome dynamics. J. Cell Sci. 2006, 119, 1053–1062. [Google Scholar] [CrossRef] [PubMed]
- Nottingham, R.M.; Ganley, I.G.; Barr, F.A.; Lambright, D.G.; Pfeffer, S.R. RUTBC1 protein, a Rab9a effector that activates GTP hydrolysis by Rab32 and Rab33b proteins. J. Biol. Chem. 2011, 286, 33213–33222. [Google Scholar] [CrossRef] [PubMed]
- Nottingham, R.M.; Pusapati, G.V.; Ganley, I.G.; Barr, F.A.; Lambright, D.G.; Pfeffer, S.R. RUTBC2 protein, a Rab9a effector and GTPase-activating protein for Rab36. J. Biol. Chem. 2012, 287, 22740–22748. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Liu, X.; Arasaki, K.; McDonough, J.; Galan, J.E.; Roy, C.R. Modulation of Rab GTPase function by a protein phosphocholine transferase. Nature 2011, 477, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Muller, M.P.; Peters, H.; Blumer, J.; Blankenfeldt, W.; Goody, R.S.; Itzen, A. The legionella effector protein DrrA ampylates the membrane traffic regulator Rab1b. Science 2010, 329, 946–949. [Google Scholar] [CrossRef] [PubMed]
- Neunuebel, M.R.; Chen, Y.; Gaspar, A.H.; Backlund, P.S., Jr.; Yergey, A.; Machner, M.P. De-ampylation of the small GTPase Rab1 by the pathogen legionella pneumophila. Science 2011, 333, 453–456. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, A.; Macfarlane, J.; Shaw, J.M. Mitochondrial association, protein phosphorylation, and degradation regulate the availability of the active Rab GTPase YPT11 for mitochondrial inheritance. Mol. Biol. Cell 2013, 24, 1185–1195. [Google Scholar] [CrossRef] [PubMed]
- Heger, C.D.; Wrann, C.D.; Collins, R.N. Phosphorylation provides a negative mode of regulation for the yeast Rab GTPase Sec4p. PLoS ONE 2011, 6, e24332. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Soule, G.; Overmeyer, J.H.; Maltese, W.A. Tyrosine phosphorylation of the Rab24 GTPase in cultured mammalian cells. Biochem. Biophys. Res. Commun. 2003, 312, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Chua, C.E.; Tang, B.L. Role of Rab GTPases and their interacting proteins in mediating metabolic signalling and regulation. Cell. Mol. Life Sci. 2015, 72, 2289–2304. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Murate, M.; Fukuda, M.; Sato, S.B.; Ohta, A.; Kobayashi, T. Cholesterol controls lipid endocytosis through Rab11. Mol. Biol. Cell 2007, 18, 2667–2677. [Google Scholar] [CrossRef] [PubMed]
- Holtta-Vuori, M.; Tanhuanpaa, K.; Mobius, W.; Somerharju, P.; Ikonen, E. Modulation of cellular cholesterol transport and homeostasis by Rab11. Mol. Biol. Cell 2002, 13, 3107–3122. [Google Scholar] [CrossRef] [PubMed]
- Ganley, I.G.; Pfeffer, S.R. Cholesterol accumulation sequesters Rab9 and disrupts late endosome function in npc1-deficient cells. J. Biol. Chem. 2006, 281, 17890–17899. [Google Scholar] [CrossRef] [PubMed]
- Welz, T.; Wellbourne-Wood, J.; Kerkhoff, E. Orchestration of cell surface proteins by Rab11. Trends Cell Biol. 2014, 24, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, F.; Shirane, M.; Matsumoto, M.; Nakayama, K.I. Protrudin serves as an adaptor molecule that connects KIF5 and its cargoes in vesicular transport during process formation. Mol. Biol. Cell 2011, 22, 4602–4620. [Google Scholar] [CrossRef] [PubMed]
- Balla, T. Phosphoinositides: Tiny lipids with giant impact on cell regulation. Physiol. Rev. 2013, 93, 1019–1137. [Google Scholar] [CrossRef] [PubMed]
- Chishti, A.H.; Kim, A.C.; Marfatia, S.M.; Lutchman, M.; Hanspal, M.; Jindal, H.; Liu, S.C.; Low, P.S.; Rouleau, G.A.; Mohandas, N.; et al. The ferm domain: A unique module involved in the linkage of cytoplasmic proteins to the membrane. Trends Biochem. Sci. 1998, 23, 281–282. [Google Scholar] [CrossRef]
- Legendre-Guillemin, V.; Wasiak, S.; Hussain, N.K.; Angers, A.; McPherson, P.S. Enth/anth proteins and clathrin-mediated membrane budding. J. Cell Sci. 2004, 117, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Wishart, M.J.; Taylor, G.S.; Dixon, J.E. Phoxy lipids: Revealing PX domains as phosphoinositide binding modules. Cell 2001, 105, 817–820. [Google Scholar] [CrossRef]
- Harlan, J.E.; Hajduk, P.J.; Yoon, H.S.; Fesik, S.W. Pleckstrin homology domains bind to phosphatidylinositol-4,5-bisphosphate. Nature 1994, 371, 168–170. [Google Scholar] [CrossRef] [PubMed]
- Wandinger-Ness, A.; Zerial, M. Rab proteins and the compartmentalization of the endosomal system. Cold Spring Harb. Perspect. Biol. 2014, 6. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Zhang, Y.; Xu, P.; Xu, T.; Lou, X. Nanoscale landscape of phosphoinositides revealed by specific pleckstrin homology (ph) domains using single-molecule superresolution imaging in the plasma membrane. J. Biol. Chem. 2015, 290, 26978–26993. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Ueda, Y.; Takagi, T.; Umezawa, Y. Production of PtdInsP3 at endomembranes is triggered by receptor endocytosis. Nat. Cell Biol. 2003, 5, 1016–1022. [Google Scholar] [CrossRef] [PubMed]
