
Biological Characteristics of Experimental Genotype Mixtures of Cydia Pomonella Granulovirus (CpGV): Ability to Control Susceptible and Resistant Pest Populations



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Viruses
2.3. Mixed Viral Populations
2.4. Amplification of the Different Viral Mixed Populations
2.5. Bioassays
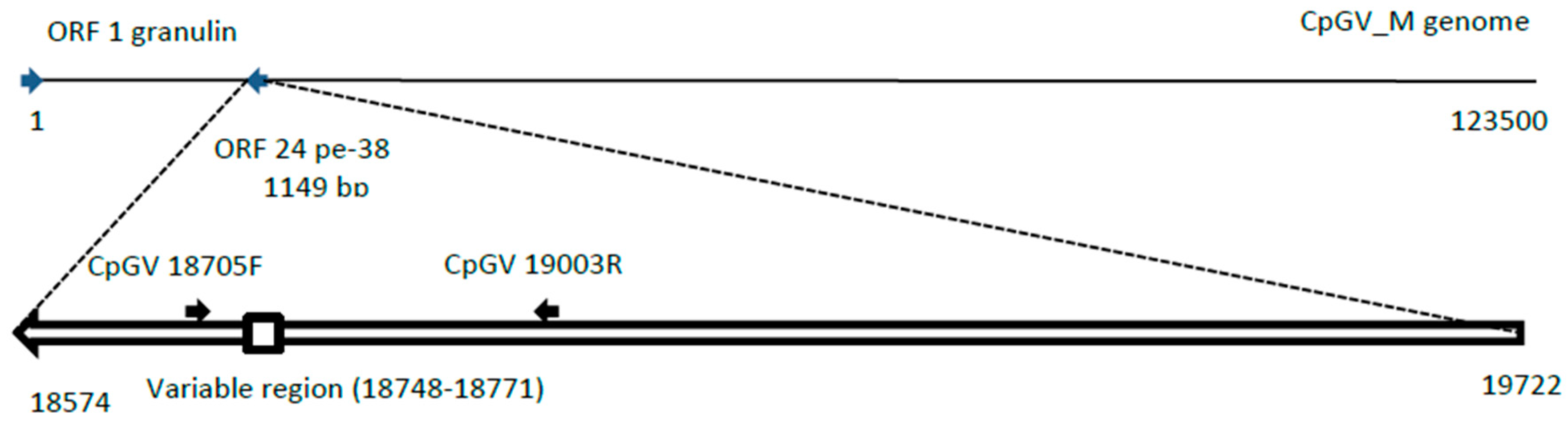
2.6. Evaluation of the Relative Proportions of Each Genotype by PCR
2.6.1. DNA Extraction
2.6.2. PCR
Identification of the Virus Genotype
3. Results
3.1. Efficacy of Viral Isolates
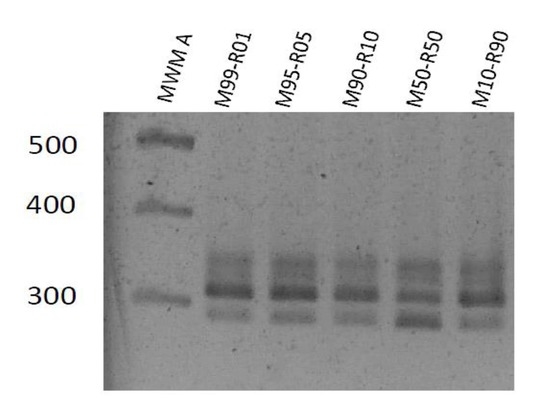
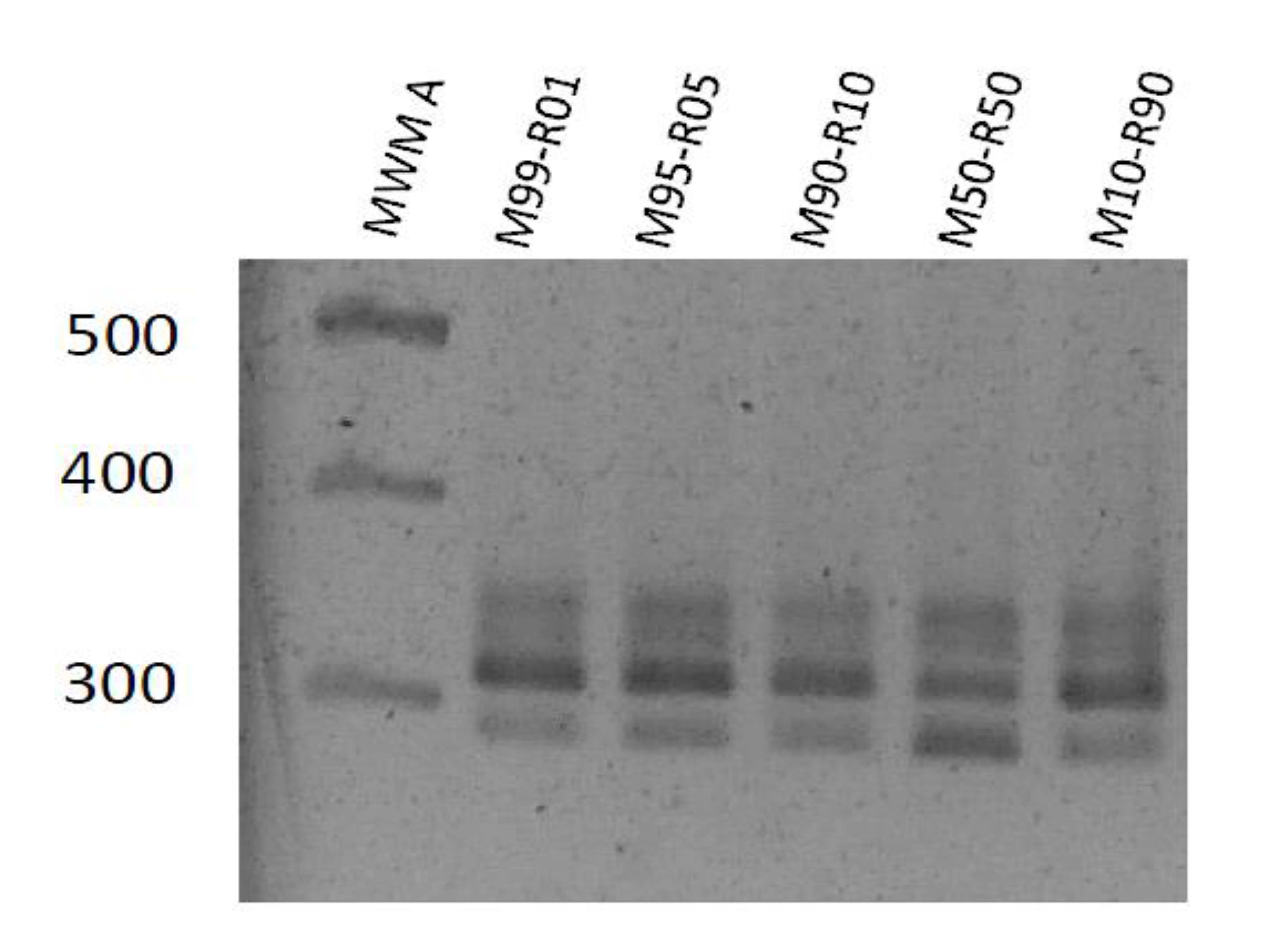
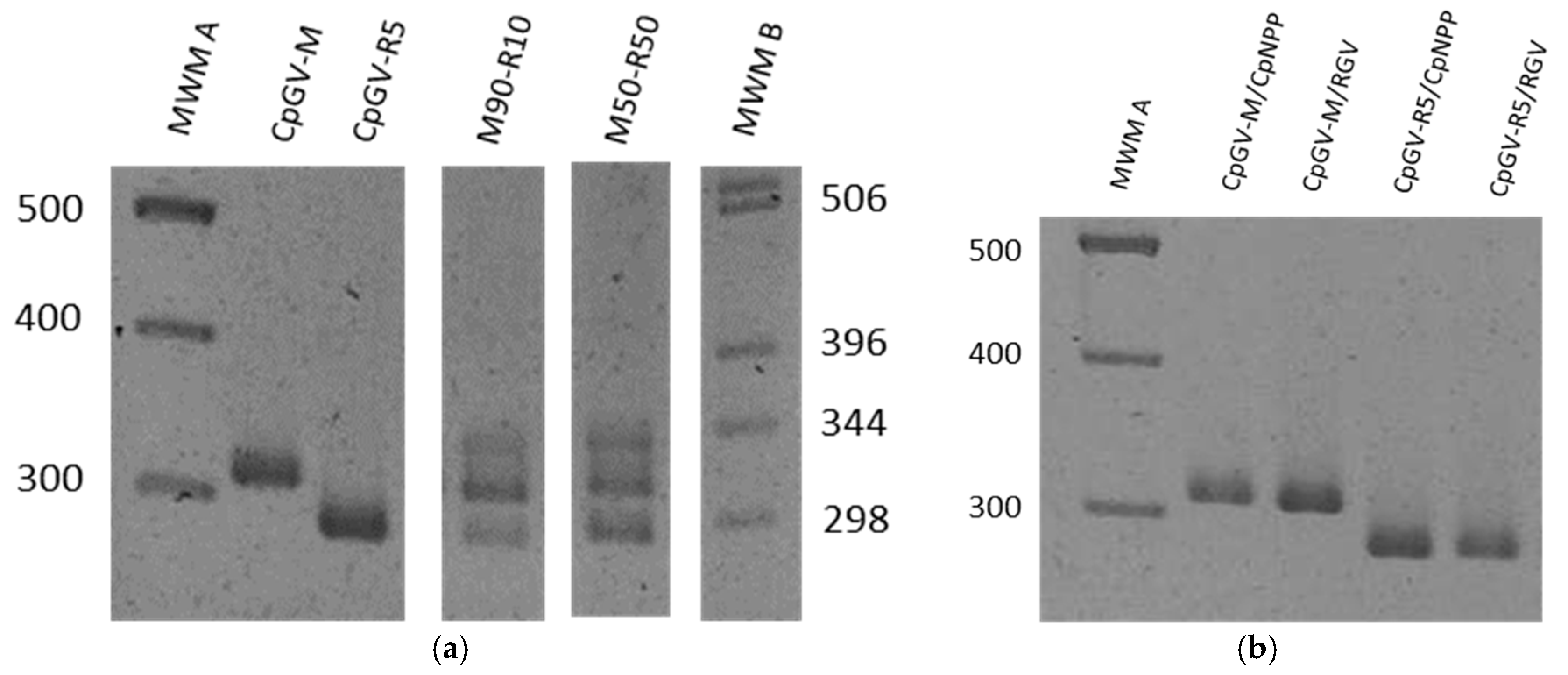
3.2. Specificity of the Genotype Markers
3.3. Replication of Viral Genotypes in Different Hosts
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- ICTV Virus Taxonomy, 2013 Release, EC 45. Available online: http://www.ictvonline.org/ (accessed on 14 September 2015).
- Tanada, Y. A granulosis virus of the codling moth, Carpocapsa pomonella (Linnaeus) (Olethreutidae, Lepidoptera). J. Insect Pathol. 1964, 6, 378–380. [Google Scholar]
- Luque, T.; Finch, R.; Crook, N.; O’Reilly, D.R.; Winstanley, D. The complete sequence of the Cydia pomonella granulovirus genome. J. Gen. Virol. 2001, 82, 2531–2547. [Google Scholar] [CrossRef] [PubMed]
- Lacey, L.A.; Thomson, D.; Vincent, C.; Arthurs, S.P. Codling moth granulovirus: A comprehensive review. Biocontrol Sci. Technol. 2008, 18, 639–663. [Google Scholar] [CrossRef]
- Crook, N.E.; Spencer, R.A.; Payne, C.C.; Leisy, D.J. Variation in Cydia pomonella granulosis virus isolates and physical maps of the DNA from three variants. J. Gen. Virol. 1985, 66, 2423–2430. [Google Scholar] [CrossRef]
- Fritsch, E.; Undorf-Span, K.; Kienle, J.; Zebitz, C.P.W.; Huber, J. Apfelwickler-Granulovirus: Erste Hinweise auf Unterschiede in der Empfindlichkeit lokaler Apfelwickler-Populationen. Nachricht. Deutsch. Pflanzenschutz. 2005, 57, 29–34. [Google Scholar]
- Sauphanor, B.; Berling, M.; Toubon, J.F.; Reyes, M.; Delnatte, J.; Allemoz, P. Carpocapse des pommes: Cas de résistance au virus de la granulose en vergers biologiques. Phytoma Def. Veg. 2006, 590, 24–27. [Google Scholar]
