
PreC and C Regions of Woodchuck Hepatitis Virus Facilitate Persistent Expression of Surface Antigen of Chimeric WHV-HBV Virus in the Hydrodynamic Injection BALB/c Mouse Model

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
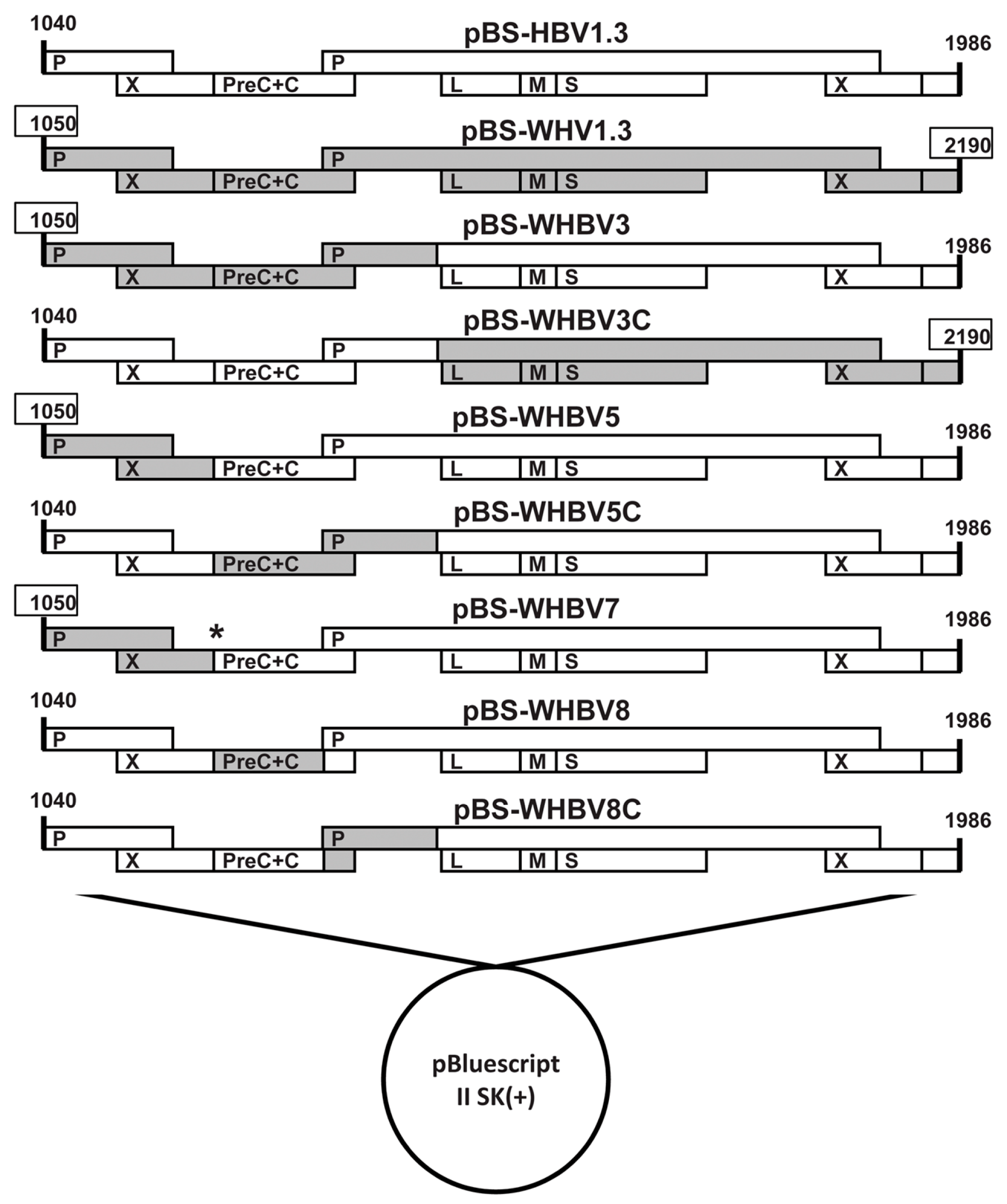
2.2. Chimeric Woodchuck Hepatitis Virus and Hepatitis B Virus (WHV-HBV) Genomes
2.3. Hydrodynamic Injection (HI) of BALB/c Mice
2.4. Serological Assays
2.5. Measurementof Serum Viral DNA (vDNA)
2.6. Measurementof WHV Core Antigen (WHcAg) and HBV Core Antigen (HBcAg) via Immunohistochemical Staining (IHC)
2.7. Measurement of Cytotoxic T Lymphocyte (CTL) Responses
2.8. Statistical Analysis
3. Results
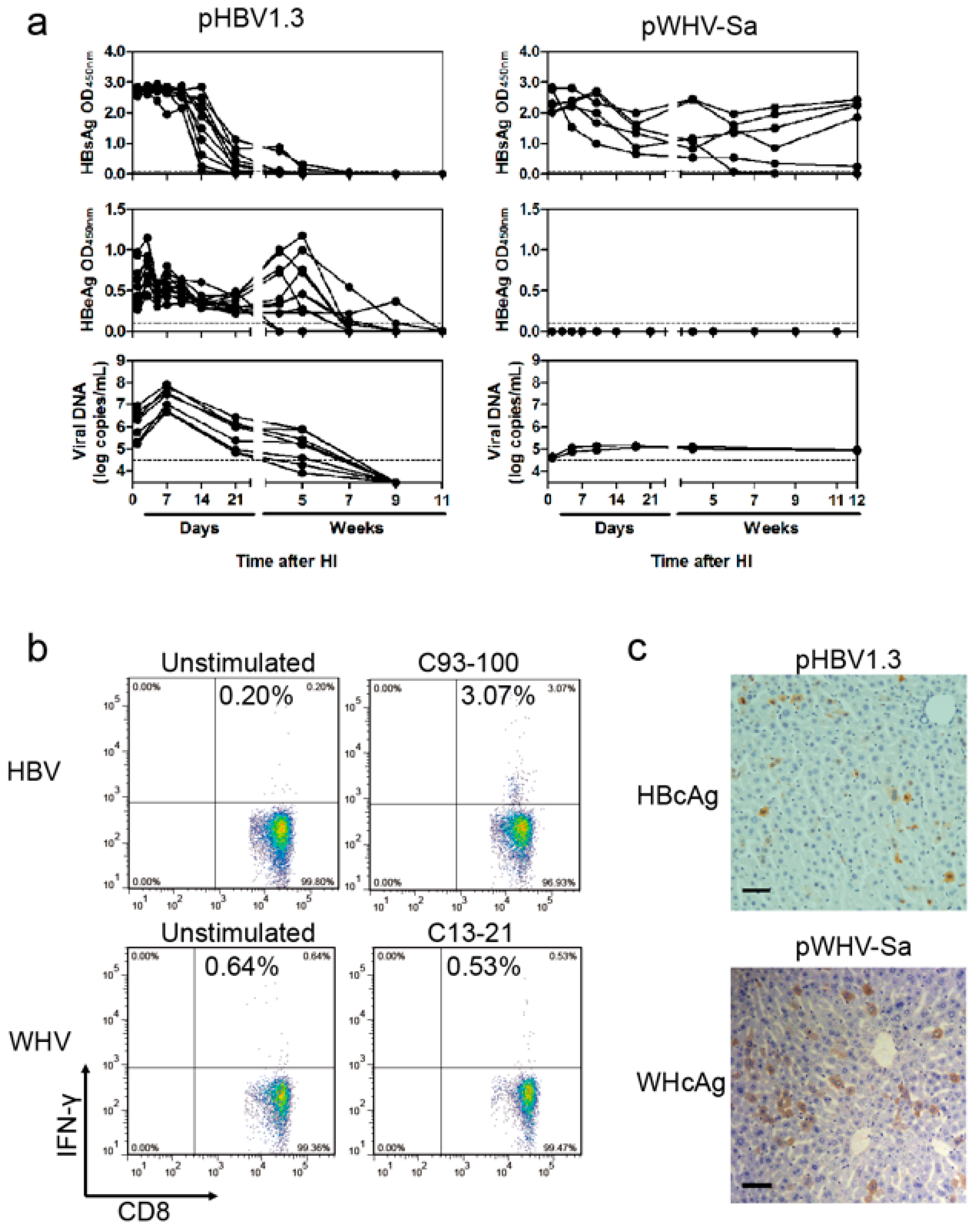
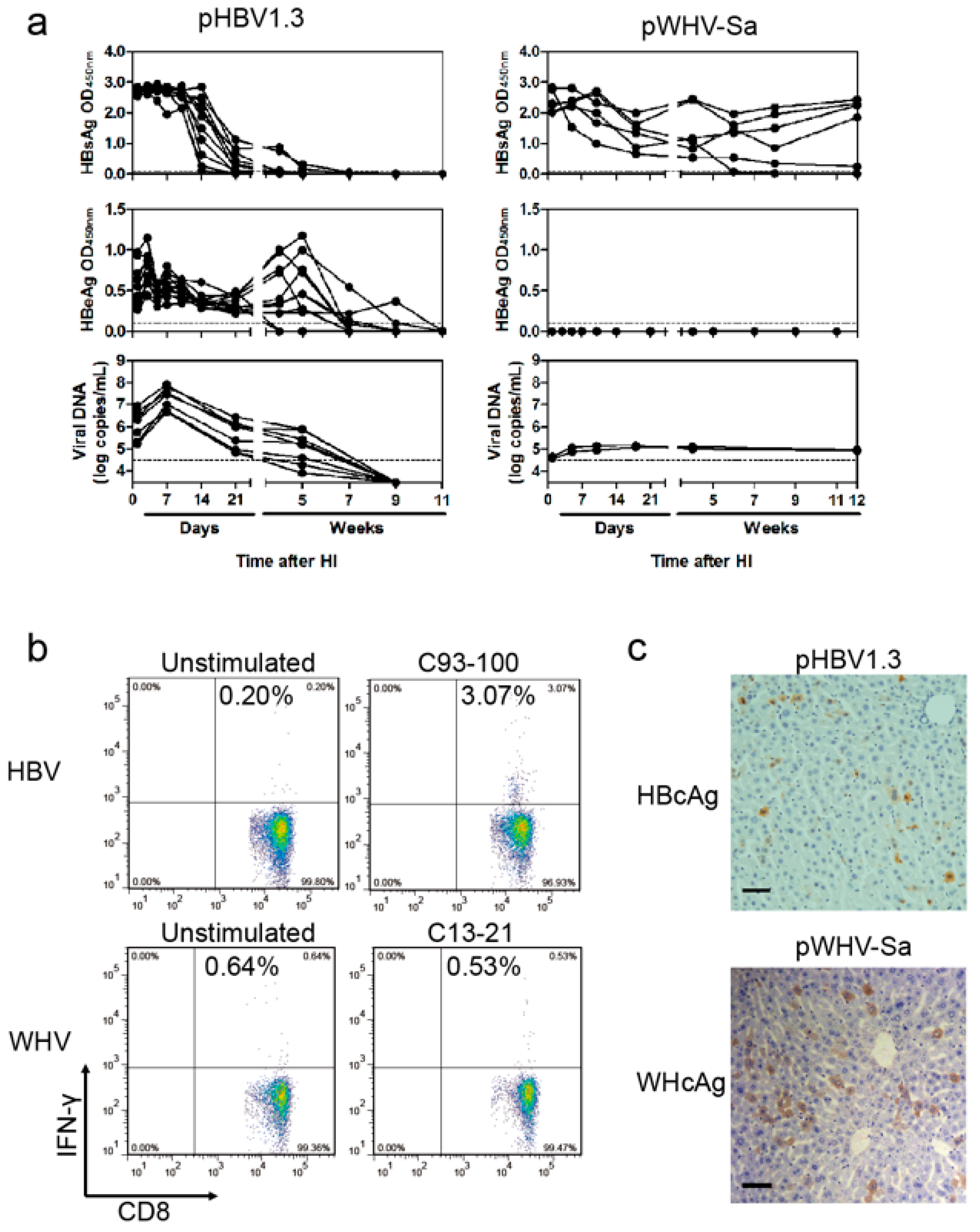
3.1. WHV Could Persist in BALB/c Mice after HI
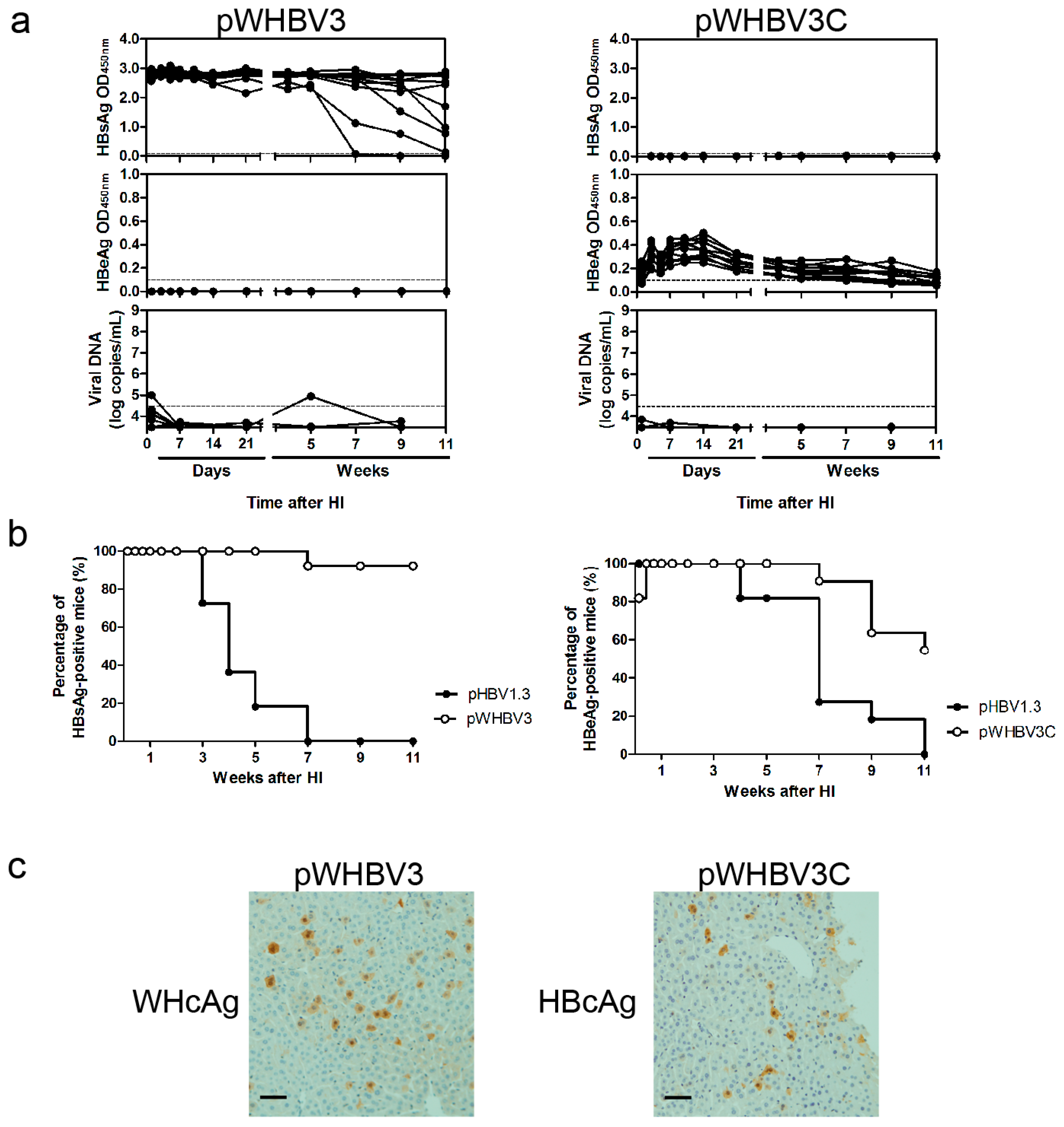
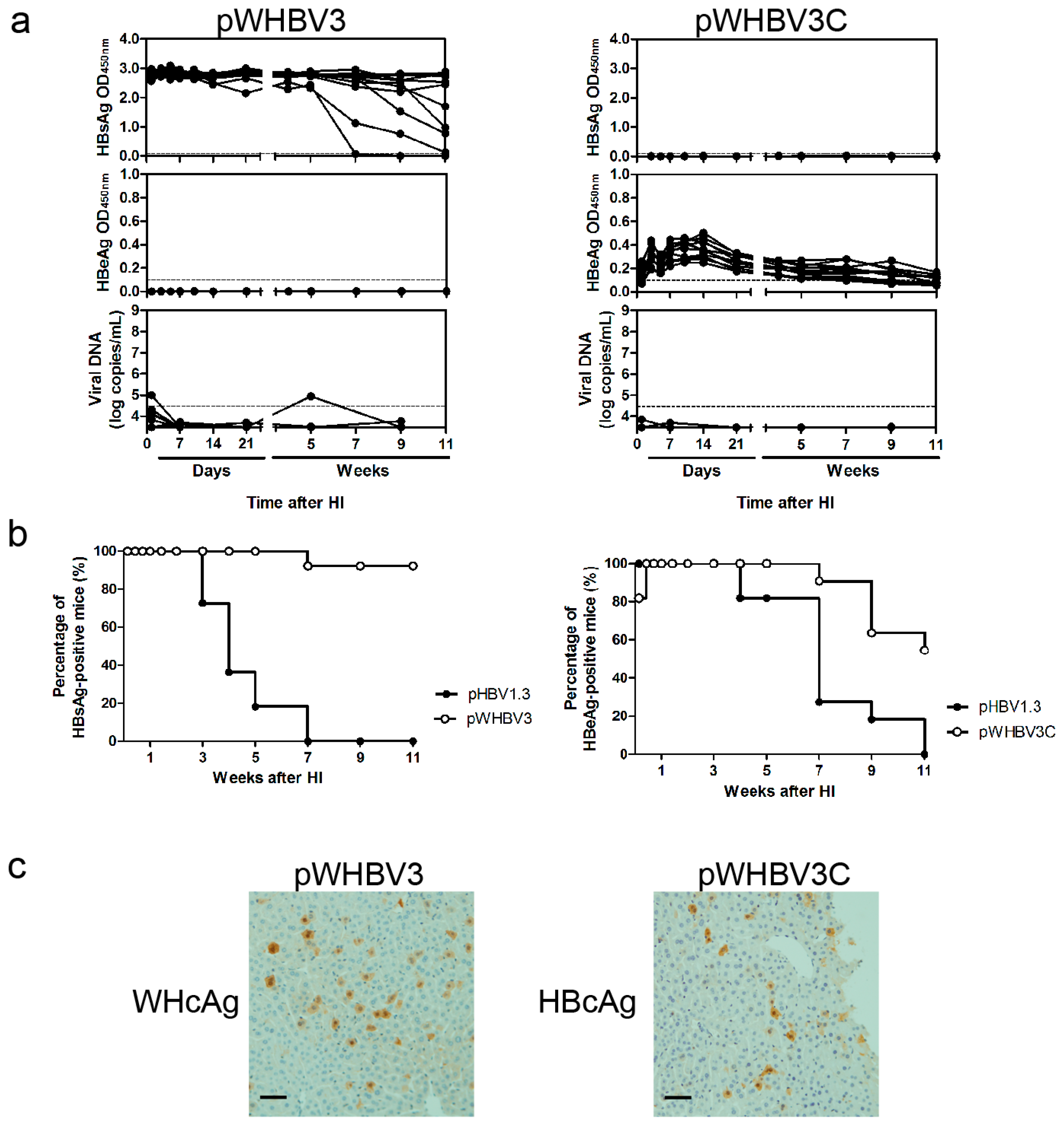
3.2. Persistence of Chimeric WHV-HBV Genome of pWHBV3 in BALB/c Mice
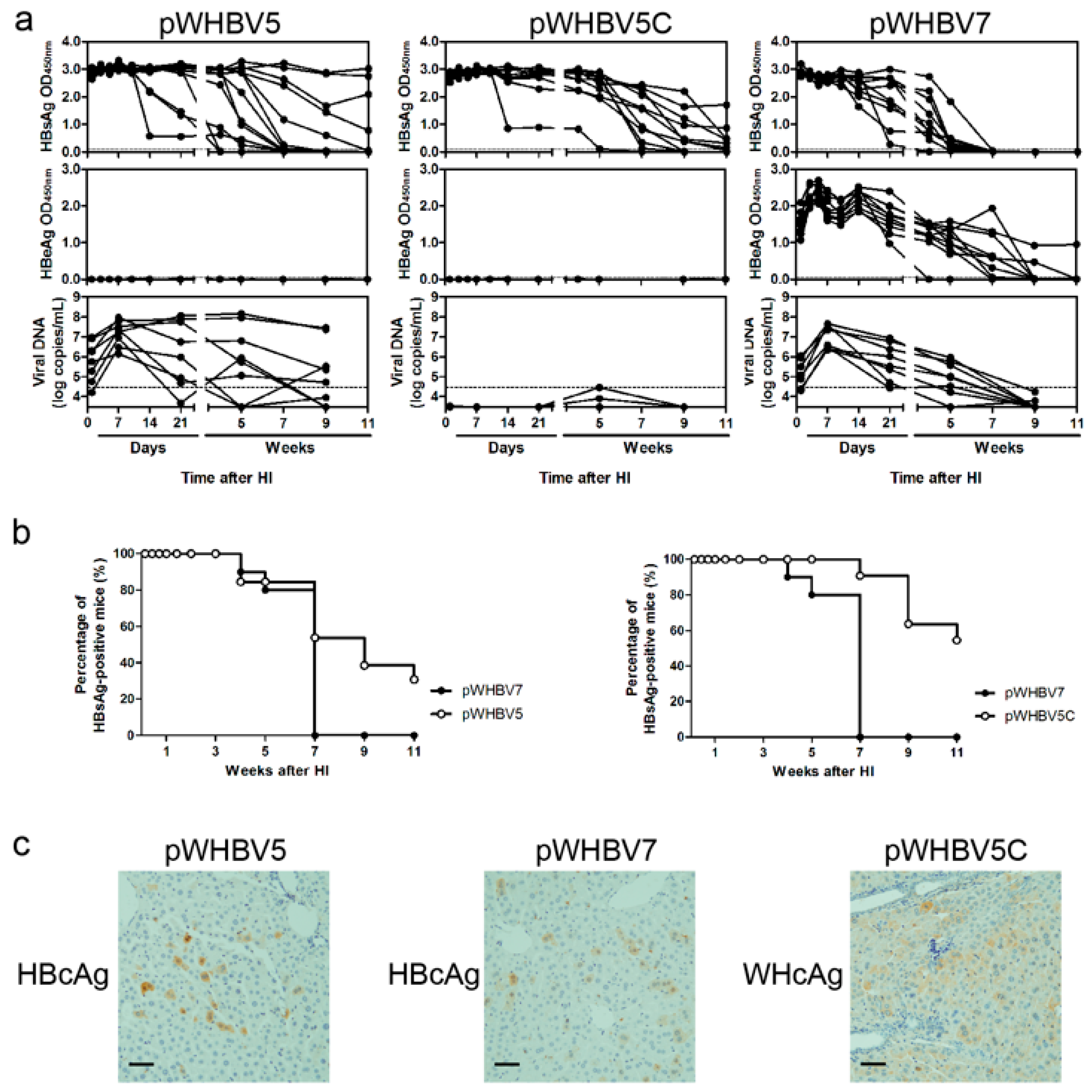
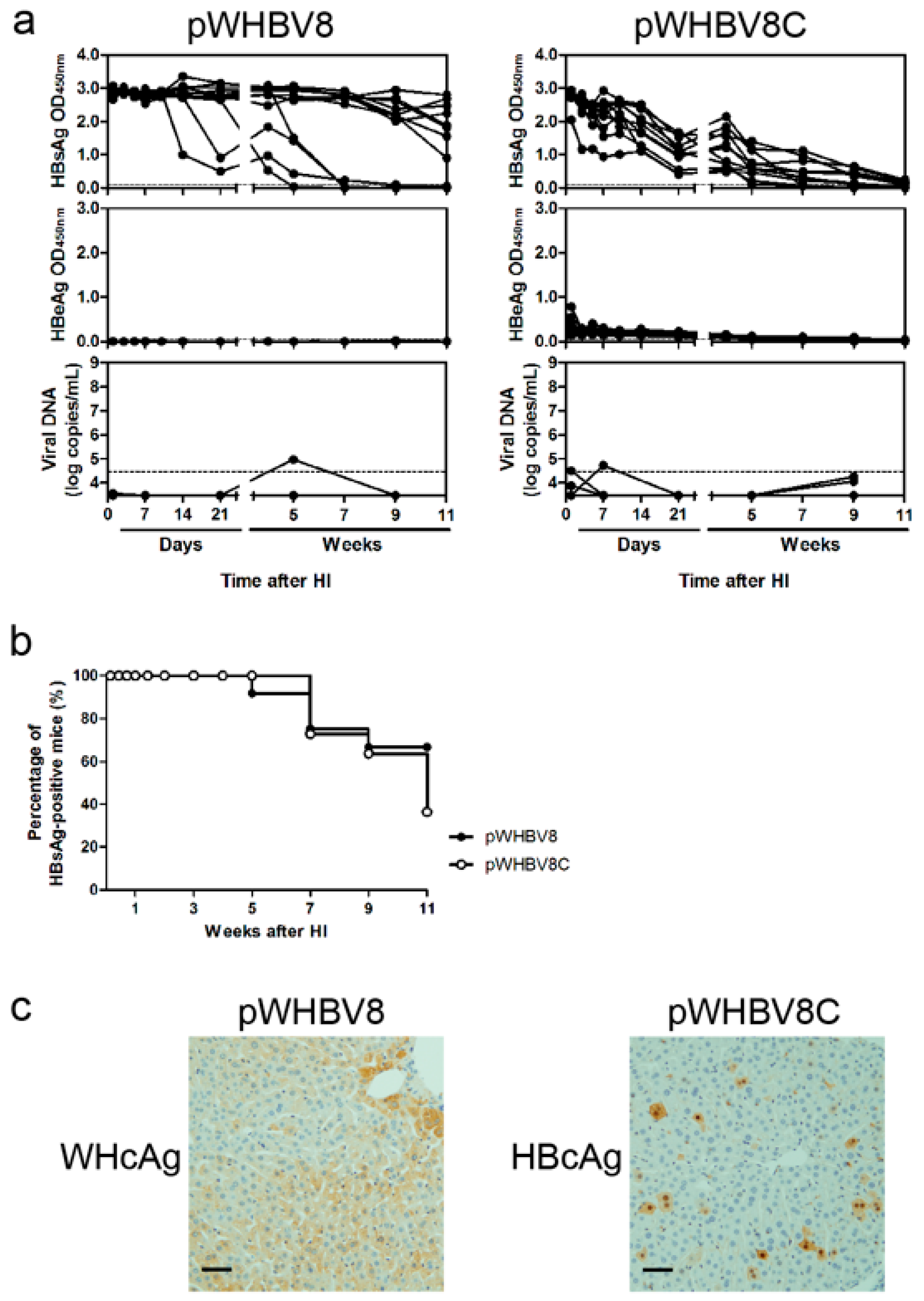
