
The p22 RNA Silencing Suppressor of the Crinivirus Tomato chlorosis virus is Dispensable for Local Viral Replication but Important for Counteracting an Antiviral RDR6-Mediated Response during Systemic Infection



Abstract
:
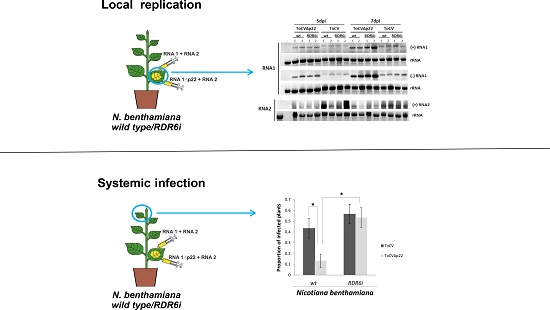{kind=link}
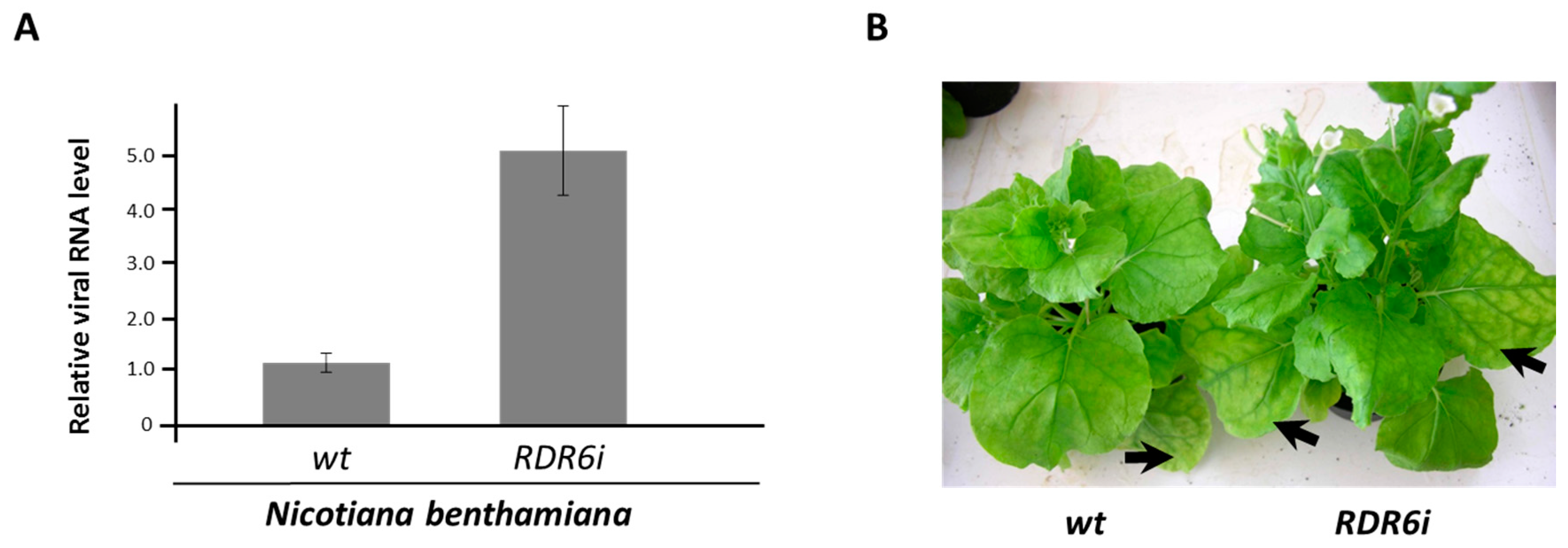{kind=link}
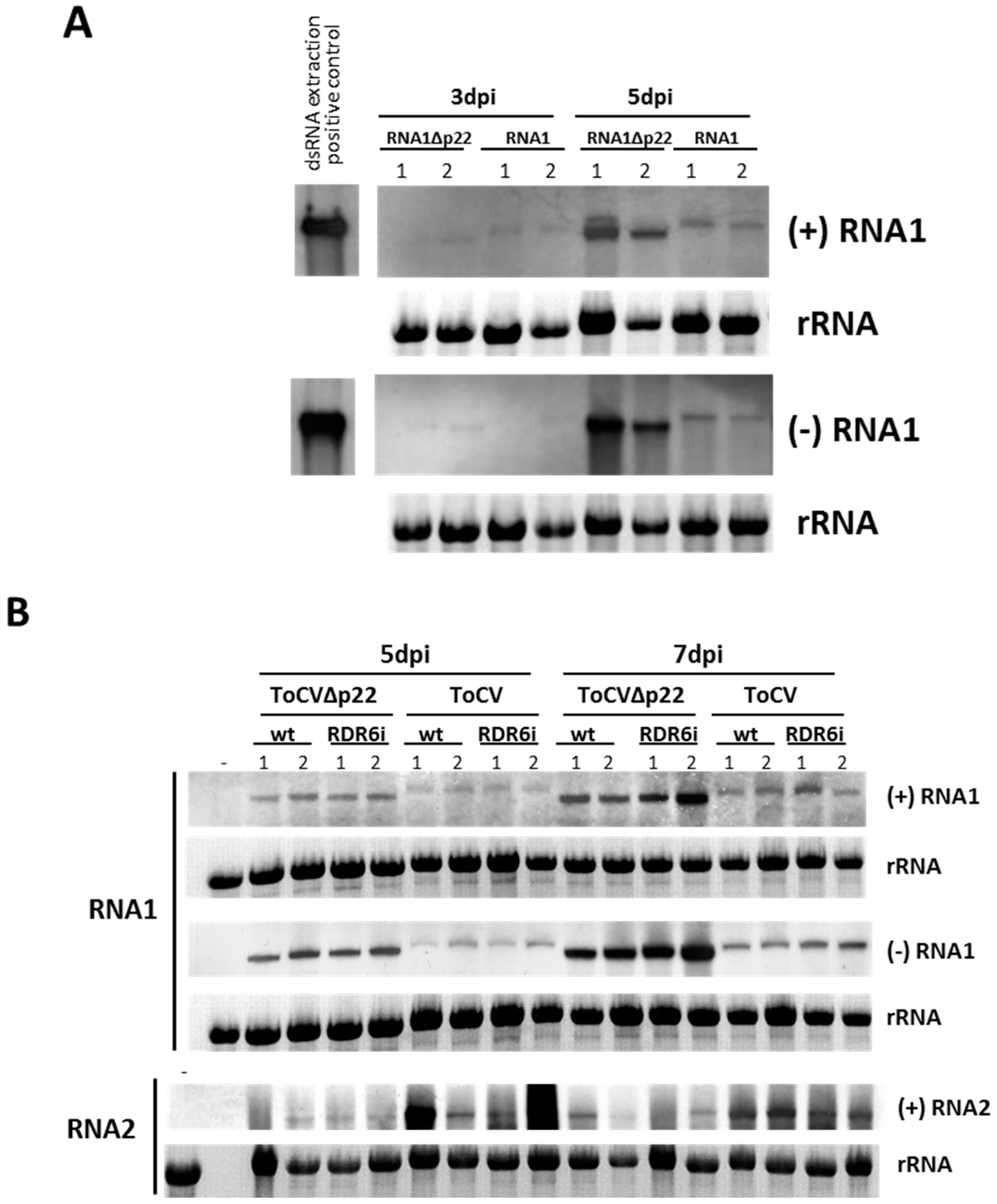{kind=link}
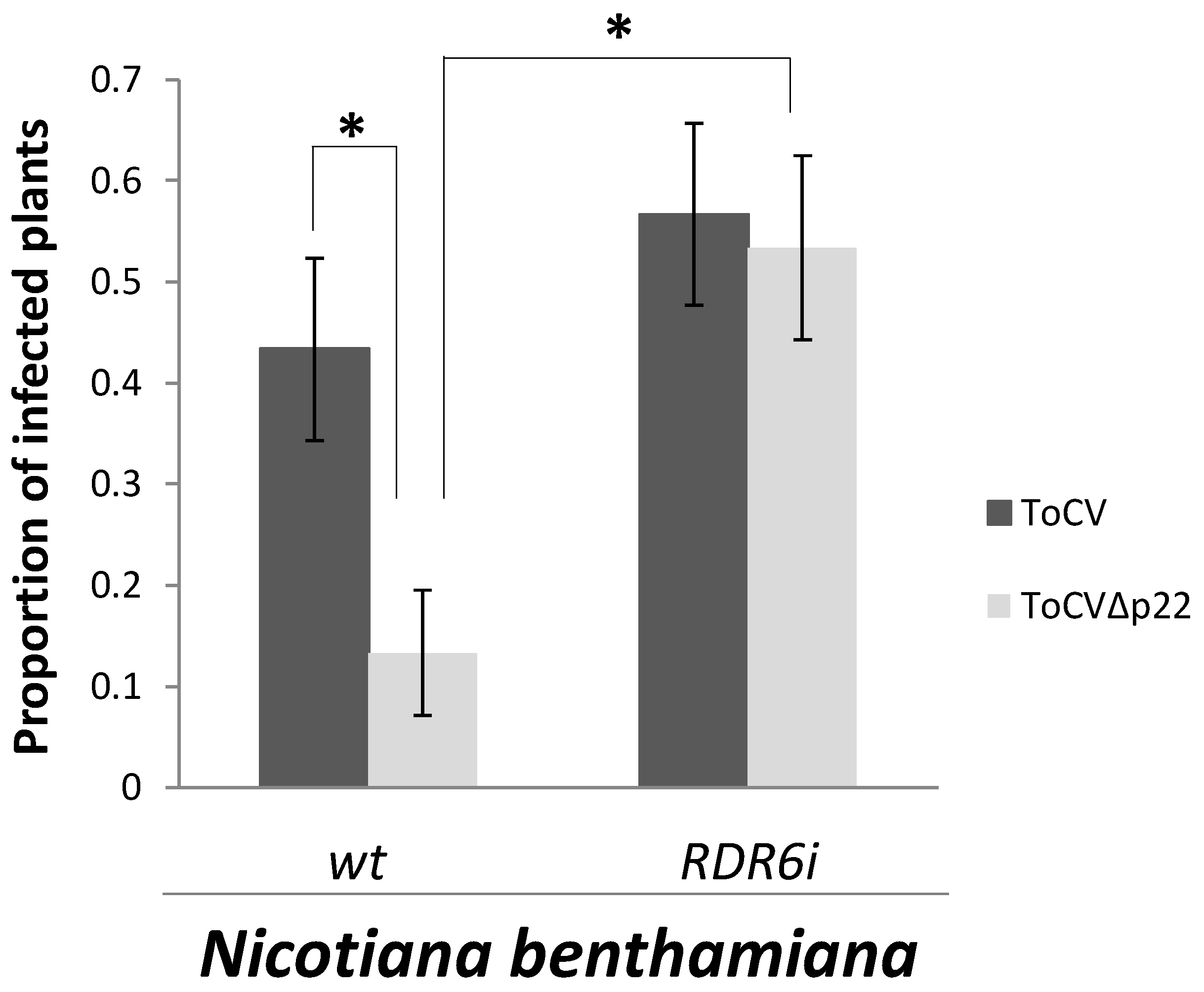{kind=link}
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Baulcombe, D. RNA silencing in plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Llave, C. Virus-derived small interfering RNAs at the core of plant-virus interactions. Trends Plant Sci. 2010, 15, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.J.; Baulcombe, D. A species of small antisense RNA in posttranscriptional gene silencing in plants. Science 1999, 286, 950–952. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Qian, D.; Zheng, H.; Meng, L.Y.; Chen, J.; Le, W.J.; Zhou, T.; Zhou, Y.J.; Wei, C.H.; Li, Y. RNA-dependent RNA polymerase 6 of rice (Oryza sativa) plays role in host defense against negative-strand RNA virus, Rice stripe virus. Virus Res. 2012, 163, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Qu, F.; Ye, X.; Hou, G.; Sato, S.; Clemente, T.E.; Morris, T.J. RDR6 has a broad-spectrum but temperature-dependent antiviral defense role in Nicotiana benthamiana. J. Virol. 2005, 79, 15209–15217. [Google Scholar] [CrossRef] [PubMed]
- Schwach, F.; Vaistij, F.E.; Jones, L.; Baulcombe, D.C. An RNA-dependent RNA polymerase prevents meristem invasion by Potato virus X and is required for the activity but not the production of a systemic silencing signal. Plant Physiol. 2005, 138, 1842–1852. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Shi, N.; Gu, M.; Zhang, H.; Li, B.; Shen, J.; Mohammed, A.; Ryabov, E.; Li, C.; Wang, H.; Liu, Y.; Osman, T.; Vatish, M.; Hong, T. Involvement of RDR6 in short-range intercellular RNA silencing in Nicotiana benthamiana. Sci. Rep. 2012, 2, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. Use, tolerance and avoidance of amplified RNA silencing by plants. Trends Plant Sci. 2008, 13, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Pendón, J.A.; Ding, S.W. Direct and indirect roles of viral suppressors of RNA silencing in pathogenesis. Annu. Rev. Phytopathol. 2008, 46, 303–326. [Google Scholar] [CrossRef] [PubMed]
- Dolja, V.V.; Kreuze, J.F.; Valkonen, J.P.T. Comparative and functional genomics of closteroviruses. Virus Res. 2006, 117, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Wintermantel, W.M.; Wisler, G.C. Vector specificity, host range, and genetic diversity of Tomato chlorosis virus. Plant Dis. 2006, 90, 814–819. [Google Scholar] [CrossRef]
- Lozano, G.; Moriones, E.; Navas-Castillo, J. Complete nucleotide sequence of the RNA2 of the crinivirus Tomato chlorosis virus. Arch. Virol. 2006, 151, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Lozano, G.; Moriones, E.; Navas-Castillo, J. Complete sequence of the RNA1 of a European isolate of Tomato chlorosis virus. Arch. Virol. 2007, 152, 839–841. [Google Scholar] [CrossRef] [PubMed]
- Wintermantel, W.M.; Wisler, G.C.; Anchieta, A.G.; Liu, H.Y.; Karasev, A.V.; Tzanetakis, I.E. The complete nucleotide sequence and genome organization of Tomato chlorosis virus. Arch. Virol. 2005, 150, 2287–2298. [Google Scholar] [CrossRef] [PubMed]