- Schmitt, A.; Bisutti, I.L.; Ladurner, E.; Benuzzi, M.; Sauphanor, B.; Kienzle, J.; Zingg, D.; Undorf-Spahn, K.; Fritsch, E.; Huber, J.; et al. The occurrence and distribution of resistance of codling moth to Cydia pomonella granulovirus in Europe. J. Appl. Entomol. 2013, 137, 641–649. [Google Scholar] [CrossRef]
- Asser-Kaiser, S.; Fritsch, E.; Undorf-Spahn, K.; Kienzle, J.; Eberle, K.E.; Gund, N.A.; Reineke, A.; Zebitz, C.P.W.; Heckel, D.G.; Huber, J.; et al. Rapid emergence of baculovirus resistance in codling moth due to dominant, sex-linked inheritance. Science 2007, 317, 1916–1918. [Google Scholar] [CrossRef] [PubMed]
- Asser-Kaiser, S.; Radtke, P.; El-Salamouny, S.; Winstanley, D.; Jehle, J.A. Baculovirus resistance in codling moth Cydia pomonella L. caused by early block of virus replication. Virology 2011, 410, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Rezapanah, M.; Shojai-Estabragh, S.; Huber, J.; Jehle, J.A. Molecular characterization of new isolates of Cydia pomonella granulovirus from Iran. J. Pest. Sci. 2008, 81, 187–191. [Google Scholar] [CrossRef]
- Eberle, K.E.; Asser-Kaiser, S.; Sayed, S.M. Overcoming the resistance of codling moth against conventional Cydia pomonella granulovirus (CpGV-M) by a new isolate CpGV-I12. J. Invertebr. Pathol. 2008, 98, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Berling, M.; Blachere-López, C.; Soubabere, O.; Lery, X.; Bonhomme, A.; Sauphanor, B.; López-Ferber, M. Cydia pomonella granulovirus genotypes overcome virus resistance in the codling moth and improve virus efficiency by selection against resistant hosts. Appl. Environ. Microbiol. 2009, 75, 925–930. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, M.M.; Eberle, K.E.; Radtke, P.; Jehle, J.A. Baculovirus resistance in codling moth is virus isolate-dependent and the consequence of a mutation in viral gene pe38. PNAS 2014, 111, 15711–15716. [Google Scholar] [CrossRef] [PubMed]
- Read, A.F.; Taylor, L.H. The ecology of genetically diverse infections. Science 2001, 292, 1099–1102. [Google Scholar] [CrossRef] [PubMed]