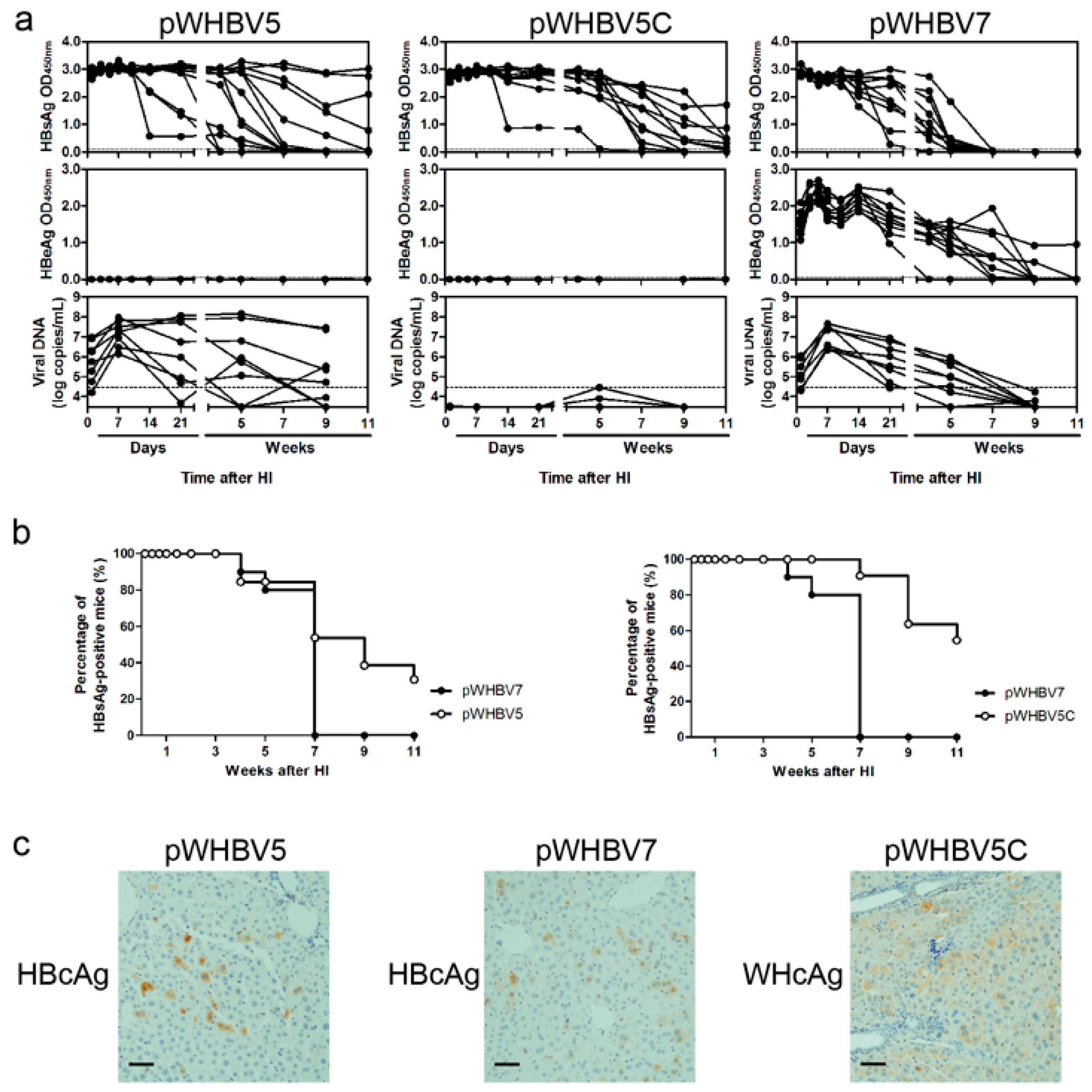
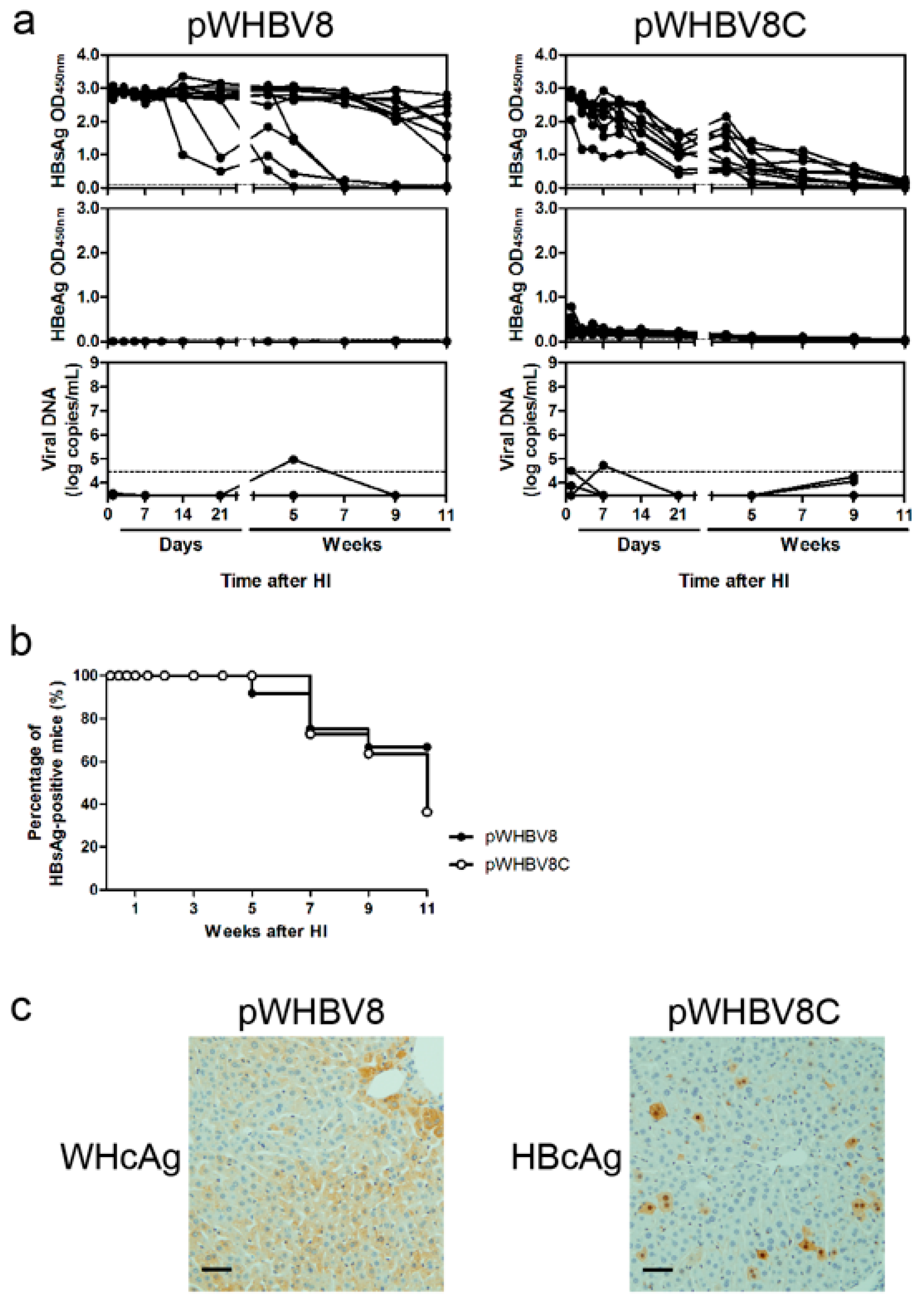
3.3. Persistent HBsAg Antigenemia after HI with Chimeric WHV-HBV Genomes of pWHBV5C and pWHBV8
3.4. C1819T Mutation in pWHBV5 Led to HBsAg Persistence
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yang, P.L.; Althage, A.; Chung, J.; Chisari, F.V. Hydrodynamic injection of viral DNA: A mouse model of acute hepatitis b virus infection. Proc. Natl. Acad. Sci. USA 2002, 99, 13825–13830. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Wu, C.; Shi, H.; Gong, Z.; Zhang, E.; Wang, H.; Zhao, K.; Liu, S.; Li, S.; Gao, X.; et al. Coexistence of hepatitis b virus quasispecies enhances viral replication and the ability to induce host antibody and cellular immune responses. J. Virol. 2014, 88, 8656–8666. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.R.; Wu, H.L.; Chen, P.J.; Chen, D.S. An immunocompetent mouse model for the tolerance of human chronic hepatitis b virus infection. Proc. Natl. Acad. Sci. USA 2006, 103, 17862–17867. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.J.; Huang, L.R.; Yang, H.C.; Tzeng, H.T.; Hsu, P.N.; Wu, H.L.; Chen, P.J.; Chen, D.S. Hepatitis b virus core antigen determines viral persistence in a c57bl/6 mouse model. Proc. Natl. Acad. Sci. USA 2010, 107, 9340–9345. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Chen, W.L.; Ou, J.H. Effects of interferon-alpha/beta on hbv replication determined by viral load. PLoS Pathog. 2011, 7, e1002159. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Wu, C.; Song, J.; Wang, J.; Zhang, E.; Liu, H.; Yang, D.; Chen, X.; Lu, M.; Xu, Y. DNA immunization with fusion of ctla-4 to hepatitis b virus (hbv) core protein enhanced th2 type responses and cleared hbv with an accelerated kinetic. PLoS ONE 2011, 6, e22524. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Deng, W.; Deng, L.; Cao, L.; Qin, B.; Li, S.; Wang, Y.; Pei, R.; Yang, D.; Lu, M.; et al. Amino acid substitutions at positions 122 and 145 of hepatitis b virus surface antigen (hbsag) determine the antigenicity and immunogenicity of hbsag and influence in vivo hbsag clearance. J. Virol. 2012, 86, 4658–4669. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Zhou, Y.; Li, S.; Wang, B.; Zheng, X.; Wu, J.; Gibbert, K.; Dittmer, U.; Lu, M.; Yang, D. Susceptibility of different hepatitis b virus isolates to interferon-alpha in a mouse model based on hydrodynamic injection. PLoS ONE 2014, 9, e90977. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.L.; Althage, A.; Chung, J.; Maier, H.; Wieland, S.; Isogawa, M.; Chisari, F.V. Immune effectors required for hepatitis b virus clearance. Proc. Natl. Acad. Sci. USA 2010, 107, 798–802. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, B.; Huang, S.; Song, Z.; Wu, J.; Zhang, E.; Zhu, Z.; Zhu, B.; Yin, Y.; Lin, Y.; et al. Immunosuppressive drugs modulate the replication of hepatitis b virus (hbv) in a hydrodynamic injection mouse model. PLoS ONE 2014, 9, e85832. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Budeus, B.; Cao, L.; Wu, C.; Wang, Y.; Zhang, X.; Rayner, S.; Hoffmann, D.; Lu, M.; Chen, X. The amino acid substitutions rtp177g and rtf249a in the reverse transcriptase domain of hepatitis b virus polymerase reduce the susceptibility to tenofovir. Antiviral Res. 2013, 97, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Menne, S.; Cote, P.J. The woodchuck as an animal model for pathogenesis and therapy of chronic hepatitis b virus infection. World J. Gastroenterol. 2007, 13, 104–124. [Google Scholar] [CrossRef] [PubMed]
- Galibert, F.; Chen, T.N.; Mandart, E. Nucleotide sequence of a cloned woodchuck hepatitis virus genome: Comparison with the hepatitis b virus sequence. J. Virol. 1982, 41, 51–65. [Google Scholar] [PubMed]
- Ziermann, R.; Ganem, D. Homologous and heterologous complementation of hbv and whv capsid and polymerase functions in rna encapsidation. Virology 1996, 219, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, H.; Omi, S.; Wang, Y.; Imai, M.; Mayumi, M. Trans-Complementation of the c gene of human and the p gene of woodchuck hepadnaviruses. J. Gen. Virol. 1990, 71 (Pt. 4), 959–963. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, S.; Glebe, D.; Wend, U.C.; Oyunbileg, J.; Gerlich, W.H. Universal primers for real-time amplification of DNA from all known orthohepadnavirus species. J. Clin. Virol. 2003, 27, 30–37. [Google Scholar] [CrossRef]
- Wang, B.J.; Tian, Y.J.; Meng, Z.J.; Jiang, M.; Wei, B.Q.; Tao, Y.Q.; Fan, W.; Li, A.Y.; Bao, J.J.; Li, X.Y.; et al. Establishing a new animal model for hepadnaviral infection: Susceptibility of chinese marmota-species to woodchuck hepatitis virus infection. J. Gen. Virol. 2011, 92, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Kosinska, A.D.; Johrden, L.; Zhang, E.; Fiedler, M.; Mayer, A.; Wildner, O.; Lu, M.; Roggendorf, M. DNA prime-adenovirus boost immunization induces a vigorous and multifunctional t-cell response against hepadnaviral proteins in the mouse and woodchuck model. J. Virol. 2012, 86, 9297–9310. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Lin, Y.; Wu, W.; Song, J.; Zhang, E.; Wu, C.; Chen, X.; Hu, K.; Yang, D.; Xu, Y.; et al. Persistence of the recombinant genomes of woodchuck hepatitis virus in the mouse model. PLoS ONE 2015, 10, e0125658. [Google Scholar] [CrossRef] [PubMed]
- Lucifora, J.; Xia, Y.; Reisinger, F.; Zhang, K.; Stadler, D.; Cheng, X.; Sprinzl, M.F.; Koppensteiner, H.; Makowska, Z.; Volz, T.; et al. Specific and nonhepatotoxic degradation of nuclear hepatitis b virus cccdna. Science 2014, 343, 1221–1228. [Google Scholar] [CrossRef] [PubMed]
- Nassal, M. The arginine-rich domain of the hepatitis b virus core protein is required for pregenome encapsidation and productive viral positive-strand DNA synthesis but not for virus assembly. J. Virol. 1992, 66, 4107–4116. [Google Scholar] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, W.; Liu, Y.; Lin, Y.; Pan, D.; Yang, D.; Lu, M.; Xu, Y. PreC and C Regions of Woodchuck Hepatitis Virus Facilitate Persistent Expression of Surface Antigen of Chimeric WHV-HBV Virus in the Hydrodynamic Injection BALB/c Mouse Model. Viruses 2017, 9, 35. https://doi.org/10.3390/v9020035
