
Effects of Female Sex Hormones on Susceptibility to HSV-2 in Vaginal Cells Grown in Air-Liquid Interface

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
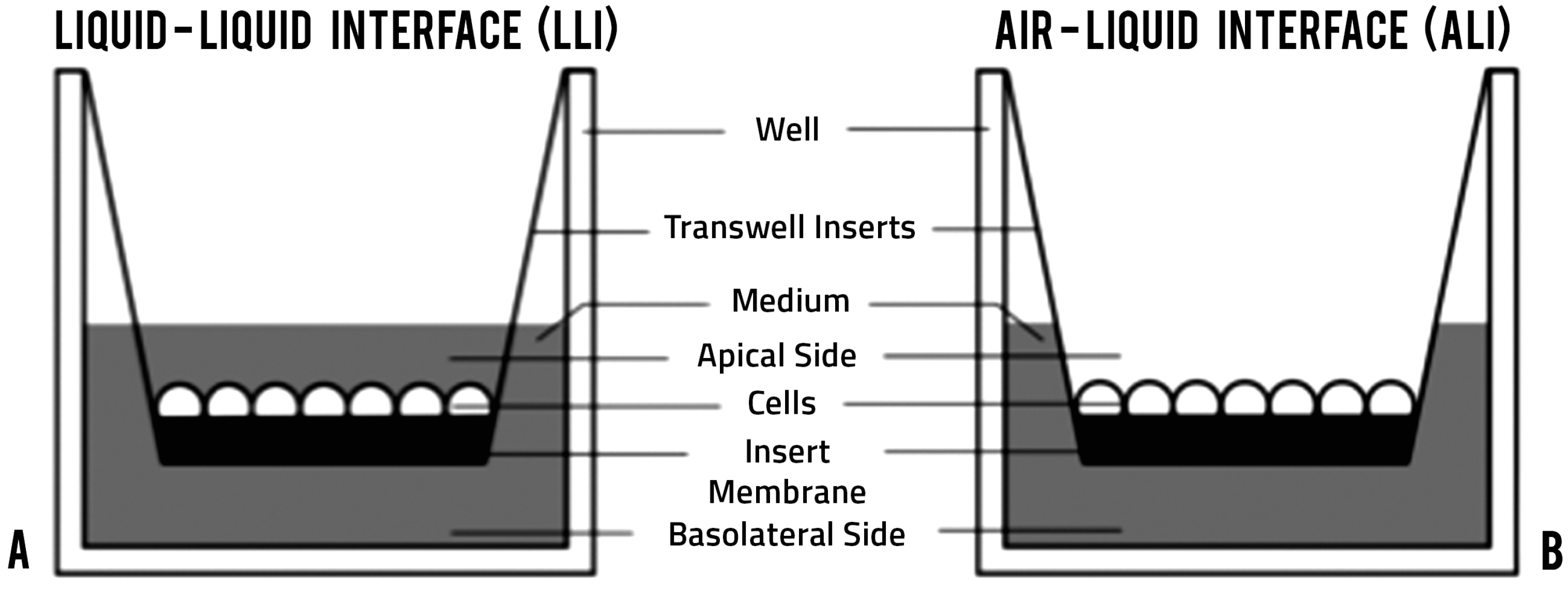
2.1. Generating Air-Liquid Interface (ALI) and Liquid-Liquid Interface (LLI) Cultures of Vk2 Cell Line
2.2. Hematoxylin and Eosin and Actin Staining of ALI and LLI Cultures
2.3. Pancytokeratin Staining of ALI and LLI Cultures Exposed to Female Sex Hormones and MPA
2.4. Cell Viability Assay
2.5. HSV-2 Infection in LLI and ALI Culture Exposed to Female Sex Hormones and MPA
2.6. Statistical Analysis
3. Results
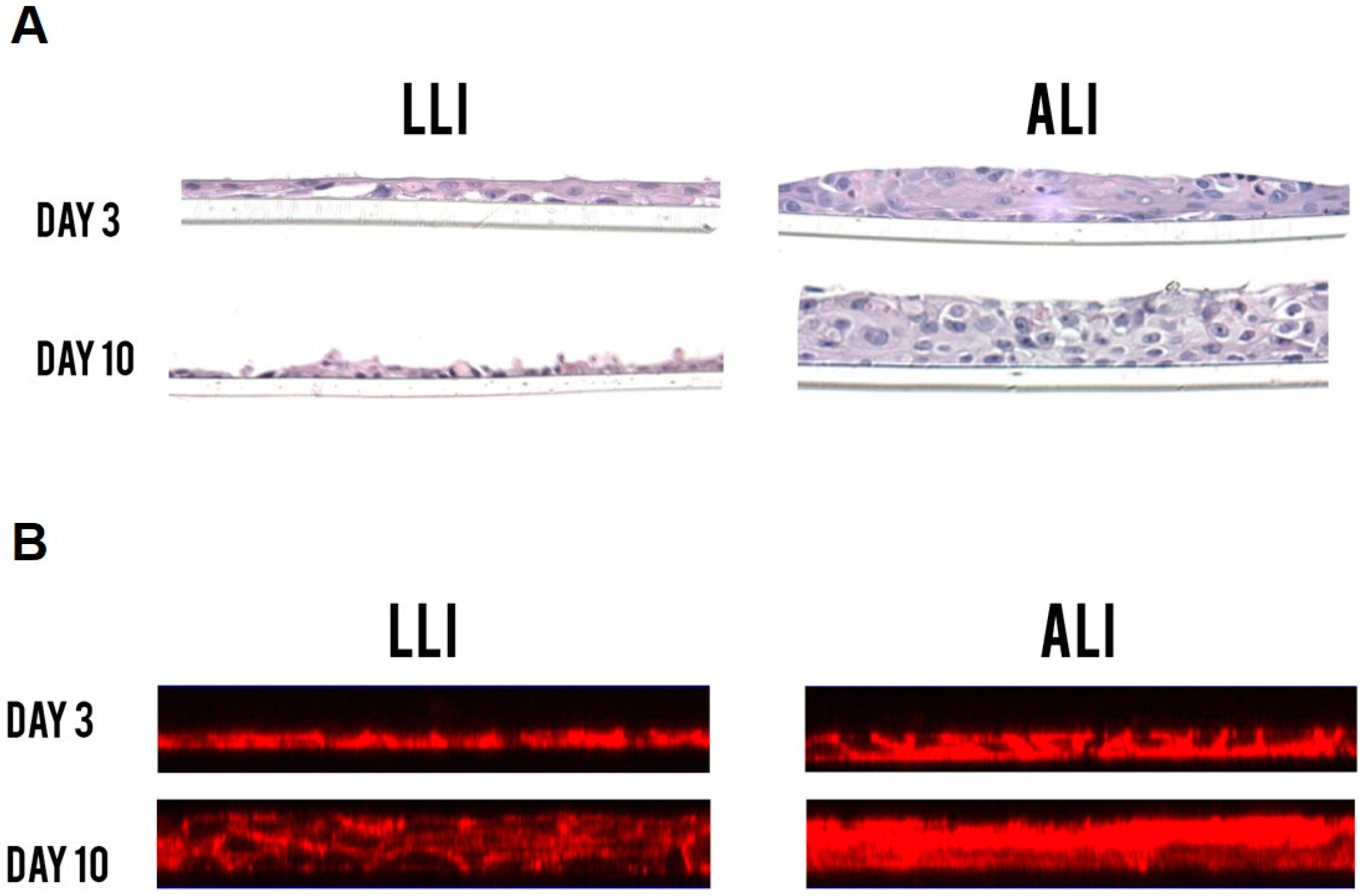
3.1. Vk2 Cells Grow into Multi-Layered Cultures in ALI Conditions
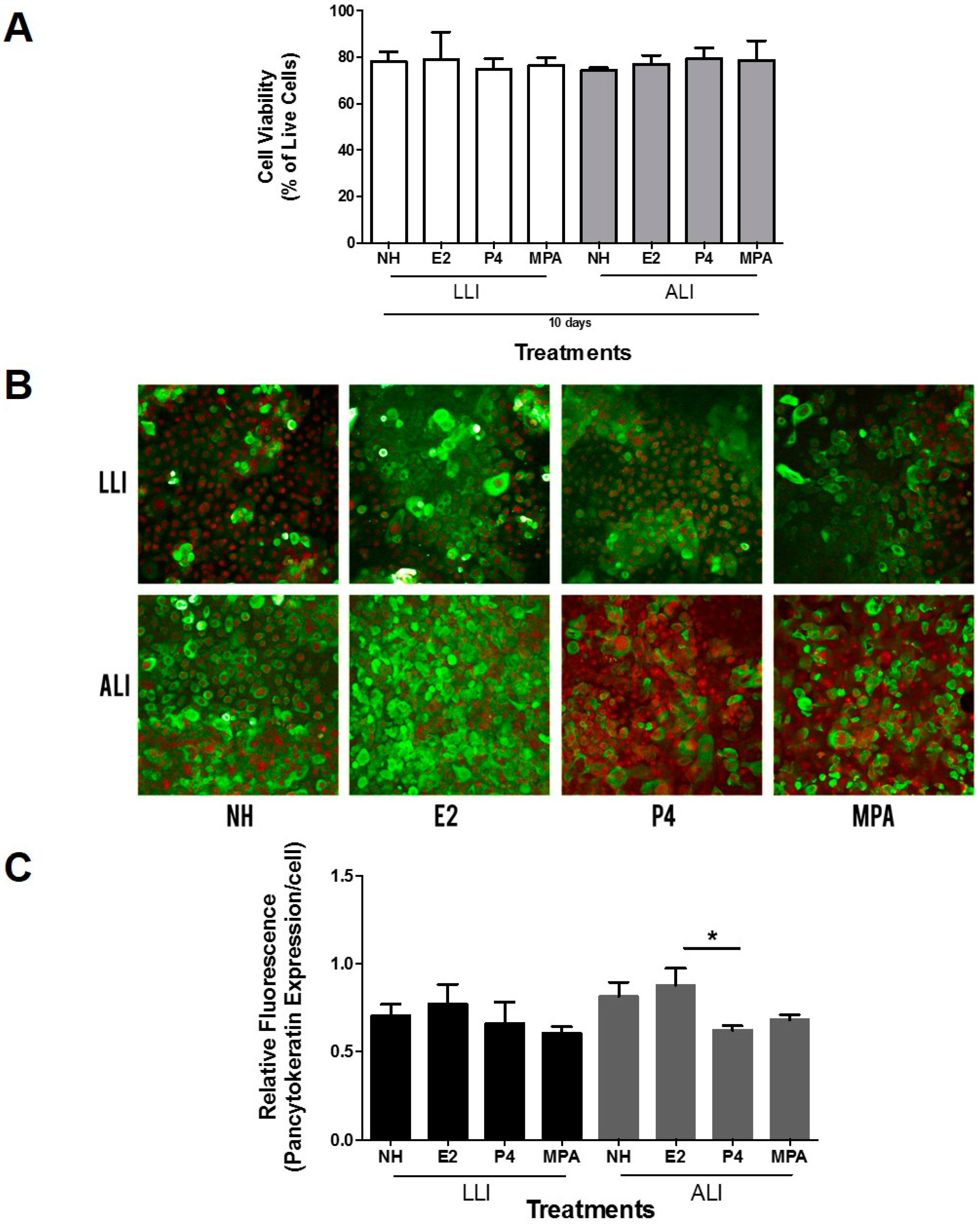
3.2. Estradiol Increases Cytokeratin Expression
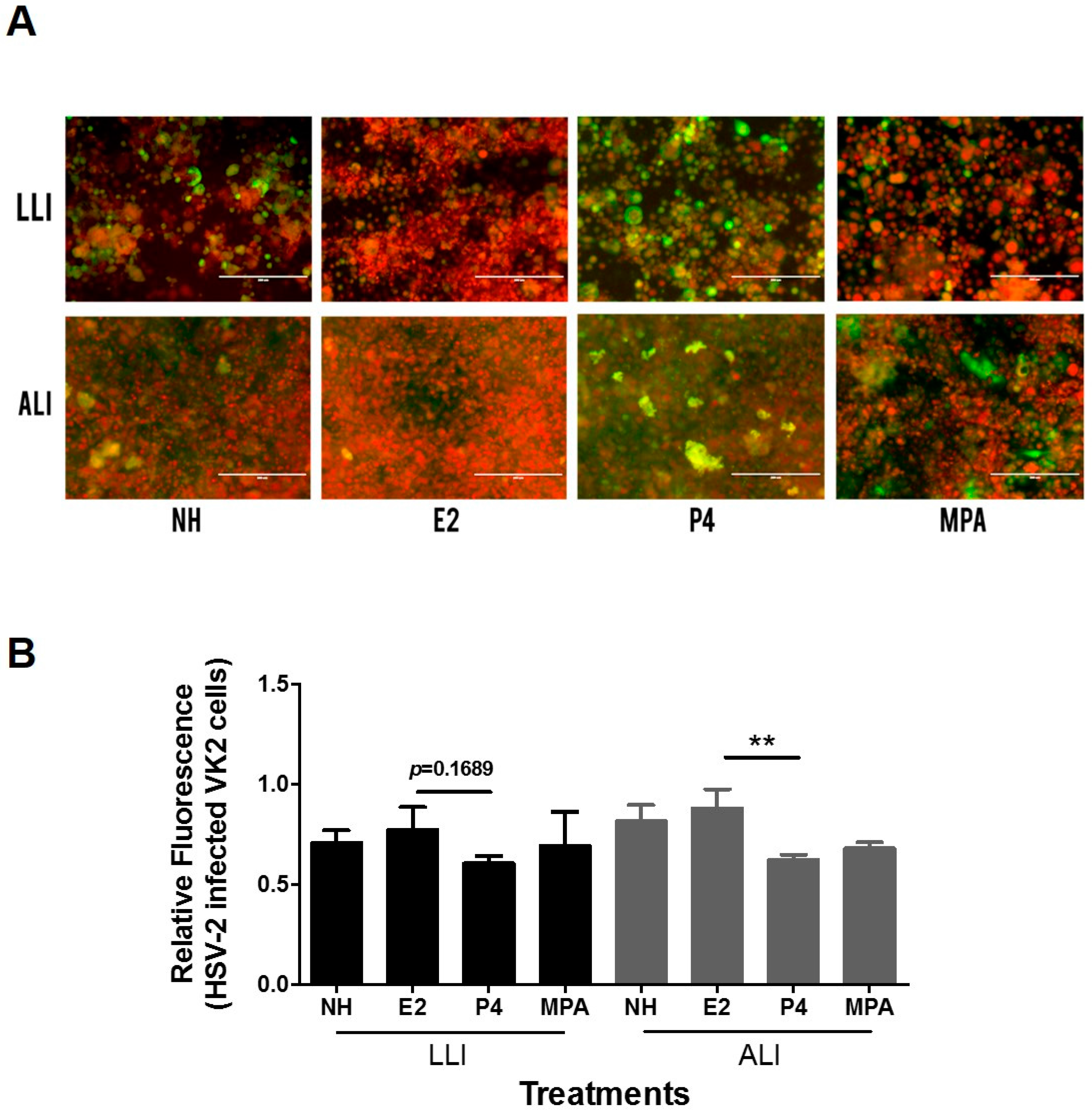
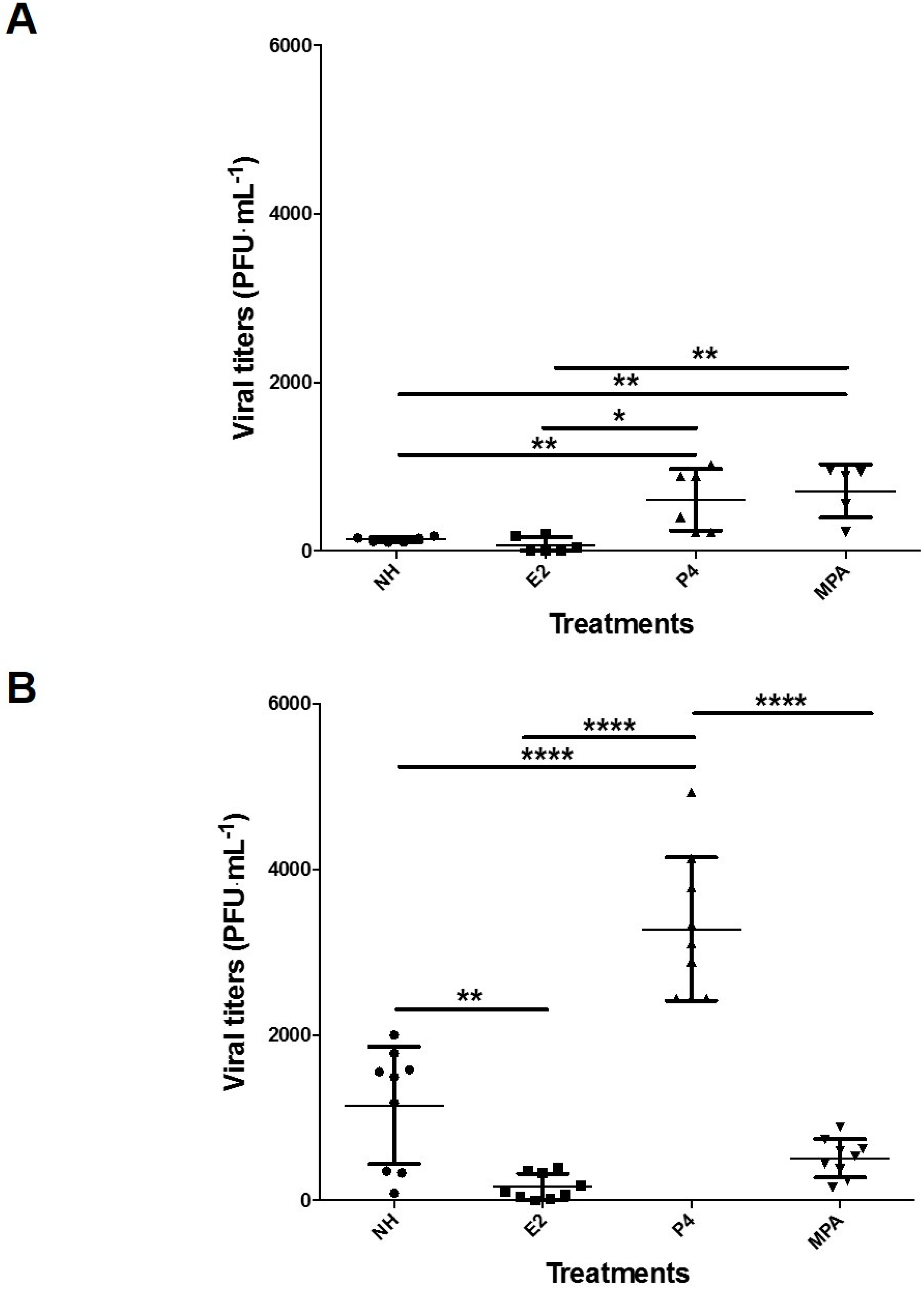
3.3. HSV-2/GFP Infection in Vk2 Cells Cultured in Progesterone Is Higher Than in Estradiol
3.4. Progesterone Increases Viral Replication in Vk2 Cells in Both ALI and LLI Cultures
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Looker, K.J.; Magaret, A.S.; Turner, K.M.E.; Vickerman, P.; Gottlieb, S.L.; Newman, L.M. Global Estimates of Prevalent and Incident Herpes Simplex Virus Type 2 Infections in 2012. PLoS ONE 2015, 10, e114989. [Google Scholar] [CrossRef] [PubMed]
- Fleming, D.T.; McQuillan, G.M.; Johnson, R.E.; Nahmias, A.J.; Aral, S.O.; Lee, F.K.; St. Louis, M.E. Herpes Simplex Virus Type 2 in the United States, 1976 to 1994. N. Engl. J. Med. 1997, 337, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.; Dolin, R.; Blaser, M. Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases. Am. Med. Assoc. 2014, 304, 2067–2701. [Google Scholar]
- Wira, C.R.; Rodriguez-Garcia, M.; Patel, M.V. The role of sex hormones in immune protection of the female reproductive tract. Nat. Rev. Immunol. 2015, 15, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Gillgrass, A.E.; Fernandez, S.A.; Rosenthal, K.L.; Kaushic, C. Estradiol regulates susceptibility following primary exposure to genital herpes simplex virus type 2, while progesterone induces inflammation. J. Virol. 2005, 79, 3107–3116. [Google Scholar] [CrossRef] [PubMed]