- Mideo, N. Parasite adaptations to within-host competition. Trends Parasit. 2009, 25, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, D.; Castillejo, J.I.; Caballero, P. Naturally occurring deletion mutants are parasitic genotypes in a wild-type nucleopolyhedrovirus population of Spodoptera exigua. Appl. Environ. Microbiol. 1998, 64, 4372–4377. [Google Scholar] [PubMed]
- Hitchman, R.B.; Hodgson, D.J.; King, L.A.; Hails, R.S.; Cory, J.S.; Possee, R.D. Host mediated selection of pathogen genotypes as a mechanism for the maintenance of baculovirus diversity in the field. J. Invertebr. Pathol. 2007, 94, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Possee, R.D.; Rohrmann, G.F. Baculovirus Genome Organization and Evolution. In The Baculoviruses; Miller, L.K., Ed.; Plenum Press: New York, NY, USA, 1997; pp. 109–140. [Google Scholar]
- Huber, J. Use of Baculoviruses in Pest Management Programs. In The Biology of Baculoviruses; Granados, R.R., Federici, B.A., Eds.; CRC Press: Boca Raton, FL, USA, 1986; Volume 2, pp. 182–202. [Google Scholar]
- Hoy, M.A. Myths, models and mitigation of resistance to pesticides. Phil. Trans. R. Soc. B 1998, 353, 1787–1795. [Google Scholar] [CrossRef] [PubMed]
- Briese, D.T.; Mende, H.A.; Grace, T.D.C.; Geier, P.W. Resistance to a nuclear polyhedrosis virus in the light-brown apple moth Epiphyas postvittana (Lepidoptera: Tortricidae). J. Invertebr. Pathol. 1980, 36, 211–215. [Google Scholar] [CrossRef]
- Fuxa, J.R. Ecology of insect nucleopolyhedroviruses. Agric. Ecosyst. Environ. 2004, 103, 27–43. [Google Scholar] [CrossRef]
- Lee, H.H.; Miller, L.K. Isolation of genetic variants of Autographa californica nuclear polyhedrosis virus. J. Virol. 1978, 3, 754–767. [Google Scholar]
- Graham, R.I.; Tyne, W.I.; Possee, R.D.; Sait, S.M.; Hails, R.S. Genetically variable nucleopolyhedrovirueses isolated from spatially separated populations of the winter moth Operophtera brumata (Lepidoptera: Geometridae) in Orkney. J. Invertebr. Pathol. 2004, 87, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Simón, O.; Williams, T.; López-Ferber, M.; Caballero, P. Genetic structure of a Spodoptera frugiperda nucleopolyhedrovirus population: High prevalence of deletion genotypes. Appl. Environ. Microbiol. 2004, 70, 5579–5588. [Google Scholar] [CrossRef] [PubMed]