- Wisler, G.C.; Li, R.H.; Liu, H.Y.; Lowry, D.S.; Duffus, J.E. Tomato chlorosis virus: A new whitefly-transmitted, phloem-limited, bipartite closterovirus of tomato. Phytopathology 1998, 88, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Orílio, F.A.; Fortes, I.M.; Navas-Castillo, J. Infectious cDNA clones of the crinivirus Tomato chlorosis virus are competent for systemic plant infection and whitefly-transmission. Virology 2014, 464, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Kreuze, J.F.; Savenkov, E.I.; Cuellar, W.; Li, X.; Valkonen, J.P.T. Viral class 1 RNase III involved in suppression of RNA silencing. J. Virol. 2005, 79, 7227–7238. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Folimonov, A.; Shintaku, M.; Li, W.X.; Falk, B.W.; Dawson, W.O.; Ding, S.W. Three distinct suppressors of RNA silencing encoded by a 20-kb viral RNA genome. Proc. Natl. Acad. Sci. USA 2004, 101, 15742–15747. [Google Scholar] [CrossRef] [PubMed]
- Cañizares, M.C.; Navas-Castillo, J.; Moriones, E. Multiple suppressors of RNA silencing encoded by both genomic RNAs of the crinivirus, Tomato chlorosis virus. Virology 2008, 379, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Satyanarayana, T.; Gowda, S.; Ayllón, M.A.; Albiach-Martí, M.R.; Rabindran, S.; Dawson, W.O. The p23 protein of Citrus tristeza virus controls asymmetrical RNA accumulation. J. Virol. 2002, 76, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.C.; Kassschau, K.D.; Prohnevsky, A.I.; Gopinath, K.; Pogue, G.P.; Carrington, J.C.; Dolja, V.V. Suppressor of RNA silencing encoded by Beet yellows virus. Virology 2003, 306, 203–209. [Google Scholar] [CrossRef]
- Peremyslov, V.Y.; Hagiwara, Y.; Dolja, V.V. Genes required for replication of the 15.5-kilobase RNA genome of a plant closterovirus. J. Virol. 1998, 72, 5870–5876. [Google Scholar] [PubMed]