- Watt, S.A.; Kimber, W.A.; Fleming, I.N.; Leslie, N.R.; Downes, C.P.; Lucocq, J.M. Detection of novel intracellular agonist responsive pools of phosphatidylinositol 3,4-bisphosphate using the TAPP1 pleckstrin homology domain in immunoelectron microscopy. Biochem. J. 2004, 377, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Egami, Y.; Taguchi, T.; Maekawa, M.; Arai, H.; Araki, N. Small GTPases and phosphoinositides in the regulatory mechanisms of macropinosome formation and maturation. Front. Physiol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Welliver, T.P.; Swanson, J.A. A growth factor signaling cascade confined to circular ruffles in macrophages. Biol. Open 2012, 1, 754–760. [Google Scholar] [CrossRef] [PubMed]
- Stenmark, H.; Aasland, R.; Toh, B.H.; D’Arrigo, A. Endosomal localization of the autoantigen EEA1 is mediated by a zinc-binding FYVE finger. J. Biol. Chem. 1996, 271, 24048–24054. [Google Scholar] [PubMed]
- Gaullier, J.M.; Simonsen, A.; D’Arrigo, A.; Bremnes, B.; Stenmark, H.; Aasland, R. FYVE fingers bind ptdins(3)p. Nature 1998, 394, 432–433. [Google Scholar] [CrossRef] [PubMed]
- Dove, S.K.; Cooke, F.T.; Douglas, M.R.; Sayers, L.G.; Parker, P.J.; Michell, R.H. Osmotic stress activates phosphatidylinositol-3,5-bisphosphate synthesis. Nature 1997, 390, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Ikonomov, O.C.; Sbrissa, D.; Foti, M.; Carpentier, J.L.; Shisheva, A. PIKFYVE controls fluid phase endocytosis but not recycling/degradation of endocytosed receptors or sorting of procathepsin d by regulating multivesicular body morphogenesis. Mol. Biol. Cell 2003, 14, 4581–4591. [Google Scholar] [CrossRef] [PubMed]
- Jin, N.; Chow, C.Y.; Liu, L.; Zolov, S.N.; Bronson, R.; Davisson, M.; Petersen, J.L.; Zhang, Y.; Park, S.; Duex, J.E.; et al. Vac14 nucleates a protein complex essential for the acute interconversion of PI3P and PI(3,5)P(2) in yeast and mouse. EMBO J. 2008, 27, 3221–3234. [Google Scholar] [CrossRef] [PubMed]
- Christoforidis, S.; McBride, H.M.; Burgoyne, R.D.; Zerial, M. The Rab5 effector EEA1 is a core component of endosome docking. Nature 1999, 397, 621–625. [Google Scholar] [PubMed]
- Simonsen, A.; Lippe, R.; Christoforidis, S.; Gaullier, J.M.; Brech, A.; Callaghan, J.; Toh, B.H.; Murphy, C.; Zerial, M.; Stenmark, H. EEA1 links PI(3)K function to Rab5 regulation of endosome fusion. Nature 1998, 394, 494–498. [Google Scholar] [PubMed]
- Franco, M.; Peters, P.J.; Boretto, J.; van Donselaar, E.; Neri, A.; D’Souza-Schorey, C.; Chavrier, P. EFA6, a sec7 domain-containing exchange factor for Arf6, coordinates membrane recycling and actin cytoskeleton organization. EMBO J. 1999, 18, 1480–1491. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.; Upender, S.; Hansen, S.H.; Casanova, J.E. ARNO is a guanine nucleotide exchange factor for ADP-ribosylation factor 6. J. Biol. Chem. 1998, 273, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, M.; Ando, T.; Terabayashi, T.; Okamoto, M.; Takei, M.; Nishioka, T.; Kaibuchi, K.; Matsunaga, K.; Ishizaki, R.; Izumi, T.; et al. PI3K regulates endocytosis after insulin secretion via signaling crosstalk between Arf6 and Rab27a. J. Cell Science 2015, 129, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Mayinger, P. Phosphoinositides and vesicular membrane traffic. Biochim. Biophys. Acta 2012, 1821, 1104–1113. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.W.; Hayashi, M.; Christoforidis, S.; Lacas-Gervais, S.; Hoepfner, S.; Wenk, M.R.; Modregger, J.; Uttenweiler-Joseph, S.; Wilm, M.; Nystuen, A.; et al. An enzymatic cascade of Rab5 effectors regulates phosphoinositide turnover in the endocytic pathway. J. Cell Biol. 2005, 170, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Cheng, K.W.; Mills, G.B. Rab25 in cancer: A brief update. Biochem. Soc. Trans. 2012, 40, 1404–1408. [Google Scholar] [CrossRef] [PubMed]
- Dozynkiewicz, M.A.; Jamieson, N.B.; Macpherson, I.; Grindlay, J.; van den Berghe, P.V.; von Thun, A.; Morton, J.P.; Gourley, C.; Timpson, P.; Nixon, C.; et al. Rab25 and clic3 collaborate to promote integrin recycling from late endosomes/lysosomes and drive cancer progression. Dev. Cell 2012, 22, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Deretic, D. Crosstalk of Arf and Rab GTPases en route to cilia. Small GTPases 2013, 4, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Pechlivanis, M.; Kuhlmann, J. Hydrophobic modifications of Ras proteins by isoprenoid groups and fatty acids--more than just membrane anchoring. Biochim. Biophys. Acta 2006, 1764, 1914–1931. [Google Scholar] [CrossRef] [PubMed]