- Eberle, K.E.; Sayed, S.; Rezapanah, M.; Shojai-Estabragh, S.; Jehle, J.A. Diversity and evolution of the Cydia pomonella granulovirus. J. Gen. Virol. 2009, 90, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Vickers, J.M.; Cory, J.S.; Entwistle, P.F. DNA characterization of eight geographic isolates of granulosis viruses from the potato tuber moth Phthorimaea operculella (Zeller) (Lepidoptera, Gelechiidae). J. Invertebr. Pathol. 1991, 57, 334–342. [Google Scholar] [CrossRef]
- Bull, J.C.; Godfray, H.C.J.; O’Reilly, D.R. Persistance of an occlusion-negative recombinant nucleopolyhedrovirus in Trichoplusia ni indicates high multiplicity of cellular infection. Appl. Environ. Microbiol. 2001, 67, 5204–5209. [Google Scholar] [CrossRef] [PubMed]
- López-Ferber, M.; Simón, O.; Williams, T.; Caballero, P. Defective or effective? Mutualistic interactions between virus genotypes. Proc. R. Soc. B 2003, 270, 2249–2255. [Google Scholar] [CrossRef] [PubMed]
- Clavijo, G.; Williams, T.; Simón, O.; Muñoz, D.; Cerutti, M.; López-Ferber, M.; Caballero, P. Mixtures of complete and pif1- and pif2-deficient genotypes are required for increased potency of an insect nucleopolyhedrovirus. J. Virol. 2009, 83, 5127–5136. [Google Scholar] [CrossRef] [PubMed]
- Simón, O.; Williams, T.; López-Ferber, M.; Caballero, P. Functional importance of deletion mutant genotypes in an insect nucleopolyhedrovirus population. Appl. Environ. Microbiol. 2005, 71, 4254–4262. [Google Scholar] [CrossRef] [PubMed]
- Graillot, B.; Berling, M.; Blachere-López, C.; Siegwart, M.; Besse, S.; López-Ferber, M. Progressive adaptation of a CpGV isolate to codling moth populations resistant to CpGV-M. Viruses 2014, 6, 5135–5144. [Google Scholar] [CrossRef] [PubMed]
- Graillot, B.; Berling, M.; Blachere-López, C.; Siegwart, M.; Besse, S.; López-Ferber, M. Correction: Progressive adaptation of a CpGV isolate to codling moth populations resistant to CpGV-M. Viruses 2015, 7, 6313–6315. [Google Scholar] [CrossRef]