- Yeh, H.H.; Tian, T.; Rubio, L.; Crawford, B.; Falk, B.W. Asynchronous accumulation of Lettuce infectious yellows virus RNAs 1 and 2 and identification of an RNA1 trans enhancer of RNA2 accumulation. J. Virol. 2000, 74, 5762–5768. [Google Scholar] [CrossRef] [PubMed]
- Noris, E.; Accotto, G.P.; Tavazza, R.; Brunetti, A.; Crespi, S.; Tavazza, M. Resistance to tomato yellow leaf curl geminivirus in Nicotiana benthamiana plants transformed with a truncated viral C1 gene. Virology 1996, 225, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, P.; Wrzesinska, B.; Obrepalska-Steplowska, A. Assessment of reference gene stability influenced by extremely divergent disease symptoms in Solanum lycopersicum. J. Virol. Methods 2013, 194, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and de 2T∆∆C method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Hechman, K.L.; Pease, L.R. Gene splicing and mutagenesis by PCR-driven overlap extension. Nat. Protoc. 2007, 2, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef] [PubMed]
- García-Ruiz, H.; Takeda, A.; Chapman, E.J.; Sullivan, C.M.; Fahlgren, N.; Brempelis, K.J.; Carrington, J.C. Arabidopsis RNA-dependent RNA polymerases and dicer-like proteins in antiviral defense and small interfering RNA biogenesis during Turnip mosaic virus infection. Plant Cell 2010, 22, 481–496. [Google Scholar]
- Wang, X.B.; Wu, Q.; Ito, T.; Cillo, F.; Li, W.X.; Chen, X.; Yu, J.L.; Ding, S.W. RNAi-mediated viral immunity requires amplification of virus derived siRNAs in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.B.; Jovel, J.; Udomporn, P.; Wang, Y.; Wu, Q.; Li, W.X.; Gasciolli, V.; Vaucheret, H.; Ding, S.W. The 21-nucleotide, but not 22-nucleotide, viral secondary small interfering RNAs direct potent antiviral defense by two cooperative argonautes in Arabidopsis thaliana. Plant Cell 2011, 23, 1625–1638. [Google Scholar] [CrossRef] [PubMed]