- Massignan, T.; Biasini, E.; Lauranzano, E.; Veglianese, P.; Pignataro, M.; Fioriti, L.; Harris, D.A.; Salmona, M.; Chiesa, R.; Bonetto, V. Mutant prion protein expression is associated with an alteration of the Rab GDP dissociation inhibitor alpha (GDI)/Rab11 pathway. Mol. Cell. Proteom. 2010, 9, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Burke, J.E.; Inglis, A.J.; Perisic, O.; Masson, G.R.; McLaughlin, S.H.; Rutaganira, F.; Shokat, K.M.; Williams, R.L. Structures of PI4KIIIbeta complexes show simultaneous recruitment of Rab11 and its effectors. Science 2014, 344, 1035–1038. [Google Scholar] [CrossRef] [PubMed]
- De Graaf, P.; Zwart, W.T.; van Dijken, R.A.; Deneka, M.; Schulz, T.K.; Geijsen, N.; Coffer, P.J.; Gadella, B.M.; Verkleij, A.J.; van der Sluijs, P.; et al. Phosphatidylinositol 4-kinasebeta is critical for functional association of Rab11 with the golgi complex. Mol. Biol. Cell 2004, 15, 2038–2047. [Google Scholar] [CrossRef] [PubMed]
- Fields, I.C.; King, S.M.; Shteyn, E.; Kang, R.S.; Folsch, H. Phosphatidylinositol 3,4,5-trisphosphate localization in recycling endosomes is necessary for ap-1b-dependent sorting in polarized epithelial cells. Mol. Biol. Cell 2010, 21, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Thapa, N.; Sun, Y.; Schramp, M.; Choi, S.; Ling, K.; Anderson, R.A. Phosphoinositide signaling regulates the exocyst complex and polarized integrin trafficking in directionally migrating cells. Dev. Cell 2012, 22, 116–130. [Google Scholar] [CrossRef] [PubMed]
- Xiong, B.; Bayat, V.; Jaiswal, M.; Zhang, K.; Sandoval, H.; Charng, W.L.; Li, T.; David, G.; Duraine, L.; Lin, Y.Q.; et al. Crag is a GEF for Rab11 required for rhodopsin trafficking and maintenance of adult photoreceptor cells. PLoS Biol. 2012, 10, e1001438. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, A.; Sato, M.; Sato, K.; Gengyo-Ando, K.; Yorimitsu, T.; Nakai, J.; Hara, T. Rei-1 is a guanine nucleotide exchange factor regulating Rab-11 localization and function in C. elegans embryos. Dev. Cell 2015, 35, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Sapp, E.; Valencia, A.; Kegel, K.B.; Qin, Z.H.; Alexander, J.; Masso, N.; Reeves, P.; Ritch, J.J.; Zeitlin, S.; et al. A function of huntingtin in guanine nucleotide exchange on Rab11. Neuroreport 2008, 19, 1643–1647. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Standley, C.; Sapp, E.; Valencia, A.; Qin, Z.H.; Kegel, K.B.; Yoder, J.; Comer-Tierney, L.A.; Esteves, M.; Chase, K.; et al. Mutant huntingtin impairs vesicle formation from recycling endosomes by interfering with Rab11 activity. Mol. Cell. Biol. 2009, 29, 6106–6116. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.H.; Lapierre, L.A.; Goldenring, J.R.; Sai, J.; Richmond, A. Rab11-family interacting protein 2 and myosin Vb are required for CXCR2 recycling and receptor-mediated chemotaxis. Mol. Biol. Cell 2004, 15, 2456–2469. [Google Scholar] [CrossRef] [PubMed]
- Lapierre, L.A.; Kumar, R.; Hales, C.M.; Navarre, J.; Bhartur, S.G.; Burnette, J.O.; Provance, D.W., Jr.; Mercer, J.A.; Bahler, M.; Goldenring, J.R. Myosin Vb is associated with plasma membrane recycling systems. Mol. Biol. Cell 2001, 12, 1843–1857. [Google Scholar] [CrossRef] [PubMed]
- Provance, D.W., Jr.; Addison, E.J.; Wood, P.R.; Chen, D.Z.; Silan, C.M.; Mercer, J.A. Myosin-Vb functions as a dynamic tether for peripheral endocytic compartments during transferrin trafficking. BMC Cell Biol. 2008, 9. [Google Scholar] [CrossRef] [PubMed]
- Desnos, C.; Huet, S.; Darchen, F. “Should i stay or should i go?”: Myosin V function in organelle trafficking. Biol. Cell 2007, 99, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Roland, J.T.; Bryant, D.M.; Datta, A.; Itzen, A.; Mostov, K.E.; Goldenring, J.R. Rab GTPase-Myo5b complexes control membrane recycling and epithelial polarization. Proc. Natl. Acad. Sci. USA 2011, 108, 2789–2794. [Google Scholar] [CrossRef] [PubMed]
- Tarafder, A.K.; Bolasco, G.; Correia, M.S.; Pereira, F.J.; Iannone, L.; Hume, A.N.; Kirkpatrick, N.; Picardo, M.; Torrisi, M.R.; Rodrigues, I.P.; et al. Rab11b mediates melanin transfer between donor melanocytes and acceptor keratinocytes via coupled exo/endocytosis. J. Investig. Dermatol. 2014, 134, 1056–1066. [Google Scholar] [CrossRef] [PubMed]
- Aikawa, Y.; Lynch, K.L.; Boswell, K.L.; Martin, T.F. A second SNARE role for exocytic SNAP25 in endosome fusion. Mol. Biol. Cell 2006, 17, 2113–2124. [Google Scholar] [CrossRef] [PubMed]