- Finney, J.R. Probit Analisis, 3rd ed.; Cambridge University Press: Cambridge, UK, 1971. [Google Scholar]
- LeOra. POLO-PC SOFTWARE. A User’s Manual for Probit or Logit Analysis; LeOra Software: Berkeley, CA, USA, 1987. [Google Scholar]
- Tanada, Y.; Hukuhara, T. Enhanced infection of a nuclear-polyhedrosis virus in larvae of the armyworm, Pseudaletia unipuncta, by a factor in the capsule of a granulosis virus. J. Invertebr. Pathol. 1971, 17, 116–126. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Corsaro, B.G.; Granados, R.R. Location and nucleotide sequence of the gene encoding the viral enhancing factor of the Trichoplusia ni granulosis virus. J. Gen. Virol. 1991, 72, 2645–2651. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.; van Lent, J.W.; Boeren, S.; Fang, M.; Theilmann, D.A.; Erlandson, M.A.; Vlak, J.M.; van Oers, M.M. Characterization of novel components of the baculovirus per os infectivity factor complex. J. Virol. 2012, 86, 4981–4988. [Google Scholar] [CrossRef] [PubMed]
- Kikhno, I.; Gutiérrez, S.; Croizier, L.; Croizier, G.; López-Ferber, M. Characterization of pif, a gene required for the per os infectivity of Spodoptera littoralis nucleopolyhedrovirus. J. Gen. Virol. 2002, 83, 3013–3022. [Google Scholar] [CrossRef] [PubMed]
- Clavijo, G.; Williams, T.; Muñoz, D.; Caballero, P.; López-Ferber, M. Mixed genotype transmission bodies and virions contribute to the maintenance of diversity in an insect virus. Proc. Roy. Soc. B 2010, 277, 943–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Insect Strain | Composition of Virus Mixtures (%) | No. Controls | No. Insects Tested | Lethal Concentrations in OBs/µL (95% CI) | Slope ± SE | χ2 | |||
|---|---|---|---|---|---|---|---|---|---|
| CpGV-M | CpGV-R5 | LC50 | LC90 | ||||||
| 1 | CpNPP | 100 | 0 | 257 | 529 | 13.10 (6.55–23.20) | 223.10 (110.70–654.18) | 1.04 ± 0.09 | 5.99 |
| 2 | 90 | 10 | 256 | 541 | 12.40 (4.92–25.42) | 119.03 (52.17–616.73) | 1.31 ± 0.12 | 11.95 | |
| 3 | 50 | 50 | 257 | 518 | 10.69 (6.67–16.11) | 94.12 (56.48–197.99) | 1.36 ± 0.12 | 4.26 | |