- Kaushic, C.; Roth, K.L.; Anipindi, V.; Xiu, F. Increased prevalence of sexually transmitted viral infections in women: The role of female sex hormones in regulating susceptibility and immune responses. J. Reprod. Immunol. 2011, 88, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Brotman, R.M.; Ravel, J.; Bavoil, P.M.; Gravitt, P.E.; Ghanem, K.G. Microbiome, sex hormones, and immune responses in the reproductive tract: Challenges for vaccine development against sexually transmitted infections. Vaccine 2014, 32, 1543–1552. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; Iwasaki, A. Generating protective immunity against genital herpes. Trends Immunol. 2013, 34, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Shattock, R.J.; Haynes, B.F.; Pulendran, B.; Flores, J.; Esparza, J. Improving defences at the portal of HIV entry: Mucosal and innate immunity: A summary report from a global HIV vaccine enterprise working group. PLoS Med. 2008, 5, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Wira, C.R.; Grant-Tschudy, K.S.; Crane-Godreau, M.A. Epithelial cells in the female reproductive tract: A central role as sentinels of immune protection. Am. J. Reprod. Immunol. 2005, 53, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Vroling, A.B.; Fokkens, W.J.; van Drunen, C.M. How epithelial cells detect danger: Aiding the immune response. Allergy 2008, 63, 1110–1123. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.J.; Marathe, J.; Pudney, J. The structure of the human vaginal stratum corneum and its role in immune defense. Am. J. Reprod. Immunol. 2014, 71, 618–623. [Google Scholar] [CrossRef] [PubMed]
- Bragulla, H.H.; Homberger, D.G. Structure and functions of keratin proteins in simple, stratified, keratinized and cornified epithelia. J. Anat. 2009, 214, 516–559. [Google Scholar] [CrossRef] [PubMed]
- Parr, M.B.; Parr, E.L. Protective Immunity against HSV-2 in the Mouse Vagina. J. Reprod. Immunol. 1997, 36, 77–92. [Google Scholar] [CrossRef]
- Wira, C.R.; Fahey, J.V.; Ghosh, M.; Patel, M.V.; Hickey, D.K.; Ochiel, D.O. Sex hormone regulation of innate immunity in the female reproductive tract: the role of epithelial cells in balancing reproductive potential with protection against sexually transmitted pathogens. Am. J. Reprod. Immunol. 2010, 63, 544–565. [Google Scholar] [CrossRef] [PubMed]