Wu W, Liu Y, Lin Y, Pan D, Yang D, Lu M, Xu Y. PreC and C Regions of Woodchuck Hepatitis Virus Facilitate Persistent Expression of Surface Antigen of Chimeric WHV-HBV Virus in the Hydrodynamic Injection BALB/c Mouse Model. Viruses. 2017; 9(2):35. https://doi.org/10.3390/v9020035
Chicago/Turabian StyleWu, Weimin, Yan Liu, Yong Lin, Danzhen Pan, Dongliang Yang, Mengji Lu, and Yang Xu. 2017. "PreC and C Regions of Woodchuck Hepatitis Virus Facilitate Persistent Expression of Surface Antigen of Chimeric WHV-HBV Virus in the Hydrodynamic Injection BALB/c Mouse Model" Viruses 9, no. 2: 35. https://doi.org/10.3390/v9020035
APA StyleWu, W., Liu, Y., Lin, Y., Pan, D., Yang, D., Lu, M., & Xu, Y. (2017). PreC and C Regions of Woodchuck Hepatitis Virus Facilitate Persistent Expression of Surface Antigen of Chimeric WHV-HBV Virus in the Hydrodynamic Injection BALB/c Mouse Model. Viruses, 9(2), 35. https://doi.org/10.3390/v9020035