- Laflamme, C.; Assaker, G.; Ramel, D.; Dorn, J.F.; She, D.; Maddox, P.S.; Emery, G. Evi5 promotes collective cell migration through its Rab-gap activity. J. Cell Biol. 2012, 198, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Schonteich, E.; Wilson, G.M.; Burden, J.; Hopkins, C.R.; Anderson, K.; Goldenring, J.R.; Prekeris, R. The Rip11/Rab11-FIP5 and kinesin II complex regulates endocytic protein recycling. J. Cell Sci. 2008, 121, 3824–3833. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Mehta, S.Q.; Pichaud, F.; Bellen, H.J.; Quiocho, F.A. Sec15 interacts with Rab11 via a novel domain and affects Rab11 localization in vivo. Nat. Struct. Mol. Biol. 2005, 12, 879–885. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.M.; Ellis, S.; Sriratana, A.; Mitchell, C.A.; Rowe, T. Sec15 is an effector for the Rab11 GTPase in mammalian cells. J. Biol. Chem. 2004, 279, 43027–43034. [Google Scholar] [CrossRef] [PubMed]
- Fielding, A.B.; Schonteich, E.; Matheson, J.; Wilson, G.; Yu, X.; Hickson, G.R.; Srivastava, S.; Baldwin, S.A.; Prekeris, R.; Gould, G.W. Rab11-FIP3 and FIP4 interact with Arf6 and the exocyst to control membrane traffic in cytokinesis. EMBO J. 2005, 24, 3389–3399. [Google Scholar] [CrossRef] [PubMed]
- Mehta, S.Q.; Hiesinger, P.R.; Beronja, S.; Zhai, R.G.; Schulze, K.L.; Verstreken, P.; Cao, Y.; Zhou, Y.; Tepass, U.; Crair, M.C.; et al. Mutations in drosophila Sec15 reveal a function in neuronal targeting for a subset of exocyst components. Neuron 2005, 46, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Grant, B.D.; Harada, A.; Sato, K. Rab11 is required for synchronous secretion of chondroitin proteoglycans after fertilization in caenorhabditis elegans. J. Cell Sci. 2008, 121, 3177–3186. [Google Scholar] [CrossRef] [PubMed]
- Gallo, L.I.; Liao, Y.; Ruiz, W.G.; Clayton, D.R.; Li, M.; Liu, Y.J.; Jiang, Y.; Fukuda, M.; Apodaca, G.; Yin, X.M. TBC1D9B functions as a GTPase-activating protein for Rab11a in polarized MDCK cells. Mol. Biol. Cell 2014, 25, 3779–3797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delevoye, C.; Miserey-Lenkei, S.; Montagnac, G.; Gilles-Marsens, F.; Paul-Gilloteaux, P.; Giordano, F.; Waharte, F.; Marks, M.S.; Goud, B.; Raposo, G. Recycling endosome tubule morphogenesis from sorting endosomes requires the kinesin motor KIF13a. Cell Rep. 2014, 6, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Boswell, K.L.; James, D.J.; Esquibel, J.M.; Bruinsma, S.; Shirakawa, R.; Horiuchi, H.; Martin, T.F. Munc13–4 reconstitutes calcium-dependent SNARE-mediated membrane fusion. J. Cell Biol. 2012, 197, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Marshall, M.R.; Pattu, V.; Halimani, M.; Maier-Peuschel, M.; Muller, M.L.; Becherer, U.; Hong, W.; Hoth, M.; Tschernig, T.; Bryceson, Y.T.; et al. Vamp8-dependent fusion of recycling endosomes with the plasma membrane facilitates t lymphocyte cytotoxicity. J. Cell Biol. 2015, 210, 135–151. [Google Scholar] [CrossRef] [PubMed]
- Horgan, C.P.; Hanscom, S.R.; Jolly, R.S.; Futter, C.E.; McCaffrey, M.W. Rab11-FIP3 links the Rab11 GTPase and cytoplasmic dynein to mediate transport to the endosomal-recycling compartment. J. Cell Sci. 2010, 123, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Horgan, C.P.; Hanscom, S.R.; McCaffrey, M.W. Grab is a binding partner for the Rab11a and Rab11b GTPases. Biochem. Biophys. Res. Commun. 2013, 441, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Miserey-Lenkei, S.; Waharte, F.; Boulet, A.; Cuif, M.H.; Tenza, D.; El Marjou, A.; Raposo, G.; Salamero, J.; Heliot, L.; Goud, B.; et al. Rab6-interacting protein 1 links Rab6 and Rab11 function. Traffic 2007, 8, 1385–1403. [Google Scholar] [CrossRef] [PubMed]
- Hales, C.M.; Griner, R.; Hobdy-Henderson, K.C.; Dorn, M.C.; Hardy, D.; Kumar, R.; Navarre, J.; Chan, E.K.; Lapierre, L.A.; Goldenring, J.R. Identification and characterization of a family of Rab11-interacting proteins. J. Biol. Chem. 2001, 276, 39067–39075. [Google Scholar] [CrossRef] [PubMed]
- Horgan, C.P.; McCaffrey, M.W. The dynamic Rab11-FIPs. Biochem. Soc. Trans. 2009, 37, 1032–1036. [Google Scholar] [CrossRef] [PubMed]
- Baetz, N.W.; Goldenring, J.R. Rab11-family interacting proteins define spatially and temporally distinct regions within the dynamic Rab11a-dependent recycling system. Mol. Biol. Cell 2013, 24, 643–658. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, A.J.; Hendrick, A.G.; Cantalupo, G.; Senic-Matuglia, F.; Goud, B.; Bucci, C.; McCaffrey, M.W. Rab coupling protein (RCP), a novel Rab4 and Rab11 effector protein. J. Biol. Chem. 2002, 277, 12190–12199. [Google Scholar] [CrossRef] [PubMed]
- Prekeris, R.; Klumperman, J.; Scheller, R.H. A Rab11/rip11 protein complex regulates apical membrane trafficking via recycling endosomes. Mol. Cell 2000, 6, 1437–1448. [Google Scholar] [CrossRef]
- Wallace, D.M.; Lindsay, A.J.; Hendrick, A.G.; McCaffrey, M.W. Rab11-FIP4 interacts with Rab11 in a GTP-dependent manner and its overexpression condenses the Rab11 positive compartment in HeLa cells. Biochem. Biophys. Res. Commun. 2002, 299, 770–779. [Google Scholar] [CrossRef]