- Wira, C.R.; Fahey, J.V.; Sentman, C.L.; Pioli, P.A.; Shen, L. Innate and adaptive immunity in female genital tract: Cellular responses and interactions. Immunol. Rev. 2005, 206, 306–335. [Google Scholar] [CrossRef] [PubMed]
- Sonnex, C. Influence of ovarian hormones on urogenital infection. Sex. Transm. Infect. 1998, 74, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Pennock, J.W.; Stegall, R.; Bell, B.; Vargas, G.; Motamedi, M.; Milligan, G.; Bourne, N. Estradiol improves genital herpes vaccine efficacy in mice. Vaccine 2009, 27, 5830–5836. [Google Scholar] [CrossRef] [PubMed]
- Kaushic, C.; Ashkar, A.A.; Reid, L.A.; Rosenthal, K.L. Progesterone increases susceptibility and decreases immune responses to genital herpes infection. J. Virol. 2003, 77, 4558–4565. [Google Scholar] [CrossRef]
- Gillgrass, A.E.; Tang, V.A.; Towarnicki, K.M.; Rosenthal, K.L.; Kaushic, C. Protection against genital herpes infection in mice immunized under different hormonal conditions correlates with induction of vagina-associated lymphoid tissue. J. Virol. 2005, 79, 3117–3126. [Google Scholar] [CrossRef] [PubMed]
- Bhavanam, S.; Snider, D.P.; Kaushic, C. Intranasal and subcutaneous immunization under the effect of estradiol leads to better protection against genital HSV-2 challenge compared to progesterone. Vaccine 2008, 26, 6165–6172. [Google Scholar] [CrossRef] [PubMed]
- Gillgrass, A.; Chege, D.; Bhavanam, S.; Kaushic, C. Original article: Estradiol limits viral replication following intravaginal immunization leading to diminished mucosal IgG response and non-sterile protection against genital herpes challenge. Am. J. Reprod. Immunol. 2010, 63, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Gillgrass, A.; Ashkar, A.; Rosenthal, K.; Kaushic, C. Prolonged exposure to progesterone prevents induction of protective mucosal responses following intravaginal immunization with attenuated herpes simplex virus type 2. J. Virol. 2003, 77, 9845–9851. [Google Scholar] [CrossRef] [PubMed]
- Anipindi, V.C.; Bagri, P.; Roth, K.; Dizzell, S.E.; Nguyen, P.V.; Shaler, C.R.; Chu, D.K.; Jiménez-Saiz, R.; Liang, H.; Swift, S.; et al. Estradiol Enhances CD4+ T-Cell Anti-Viral Immunity by Priming Vaginal DCs to Induce Th17 Responses via an IL-1-Dependent Pathway. PLoS Pathog. 2016, 12, e1005589. [Google Scholar]