| 4 | 0 | 100 | 257 | 533 | 6.76 (2.59–13.37) | 59.63 (27.54−278.55) | 1.36 ± 0.13 | 11.42 | |
| 5 | RGV | 100 | 0 | 476 | 1143 | 2.22 × 106 (1.19 × 106–5.67 × 106) | - | 0.50 ± 0.07 | 10.6 |
| 6 | 90 | 10 | 215 | 817 | 201.78 (139.95–280.83) | 2.35 × 103 (1.51 × 103–4.23 × 103) | 1.20 ± 0.09 | 5.46 | |
| 7 | 50 | 50 | 354 | 989 | 16.45 (8.41–29.07) | 311.55 (151.53–938.21) | 1.00 ± 0.06 | 16.35 | |
| 8 | 0 | 100 | 176 | 369 | 22.43 (13.73–34.36) | 410.67 (240.16–846.43) | 1.02 ± 0.11 | 3.60 | |
| Insect Strain | Composition of Virus Mixtures (%) | Chi-Square | Degrees of Freedom | P | |
|---|---|---|---|---|---|
| CpGV-M | CpGV-R5 | ||||
| CpNPP | 50 | 50 | 91.002 | 6 | <0.01 |
| 90 | 10 | 8.98 | 6 | >0.05 | |
| RGV | 50 | 50 | 40.276 | 6 | <0.01 |
| 90 | 10 | 4.581 | 6 | >0.05 | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Graillot, B.; Bayle, S.; Blachere-Lopez, C.; Besse, S.; Siegwart, M.; Lopez-Ferber, M. Biological Characteristics of Experimental Genotype Mixtures of Cydia Pomonella Granulovirus (CpGV): Ability to Control Susceptible and Resistant Pest Populations. Viruses 2016, 8, 147. https://doi.org/10.3390/v8050147
Graillot B, Bayle S, Blachere-Lopez C, Besse S, Siegwart M, Lopez-Ferber M. Biological Characteristics of Experimental Genotype Mixtures of Cydia Pomonella Granulovirus (CpGV): Ability to Control Susceptible and Resistant Pest Populations. Viruses. 2016; 8(5):147. https://doi.org/10.3390/v8050147
Chicago/Turabian StyleGraillot, Benoit, Sandrine Bayle, Christine Blachere-Lopez, Samantha Besse, Myriam Siegwart, and Miguel Lopez-Ferber. 2016. "Biological Characteristics of Experimental Genotype Mixtures of Cydia Pomonella Granulovirus (CpGV): Ability to Control Susceptible and Resistant Pest Populations" Viruses 8, no. 5: 147. https://doi.org/10.3390/v8050147
APA StyleGraillot, B., Bayle, S., Blachere-Lopez, C., Besse, S., Siegwart, M., & Lopez-Ferber, M. (2016). Biological Characteristics of Experimental Genotype Mixtures of Cydia Pomonella Granulovirus (CpGV): Ability to Control Susceptible and Resistant Pest Populations. Viruses, 8(5), 147. https://doi.org/10.3390/v8050147