- Schafer, J.C.; Baetz, N.W.; Lapierre, L.A.; McRae, R.E.; Roland, J.T.; Goldenring, J.R. Rab11-FIP2 interaction with Myo5b regulates movement of Rab11a-containing recycling vesicles. Traffic 2014, 15, 292–308. [Google Scholar] [CrossRef] [PubMed]
- Ducharme, N.A.; Hales, C.M.; Lapierre, L.A.; Ham, A.J.; Oztan, A.; Apodaca, G.; Goldenring, J.R. MARK2/EMK1/Par-1Balpha phosphorylation of Rab11-family interacting protein 2 is necessary for the timely establishment of polarity in madin-darby canine kidney cells. Mol. Biol. Cell 2006, 17, 3625–3637. [Google Scholar] [CrossRef] [PubMed]
- Collins, L.L.; Simon, G.; Matheson, J.; Wu, C.; Miller, M.C.; Otani, T.; Yu, X.; Hayashi, S.; Prekeris, R.; Gould, G.W. Rab11-FIP3 is a cell cycle-regulated phosphoprotein. BMC Cell Biol. 2012, 13. [Google Scholar] [CrossRef] [PubMed]
- Bridgewater, R.E.; Norman, J.C.; Caswell, P.T. Integrin trafficking at a glance. J. Cell Sci. 2012, 125, 3695–3701. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Setou, M.; Seog, D.; Ogasawara, K.; Dohmae, N.; Takio, K.; Hirokawa, N. A novel motor, KIF13a, transports mannose-6-phosphate receptor to plasma membrane through direct interaction with ap-1 complex. Cell 2000, 103, 569–581. [Google Scholar] [CrossRef]
- Bombardier, J.P.; Munson, M. Three steps forward, two steps back: Mechanistic insights into the assembly and disassembly of the SNARE complex. Curr. Opin. Chem. Biol. 2015, 29, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.L.; He, J.; Ramadass, M.; Pestonjamasp, K.; Kiosses, W.; Zhang, J.; Catz, S.D. Munc13-4 is a Rab11-binding protein that regulates Rab11-positive vesicle trafficking and docking at the plasma membrane. J. Biol. Chem. 2015. [Google Scholar] [CrossRef] [PubMed]
- Dabbeekeh, J.T.; Faitar, S.L.; Dufresne, C.P.; Cowell, J.K. The EVI5 TBC domain provides the GTPase-activating protein motif for Rab11. Oncogene 2007, 26, 2804–2808. [Google Scholar] [CrossRef] [PubMed]
- Westlake, C.J.; Junutula, J.R.; Simon, G.C.; Pilli, M.; Prekeris, R.; Scheller, R.H.; Jackson, P.K.; Eldridge, A.G. Identification of Rab11 as a small GTPase binding protein for the EVI5 oncogene. Proc. Natl. Acad. Sci. USA 2007, 104, 1236–1241. [Google Scholar] [CrossRef] [PubMed]
- Stone, R.; Hayashi, T.; Bajimaya, S.; Hodges, E.; Takimoto, T. Critical role of Rab11a-mediated recycling endosomes in the assembly of typeI parainfluenza viruses. Virology 2015, 487, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Amorim, M.J.; Bruce, E.A.; Read, E.K.; Foeglein, A.; Mahen, R.; Stuart, A.D.; Digard, P. A Rab11- and microtubule-dependent mechanism for cytoplasmic transport of influenza a virus viral RNA. J. Virol. 2011, 85, 4143–4156. [Google Scholar] [CrossRef] [PubMed]
- Landry, M.C.; Sicotte, A.; Champagne, C.; Lavoie, J.N. Regulation of cell death by recycling endosomes and golgi membrane dynamics via a pathway involving Src-family kinases, Cdc42 and Rab11a. Mol. Biol. Cell 2009, 20, 4091–4106. [Google Scholar] [CrossRef] [PubMed]
- Landry, M.C.; Champagne, C.; Boulanger, M.C.; Jette, A.; Fuchs, M.; Dziengelewski, C.; Lavoie, J.N. A functional interplay between the small GTPase Rab11a and mitochondria-shaping proteins regulates mitochondrial positioning and polarization of the actin cytoskeleton downstream of Src family kinases. J. Biol. Chem. 2014, 289, 2230–2249. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, A.; Das, S.R.; Jameel, S.; George, A.; Bal, V.; Mayor, S.; Rath, S. Hiv-1 nef induces a Rab11-dependent routing of endocytosed immune costimulatory proteins CD80 and CD86 to the golgi. Traffic 2008, 9, 1925–1935. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, J.C.; Chu, L.W.; Lo, Y.T.; Lee, S.P.; Chen, T.J.; Huang, C.Y.; Ping, Y.H.; Chang, W. Intracellular transport of vaccinia virus in HeLa cells requires WASH-VPEF/FAM21-Retromer complexes and recycling molecules Rab11 and Rab22. J. Virol. 2015, 89, 8365–8382. [Google Scholar] [CrossRef] [PubMed]
- Mannova, P.; Forstova, J. Mouse polyomavirus utilizes recycling endosomes for a traffic pathway independent of COPI vesicle transport. J. Virol. 2003, 77, 1672–1681. [Google Scholar] [CrossRef] [PubMed]
- Suikkanen, S.; Saajarvi, K.; Hirsimaki, J.; Valilehto, O.; Reunanen, H.; Vihinen-Ranta, M.; Vuento, M. Role of recycling endosomes and lysosomes in dynein-dependent entry of canine parvovirus. J. Virol. 2002, 76, 4401–4411. [Google Scholar] [CrossRef] [PubMed]
- Acosta, E.G.; Castilla, V.; Damonte, E.B. Differential requirements in endocytic trafficking for penetration of dengue virus. PLoS ONE 2012, 7, e44835. [Google Scholar] [CrossRef] [PubMed]