- United Nations. World Contraceptive Use 2012; United Nations Reports; United Nations: New York, NY, USA, 2012. [Google Scholar]
- Ferreira, V.H.; Dizzell, S.; Nazli, A.; Kafka, J.K.; Mueller, K.; Nguyen, P.V.; Tremblay, M.J.; Cochrane, A.; Kaushic, C. Medroxyprogesterone acetate regulates HIV-1 uptake and transcytosis but not replication in primary genital epithelial cells, resulting in enhanced T-cell infection. J. Infect. Dis. 2014, 211, 1745–1756. [Google Scholar] [CrossRef] [PubMed]
- Fichorova, R.N.; Desai, P.J.; Gibson, F.C.; Genco, C.A. Distinct Proinflammatory Host Responses to Neisseria Gonorrhoeae Infection in Immortalized Human Cervical and Vaginal Epithelial Cells. Infect. Immun. 2001, 69, 5840–5848. [Google Scholar] [CrossRef] [PubMed]
- Marrs, C.N.; Knobel, S.M.; Zhu, W.Q.; Sweet, S.D.; Chaudhry, A.R.; Alcendor, D.J. Evidence for Gardnerella vaginalis uptake and internalization by squamous vaginal epithelial cells: Implications for the pathogenesis of bacterial vaginosis. Microb. Infect. 2012, 14, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Africander, D.; Louw, R.; Verhoog, N.; Noeth, D.; Hapgood, J.P. Differential regulation of endogenous pro-inflammatory cytokine genes by medroxyprogesterone acetate and norethisterone acetate in cell lines of the female genital tract. Contraception 2011, 84, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Mossop, H.; Linhares, I.M.; Bongiovanni, A.M.; Ledger, W.J.; Witkin, S.S. Influence of lactic acid on endogenous and viral RNA-induced immune mediator production by vaginal epithelial cells. Obstet. Gynecol. 2011, 118, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Anukam, K.C.; Reid, G. In vitro evaluation of the viability of vaginal cells (Vk2/E6E7) and probiotic lactobacillus species in lemon juice. Sex. Health 2009, 6, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Kinlock, B.L.; Wang, Y.; Turner, T.M.; Wang, C.; Liu, B. Transcytosis of HIV-1 through vaginal epithelial cells is dependent on trafficking to the endocytic recycling pathway. PLoS ONE 2014, 9, e96760. [Google Scholar] [CrossRef]
- Hillier, S.L.; Krohn, M.A.; Rabe, L.K.; Klebanoff, S.J.; Eschenbach, D.A. The normal vaginal flora, H2O2-producing lactobacilli, and bacterial vaginosis in pregnant women. Clin. Infect. Dis. 1993, 16, S273–S281. [Google Scholar] [CrossRef] [PubMed]