- Landeo-Ríos, Y.M.; Navas-Castillo, J.; Moriones, E.; Cañizares, M.C. The p22 RNA silencing suppressor of the crinivirus Tomato chlorosis virus preferentially binds long dsRNAs preventing them from cleavage. Virology 2016, 488, 129–136. [Google Scholar] [CrossRef] [PubMed]



© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Landeo-Ríos, Y.; Navas-Castillo, J.; Moriones, E.; Cañizares, M.C. The p22 RNA Silencing Suppressor of the Crinivirus Tomato chlorosis virus is Dispensable for Local Viral Replication but Important for Counteracting an Antiviral RDR6-Mediated Response during Systemic Infection. Viruses 2016, 8, 182. https://doi.org/10.3390/v8070182
Landeo-Ríos Y, Navas-Castillo J, Moriones E, Cañizares MC. The p22 RNA Silencing Suppressor of the Crinivirus Tomato chlorosis virus is Dispensable for Local Viral Replication but Important for Counteracting an Antiviral RDR6-Mediated Response during Systemic Infection. Viruses. 2016; 8(7):182. https://doi.org/10.3390/v8070182
Chicago/Turabian StyleLandeo-Ríos, Yazmín, Jesús Navas-Castillo, Enrique Moriones, and M. Carmen Cañizares. 2016. "The p22 RNA Silencing Suppressor of the Crinivirus Tomato chlorosis virus is Dispensable for Local Viral Replication but Important for Counteracting an Antiviral RDR6-Mediated Response during Systemic Infection" Viruses 8, no. 7: 182. https://doi.org/10.3390/v8070182
APA StyleLandeo-Ríos, Y., Navas-Castillo, J., Moriones, E., & Cañizares, M. C. (2016). The p22 RNA Silencing Suppressor of the Crinivirus Tomato chlorosis virus is Dispensable for Local Viral Replication but Important for Counteracting an Antiviral RDR6-Mediated Response during Systemic Infection. Viruses, 8(7), 182. https://doi.org/10.3390/v8070182