- Johns, H.L.; Berryman, S.; Monaghan, P.; Belsham, G.J.; Jackson, T. A dominant-negative mutant of Rab5 inhibits infection of cells by foot-and-mouth disease virus: Implications for virus entry. J. Virol. 2009, 83, 6247–6256. [Google Scholar] [CrossRef] [PubMed]
- Greene, W.; Gao, S.J. Actin dynamics regulate multiple endosomal steps during Kaposi’s sarcoma-associated herpes virus entry and trafficking in endothelial cells. PLoS Pathog. 2009, 5, e1000512. [Google Scholar] [CrossRef] [PubMed]
- Zila, V.; Difato, F.; Klimova, L.; Huerfano, S.; Forstova, J. Involvement of microtubular network and its motors in productive endocytic trafficking of mouse polyomavirus. PLoS ONE 2014, 9, e96922. [Google Scholar] [CrossRef] [PubMed]
- Chambers, R.; Takimoto, T. Trafficking of Sendai virus nucleocapsids is mediated by intracellular vesicles. PLoS ONE 2010, 5, e10994. [Google Scholar] [CrossRef] [PubMed]
- Katoh, H.; Nakatsu, Y.; Kubota, T.; Sakata, M.; Takeda, M.; Kidokoro, M. Mumps virus is released from the apical surface of polarized epithelial cells, and the release is facilitated by a Rab11-mediated transport system. J. Virol. 2015, 89, 12026–12034. [Google Scholar] [CrossRef] [PubMed]
- Nakatsu, Y.; Ma, X.; Seki, F.; Suzuki, T.; Iwasaki, M.; Yanagi, Y.; Komase, K.; Takeda, M. Intracellular transport of the measles virus ribonucleoprotein complex is mediated by Rab11a-positive recycling endosomes and drives virus release from the apical membrane of polarized epithelial cells. J. Virol. 2013, 87, 4683–4693. [Google Scholar] [CrossRef] [PubMed]
- Momose, F.; Sekimoto, T.; Ohkura, T.; Jo, S.; Kawaguchi, A.; Nagata, K.; Morikawa, Y. Apical transport of influenza a virus ribonucleoprotein requires Rab11-positive recycling endosome. PLoS ONE 2011, 6, e21123. [Google Scholar] [CrossRef] [PubMed]
- Bruce, E.A.; Digard, P.; Stuart, A.D. The Rab11 pathway is required for influenza a virus budding and filament formation. J. Virol. 2010, 84, 5848–5859. [Google Scholar] [CrossRef] [PubMed]
- Avilov, S.V.; Moisy, D.; Munier, S.; Schraidt, O.; Naffakh, N.; Cusack, S. Replication-competent influenza A virus that encodes a split-green fluorescent protein-tagged PB2 polymerase subunit allows live-cell imaging of the virus life cycle. J. Virol. 2012, 86, 1433–1448. [Google Scholar] [CrossRef] [PubMed]
- Eisfeld, A.J.; Kawakami, E.; Watanabe, T.; Neumann, G.; Kawaoka, Y. Rab11a is essential for transport of the influenza virus genome to the plasma membrane. J. Virol. 2011, 85, 6117–6126. [Google Scholar] [CrossRef] [PubMed]
- Rowe, R.K.; Suszko, J.W.; Pekosz, A. Roles for the recycling endosome, Rab8, and Rab11 in hantavirus release from epithelial cells. Virology 2008, 382, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Chou, Y.Y.; Heaton, N.S.; Gao, Q.; Palese, P.; Singer, R.H.; Lionnet, T. Colocalization of different influenza viral RNA segments in the cytoplasm before viral budding as shown by single-molecule sensitivity fish analysis. PLoS Pathog. 2013, 9, e1003358. [Google Scholar] [CrossRef]
- Lakdawala, S.S.; Wu, Y.; Wawrzusin, P.; Kabat, J.; Broadbent, A.J.; Lamirande, E.W.; Fodor, E.; Altan-Bonnet, N.; Shroff, H.; Subbarao, K. Influenza A virus assembly intermediates fuse in the cytoplasm. PLoS Pathog. 2014, 10, e1003971. [Google Scholar] [CrossRef] [PubMed]
- Hollinshead, M.; Johns, H.L.; Sayers, C.L.; Gonzalez-Lopez, C.; Smith, G.L.; Elliott, G. Endocytic tubules regulated by Rab GTPases 5 and 11 are used for envelopment of herpes simplex virus. EMBO J. 2012, 31, 4204–4220. [Google Scholar] [CrossRef] [PubMed]
- Diederich, S.; Sauerhering, L.; Weis, M.; Altmeppen, H.; Schaschke, N.; Reinheckel, T.; Erbar, S.; Maisner, A. Activation of the Nipah virus fusion protein in MDCK cells is mediated by cathepsin b within the endosome-recycling compartment. J. Virol. 2012, 86, 3736–3745. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.; Williams, J.A.; Chu, H.; Chen, X.; Wang, J.J.; Ding, L.; Akhirome, E.; Wen, X.; Lapierre, L.A.; Goldenring, J.R.; et al. Rab11-FIP1c and Rab14 direct plasma membrane sorting and particle incorporation of the HIV-1 envelope glycoprotein complex. PLoS Pathog. 2013, 9, e1003278. [Google Scholar] [CrossRef] [PubMed]
- Varthakavi, V.; Smith, R.M.; Martin, K.L.; Derdowski, A.; Lapierre, L.A.; Goldenring, J.R.; Spearman, P. The pericentriolar recycling endosome plays a key role in Vpu-mediated enhancement of HIV-1 particle release. Traffic 2006, 7, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Brock, S.C.; Goldenring, J.R.; Crowe, J.E., Jr. Apical recycling systems regulate directional budding of respiratory syncytial virus from polarized epithelial cells. Proc. Natl. Acad. Sci. USA 2003, 100, 15143–15148. [Google Scholar] [CrossRef] [PubMed]