- Hjelm, B.E.; Berta, A.N.; Nickerson, C.A.; Arntzen, C.J.; Herbst-Kralovetz, M.M. Development and characterization of a three-dimensional organotypic human vaginal epithelial cell model. Biol. Reprod. 2010, 82, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Miessen, K.; Einspanier, R.; Schoen, J. Establishment and characterization of a differentiated epithelial cell culture model derived from the porcine cervix uteri. BMC Vet. Res. 2012, 8, 31. [Google Scholar] [CrossRef] [PubMed]
- Canny, G.; Trifonova, R.; Kindelberger, D.; Colgan, S.; Fichorova, R. Expression and function of bactericidal/permeability-increasing protein in human genital tract epithelial cells. J. Infect. Dis. 2006, 194, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Fichorova, R.; Zhou, F.; Ratnam, V.; Atanassova, V.; Jiang, S.; Strick, N.; Neurath, A. Anti-human immunodeficiency virus type 1 microbicide cellulose acetate 1,2-benzenedicarboxylate in a human in vitro model of vaginal inflammation. Antimicrob. Agents Chemother. 2004, 49, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Pezzulo, A.A.; Starner, T.D.; Scheetz, T.E.; Traver, G.L.; Tilley, A.E.; Harvey, B.-G.; Crystal, R.G.; McCray, P.B.; Zabner, J. The air-liquid interface and use of primary cell cultures are important to recapitulate the transcriptional profile of in vivo airway epithelia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2011, 300, L25–L31. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Li, H.; Cho, H.-J.; Bian, S.; Roh, H.-J.; Lee, M.-K.; Kim, J.S.; Chung, S.-J.; Shim, C.-K.; Kim, D.-D. Air-Liquid interface (ALI) culture of human bronchial epithelial cell monolayers as an in vitro model for airway drug transport studies. J. Pharm. Sci. 2007, 96, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.P.; Bear, C.E.; Chin, S.; Pasceri, P.; Thompson, T.O.; Huan, L.-J.; Ratjen, F.; Ellis, J.; Rossant, J. Directed differentiation of human pluripotent stem cells into mature airway epithelia expressing functional CFTR protein. Nat. Biotechnol. 2012, 30, 876–882. [Google Scholar] [CrossRef] [PubMed]
- Fichorova, R.N.; Rheinwald, J.G.; Anderson, D.J. Generation of papillomavirus-immortalized cell lines from normal human ectocervical, endocervical, and vaginal epithelium that maintain expression of tissue-specific differentiation proteins. Biol. Reprod. 1997, 57, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Eade, C.; Diaz, C.; Wood, M.; Anastos, K.; Patterson, B.; Gupta, P.; Cole, A.; Cole, A. Identification and characterization of bacterial vaginosis-associated pathogens using a comprehensive cervical-vaginal epithelial coculture assay. PLoS ONE 2012, 7, e50106. [Google Scholar] [CrossRef] [PubMed]
- Fichorova, R.; Yamamoto, H.; Delaney, M.; Onderdonk, A.; Doncel, G. Novel vaginal microflora colonization model providing new insight into microbicide mechanism of action. mBio 2011, 2, e00168-11. [Google Scholar] [CrossRef] [PubMed]
- Fichorova, R.; Anderson, D.J. Differential expression of immunobiological mediators by immortalized human cervical and vaginal epithelial cells. Biol. Reprod. 1999, 60, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Krebs, F.; Miller, S.; Catalone, B.; Fichorova, R.; Anderson, D.; Malamud, D.; Howett, M.; Wigdahl, B. Comparative in vitro sensitivities of human immune cell lines, vaginal and cervical epithelial cell lines, and primary cells to candidate microbicides nonoxynol 9, C31G, and sodium dodecyl sulfate. Antimicrob. Agents Chemother. 2002, 46, 2292–2298. [Google Scholar] [CrossRef] [PubMed]
- Fichorova, R.; Buck, O.; Yamamoto, H.; Fashemi, T.; Dawood, H.; Fashemi, B.; Hayes, G.; Beach, D.; Takagi, Y.; Delaney, M.; et al. The villain team-up or how Trichomonas vaginalis and bacterial vaginosis alter innate immunity in concert. Sex. Transm. Infect. 2013, 89, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Zalenskaya, I.; Joseph, T.; Bavarva, J.; Yousefieh, N.; Jackson, S.; Fashemi, T.; Yamamoto, H.; Settlage, R.; Fichorova, R.; Doncel, G. Gene expression profiling of human vaginal cells in vitro discriminates compounds with pro-inflammatory and mucosa-altering properties: Novel biomarkers for preclinical testing of HIV microbicide candidates. PLoS ONE 2015, 10, e0128557. [Google Scholar] [CrossRef] [PubMed]
- Cummins, J.; Doncel, G. Biomarkers of cervicovaginal inflammation for the assessment of microbicide safety. Sex. Transm. Dis. 2009, 36, S84–S91. [Google Scholar] [CrossRef] [PubMed]