- Utley, T.J.; Ducharme, N.A.; Varthakavi, V.; Shepherd, B.E.; Santangelo, P.J.; Lindquist, M.E.; Goldenring, J.R.; Crowe, J.E., Jr. Respiratory syncytial virus uses a Vps4-independent budding mechanism controlled by Rab11-FIP2. Proc. Natl. Acad. Sci. USA 2008, 105, 10209–10214. [Google Scholar] [CrossRef] [PubMed]
- Sfakianos, J.N.; Hunter, E. M-PMV capsid transport is mediated by env/gag interactions at the pericentriolar recycling endosome. Traffic 2003, 4, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Arnaud, F.; Murcia, P.R.; Palmarini, M. Mechanisms of late restriction induced by an endogenous retrovirus. J. Virol. 2007, 81, 11441–11451. [Google Scholar] [CrossRef] [PubMed]
- Kinlock, B.L.; Wang, Y.; Turner, T.M.; Wang, C.; Liu, B. Transcytosis of HIV-1 through vaginal epithelial cells is dependent on trafficking to the endocytic recycling pathway. PLoS ONE 2014, 9, e96760. [Google Scholar] [CrossRef] [PubMed]
- Coller, K.E.; Heaton, N.S.; Berger, K.L.; Cooper, J.D.; Saunders, J.L.; Randall, G. Molecular determinants and dynamics of hepatitis c virus secretion. PLoS Pathog. 2012, 8, e1002466. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.E.; Clark, J.; Grznarova, P.; Wen, X.; LaCasse, R.; Ruml, T.; Spearman, P.; Hunter, E. Direct evidence for intracellular anterograde co-transport of M-PMV Gag and Env on microtubules. Virology 2014, 449, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, A.; Matsumoto, K.; Nagata, K. Yb-1 functions as a porter to lead influenza virus ribonucleoprotein complexes to microtubules. J. Virol. 2012, 86, 11086–11095. [Google Scholar] [CrossRef] [PubMed]
- Lachambre, S.; Chopard, C.; Beaumelle, B. Preliminary characterisation of nanotubes connecting T-cells and their use by HIV-1. Biol. Cell 2014, 106, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Wilcke, M.; Johannes, L.; Galli, T.; Mayau, V.; Goud, B.; Salamero, J. Rab11 regulates the compartmentalization of early endosomes required for efficient transport from early endosomes to the trans-golgi network. J. Cell Biol. 2000, 151, 1207–1220. [Google Scholar] [CrossRef] [PubMed]






| Recruitment/Activation | Transport | Tethering | Fusion | Inactivation/Recycling |
|---|---|---|---|---|
| Crag [124] | Myosin Vb (FIP2) [128,129,130,131,132] | Rab11b [133] | SNAP25 [134] | Evi5 [135] |
| REI-1 [125] | KIF5a and KIF3 (Kinesin II) (FIP5) [136] | Sec15 (exocyst) [137,138,139,140] | SYN4 [141] | TBC1D9B [142] |
| PIP4KIII [120,121] | KIF13A [143] | Munc 13-4 [144] | VAMP8 [145] | Cholesterol [87,88,89] |
| DLIC1/2 * (FIP3) [146] |
| Family | Genome | Virus | Host | Cell Type | Pathology | Evidence for Usage of ERC | References |
|---|---|---|---|---|---|---|---|
| Poxviridae | ds DNA | Vaccinia virus (VV) | human and a wide range of animals | Keratinocytes, dermal fibroblasts and microvascular endothelial cells; tropism for tumour cells | Similar to smallpox, but milder; causes rash, fever, headache and body aches | Virus uncoating into the cytoplasm occurs in Rab11- and Rab22-positive recycling endosomes | [169] |
| Polyomaviridae | ds DNA | Mouse polyomavirus (mPyV) | mouse | Epithelial and mesenchymal cells of the respiratory system, kidneys and brain | Induce carcinogenesis | Partial co-localization of capsids with the Rab11-recycling endosomes upon entry | [170] |
| Herpesviridae | ds DNA | Kaposi’s sarcoma-associated herpesvirus (KSHV) | human | B lymphocytes, endothelial, and epithelial cells | Kaposi’s sarcoma, peripheral effusion lymphoma or multicentric Castleman’s disease | Co-localization of viral particles with Rab11-endosomes early after entry | [174] |
| Parvovirus | ss DNA | Canine parvovirus (CPV) | dog | Rapidly dividing lymphocytes and epithelium of the small intestine | Lethargy, vomiting, fever, and diarrhea | Co-localization of capsids with transferrin in a perinuclear area soon after internalization | [171] |
| Flaviviridae | + ss RNA | Dengue virus (DENV) | human | Keratinocytes, Langerhans cells, monocytes and macrophages | Mild fever; complications like dengue hemorrhagic fever may occur (fever, damage to lymphatic and circulatory system) | The recycling pathway (Rab22 and Rab11) is used for viral fusion and uncoating steps | [172] |
| Picornaviridae | + ss RNA | Foot-and-mouth disease virus (FMDV) | cattle, pig, sheep and goat | Epithelia | Fever, followed by blisters in the mouth and feet that may rupture and lead to lameness | Capacity to infect cells dependent on active Rab11 | [173] |
| Family | Genome | Virus | Host | Cell Type | Pathology | Evidence for Usage of ERC | References |
|---|---|---|---|---|---|---|---|
| Paramyxoviridae | − ss RNA | Respiratory syncytial virus (RSV) | human | lung epithelia | upper and lower respiratory tract infections (such as colds, bronchiolitis and pneumonia) | Rab11-FIP1 and -FIP2 are required for virus replication and budding at the apical plasma membrane | [190,191] |