- Fichorova, R.; Cronin, A.; Lien, E.; Anderson, D.; Ingalls, R. Response to Neisseria gonorrhoeae by cervicovaginal epithelial cells occurs in the absence of toll-like receptor 4-mediated signaling. J. Immunol. 2002, 168, 2424–2432. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Rosenthal, K. Herpes simplex virus type 2 virion host shutoff protein suppresses innate dsRNA antiviral pathways in human vaginal epithelial cells. J. Gen. Virol. 2011, 92, 1981–1993. [Google Scholar] [CrossRef] [PubMed]
- Strober, W. Trypan Blue Exclusion Test of Cell Viability. Curr. Protoc. Immunol. 2001. [Google Scholar] [CrossRef]
- Ferreira, V.; Nazli, A.; Mossman, K.; Kaushic, C. Proinflammatory cytokines and chemokines—But not interferon-β—Produced in response to HSV-2 in primary human genital epithelial cells are associated with viral replication and the presence of the virion host shutoff protein. Am. J. Reprod. Immunol. 2013, 70, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Kouklis, P.D.; Hutton, E.; Fuchs, E. Making a Connection: Direct Binding between Keratin Intermediate Filaments and Desmosomal Proteins. J. Cell Biol. 1994, 127, 1049–1060. [Google Scholar] [CrossRef] [PubMed]
- Uzman, A.; Lodish, H.; Berk, A.; Zipursky, L.; Baltimore, D. The Molecules of Life. In Molecular Cell Biology, 4th ed.; W.H. Freeman and Company: New York, NY, USA, 2000; ISBN 0-7167-3136-3. pp. 8–13. [Google Scholar]
- Osmani, N.; Labouesse, M. Remodeling of Keratin-Coupled Cell Adhesion Complexes. Curr. Opin. Cell Biol. 2015, 32, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Xu, Q.; Fichorova, R. Homeostatic properties of Lactobacillus jensenii engineered as a live vaginal anti-HIV microbicide. BMC Microbiol. 2013, 13, 4. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.; Gudas, L.J.; Katzenellenbogen, B.S. Regulation of Keratin 19 Gene Expression by Estrogen in Human Breast Cancer Cells and Identification of the Estrogen Responsive Gene Region. Mol. Cell. Endocrinol. 2000, 164, 225–237. [Google Scholar] [CrossRef]
- Basu, S.; Pioli, P.A.; Conejo-Garcia, J.; Wira, C.R.; Sentman, C.L. Estradiol Regulates MICA Expression in Human Endometrial Cells. Clin. Immunol. 2008, 129, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Patton, D.; Thwin, S.; Meier, A.; Hooton, T.; Stapleton, A.; Eschenbach, D. Epithelial cell layer thickness and immune cell populations in the normal human vagina at different stages of the menstrual cycle. Am. J. Obstet. Gynecol. 2000, 183, 967–973. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, D.L.; Kurita, T.; Taylor, J.A.; Lubahn, D.B.; Cunha, G.R.; Cooke, P.S. Role of Stromal and Epithelial Estrogen Receptors in Vaginal Epithelial Proliferation, Stratification, and Cornification. Endocrinology 1998, 139, 4345–4352. [Google Scholar] [CrossRef] [PubMed]
- Martin, H.L., Jr.; Nyange, P.M.; Richardson, B.A.; Lavreys, L.; Mandaliya, K.; Jackson, D.J.; Ndinya-Achola, J.O.; Kreiss, J. Hormonal contraception, sexually transmitted diseases, and risk of heterosexual transmission of human immunodeficiency virus type 1. J. Infect. Dis. 1998, 178, 1053–1059. [Google Scholar] [CrossRef] [PubMed]
- Marx, P.A.; Spira, A.I.; Gettie, A.; Dailey, P.J.; Veazey, R.S.; Lackner, A.A.; Mahoney, C.J.; Miller, C.J.; Claypool, L.E.; Ho, D.D.; et al. Progesterone Implants Enhance SIV Vaginal Transmission and Early Virus Load. Nat. Med. 1996, 2, 1084–1089. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Forney, L.J.; Ravel, J. Vaginal Microbiome: Rethinking Health and Disease. Annu. Rev. Microbiol. 2012, 66, 371–389. [Google Scholar] [CrossRef] [PubMed]





© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.; Dizzell, S.E.; Leung, V.; Nazli, A.; Zahoor, M.A.; Fichorova, R.N.; Kaushic, C. Effects of Female Sex Hormones on Susceptibility to HSV-2 in Vaginal Cells Grown in Air-Liquid Interface. Viruses 2016, 8, 241. https://doi.org/10.3390/v8090241
Lee Y, Dizzell SE, Leung V, Nazli A, Zahoor MA, Fichorova RN, Kaushic C. Effects of Female Sex Hormones on Susceptibility to HSV-2 in Vaginal Cells Grown in Air-Liquid Interface. Viruses. 2016; 8(9):241. https://doi.org/10.3390/v8090241
Chicago/Turabian StyleLee, Yung, Sara E. Dizzell, Vivian Leung, Aisha Nazli, Muhammad A. Zahoor, Raina N. Fichorova, and Charu Kaushic. 2016. "Effects of Female Sex Hormones on Susceptibility to HSV-2 in Vaginal Cells Grown in Air-Liquid Interface" Viruses 8, no. 9: 241. https://doi.org/10.3390/v8090241
APA StyleLee, Y., Dizzell, S. E., Leung, V., Nazli, A., Zahoor, M. A., Fichorova, R. N., & Kaushic, C. (2016). Effects of Female Sex Hormones on Susceptibility to HSV-2 in Vaginal Cells Grown in Air-Liquid Interface. Viruses, 8(9), 241. https://doi.org/10.3390/v8090241