| Sendai virus (SeV) | rodent | lung epithelia | respiratory tract infection | Rab11a and transferrin co-localize with vRNPs in large intracellular aggregates | [164,176] | ||
| Human parainfluenza virus type 1 (hPIV1) | human | lung epithelia | respiratory tract infection; croup or pneumonia | Rab11a co-localizes with vRNPs in large intracellular aggregates | [164] | ||
| Measles virus (MV) | human | lung tissue macrophages and dendritic cells, lymph node B and T cells, epithelial cells of the liver, spleen or even brain | immune system infection and respiratory tract infection | RNPs co-traffic with Rab11a endosomes, accumulate at the apical recycling compartment and beneath the apical membrane | [178] | ||
| Mumps virus (MuV) | human | systemic epithelia | swelling of parotid glands, accompanied by severe complications such as orchitis, aseptic meningitis, pancreatitis and deafness | Rab11 recycling endosomes transport vRNPs to the apical membrane | [177] | ||
| Nipah virus (NiV) | human and animals | systemic epithelia and endothelia | severe encephalitic and respiratory diseases | Early (Rab4) and late (Rab11) recycling endosomes are likely involved in the cleavage and activation of F protein | [187] | ||
| Orthomyxoviridae | − ss RNA (8 segments) | Influenza A virus (IAV) | human, pig, horse, bird | lung epithelia | respiratory tract infection | Rab11 is required for vRNP trafficking and virion budding at the surface; interaction between vRNPs and Rab11 is mediated by viral PB2; Rab11-FIPs influence vRNPs localization at the recycling endosome and later at the surface; Rab11-FIP3 required for filamentous virion formation | [165,179,180,181,182] |
| Bunyaviridae | − ss RNA (3 segments) | Andes virus (ANDV) | humans and rodents | endothelia | hantavirus cardiopulmonary syndrome (HPS) | Rab11 is required for viral production and co-localizes with nucleocapsid protein | [183] |
| Retroviridae | + ss RNA (2 copies) | Mason-Pfizer monkey virus (M-PMV) | macaque | epithelial | fatal immunodeficiency syndrome | Recycling endosome (Rab11) required for the Env-dependent export of Gag-assembled capsids towards the surface | [192] |
| Jaagsiekte sheep retrovirus (JRSV) | sheep and goat | lung epithelia, lymphocytes and myeloid cells | ovine pulmonary adenocarcinoma (OPA) | Recycling endosomes (Rab11) co-localize with Gag protein in the pericentriolar region and are involved in virion cell exit | [193] | ||
| Human immunodeficiency virus 1 (HIV-1) | human | CD4+ T cells, macrophages, microglial cells and dendritic cells | acquired immunodeficiency syndrome (AIDS) | Vpu co-localizes with the pericentriolar recycling endosome (transferrin and Rab11); Rab11 is required for the Vpu-enhancement of viral particle release | [189] | ||
| Human immunodeficiency virus 1 (HIV-1) | human | CD4+ T cells, macrophages, microglial cells and dendritic cells | acquired immunodeficiency syndrome (AIDS) | The endocytic recycling compartment (transferrin) is used for the transcytosis of HIV-1 in vaginal epithelial cells | [194] | ||
| Human immunodeficiency virus 1 (HIV-1) | human | CD4+ T cells, macrophages, microglial cells and dendritic cells | acquired immunodeficiency syndrome (AIDS) | FIP1C binds to Rab14 and redistributes out of the endosomal recycling complex for Env trafficking and incorporation onto virions towards the surface | [184] | ||
| Flaviviridae | + ss RNA | Hepatitis C virus (HCV) | human | hepatocytes (epithelial) | liver damage, cancer or chirrosis | Recycling endosome (Rab11a) is involved in egress of viral cores from the Golgi to cell periphery | [195] |
| Herpesviridae | ds DNA | Herpes simplex virus 1 (HSV-1) | human | epithelial and neuronal cells | forms blisters on or around affected areas - usually the mouth, genitals, or rectum. The blisters break, leaving tender sores | Recycling endosomes (Rab11) are likely involved in internalization of viral glycoproteins from the surface to be included in the capsids | [186] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vale-Costa, S.; Amorim, M.J. Recycling Endosomes and Viral Infection. Viruses 2016, 8, 64. https://doi.org/10.3390/v8030064
Vale-Costa S, Amorim MJ. Recycling Endosomes and Viral Infection. Viruses. 2016; 8(3):64. https://doi.org/10.3390/v8030064
Chicago/Turabian StyleVale-Costa, Sílvia, and Maria João Amorim. 2016. "Recycling Endosomes and Viral Infection" Viruses 8, no. 3: 64. https://doi.org/10.3390/v8030064
APA StyleVale-Costa, S., & Amorim, M. J. (2016). Recycling Endosomes and Viral Infection. Viruses, 8(3), 64. https://doi.org/10.3390/v8030064