
Evaluation of Rice Resistance to Southern Rice Black-Streaked Dwarf Virus and Rice Ragged Stunt Virus through Combined Field Tests, Quantitative Real-Time PCR, and Proteome Analysis

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Tests
2.2. Quantification of SRBSDV and RRSV Gene Expression Levels by qRT-PCR
2.3. Quantitative Label-Free Shotgun Proteomic Analysis of Three Rice Varieties
3. Results
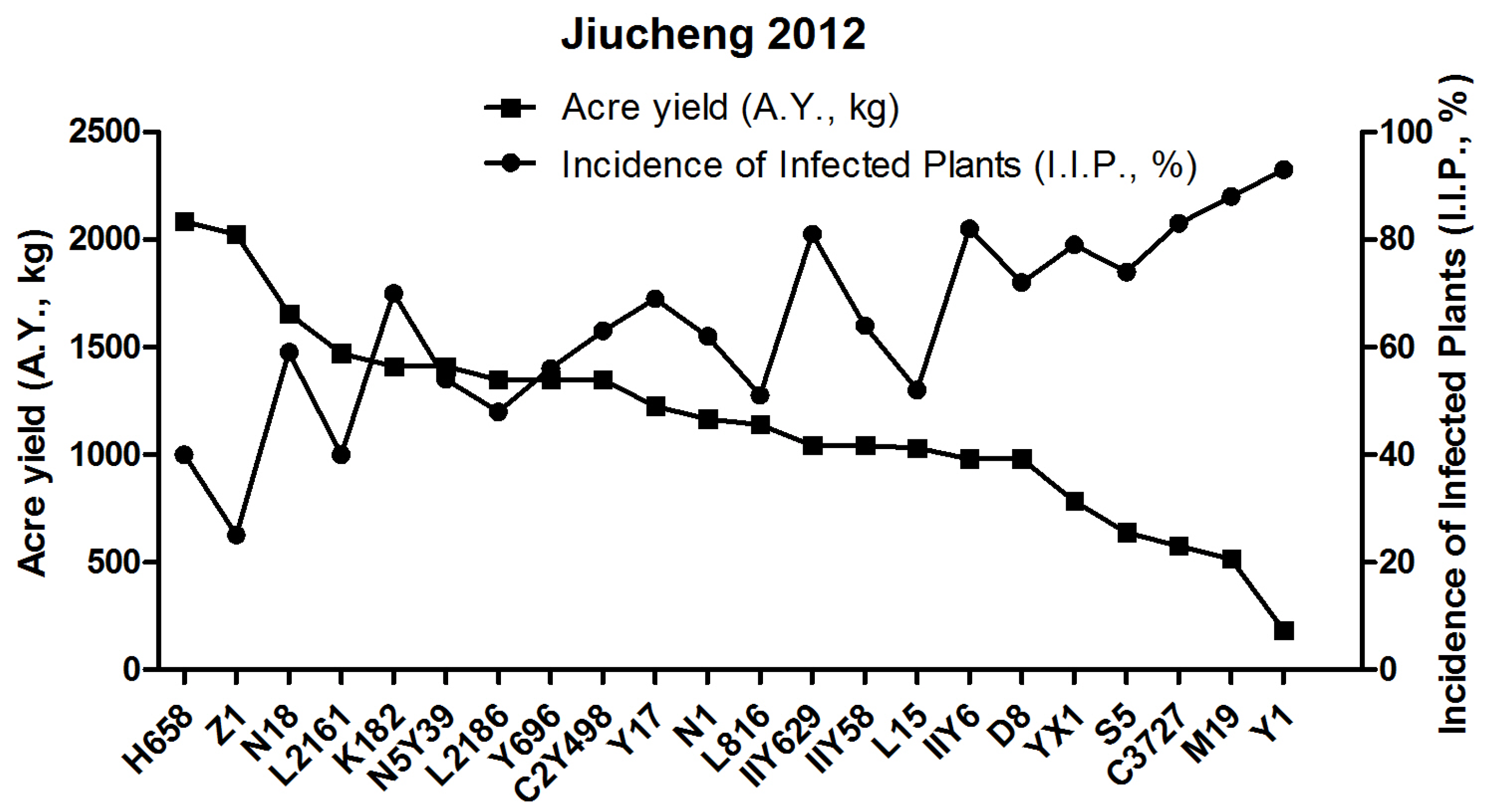
3.1. Field Yields Test of 22 Rice Candidates in Jiucheng in 2012
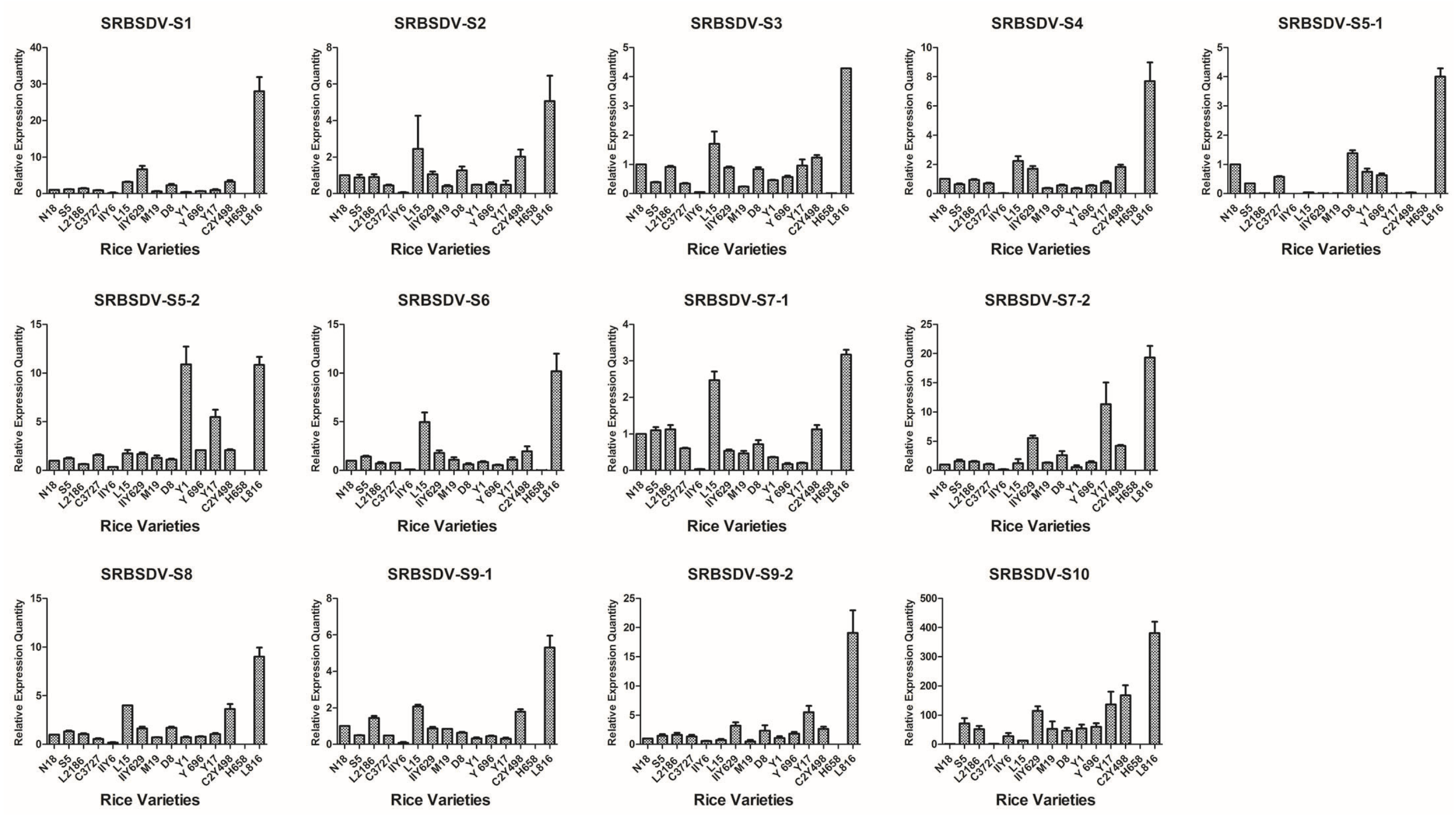
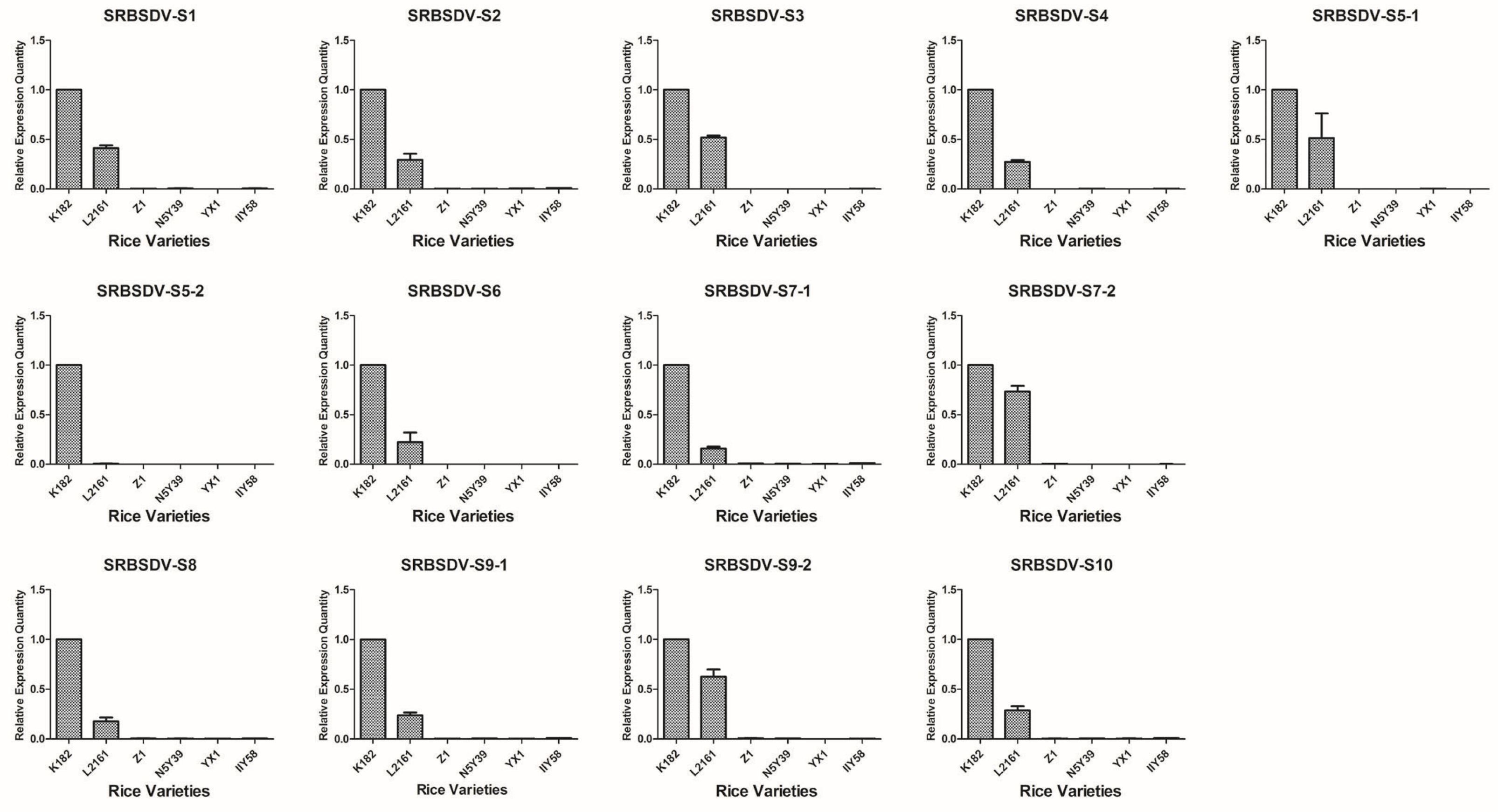
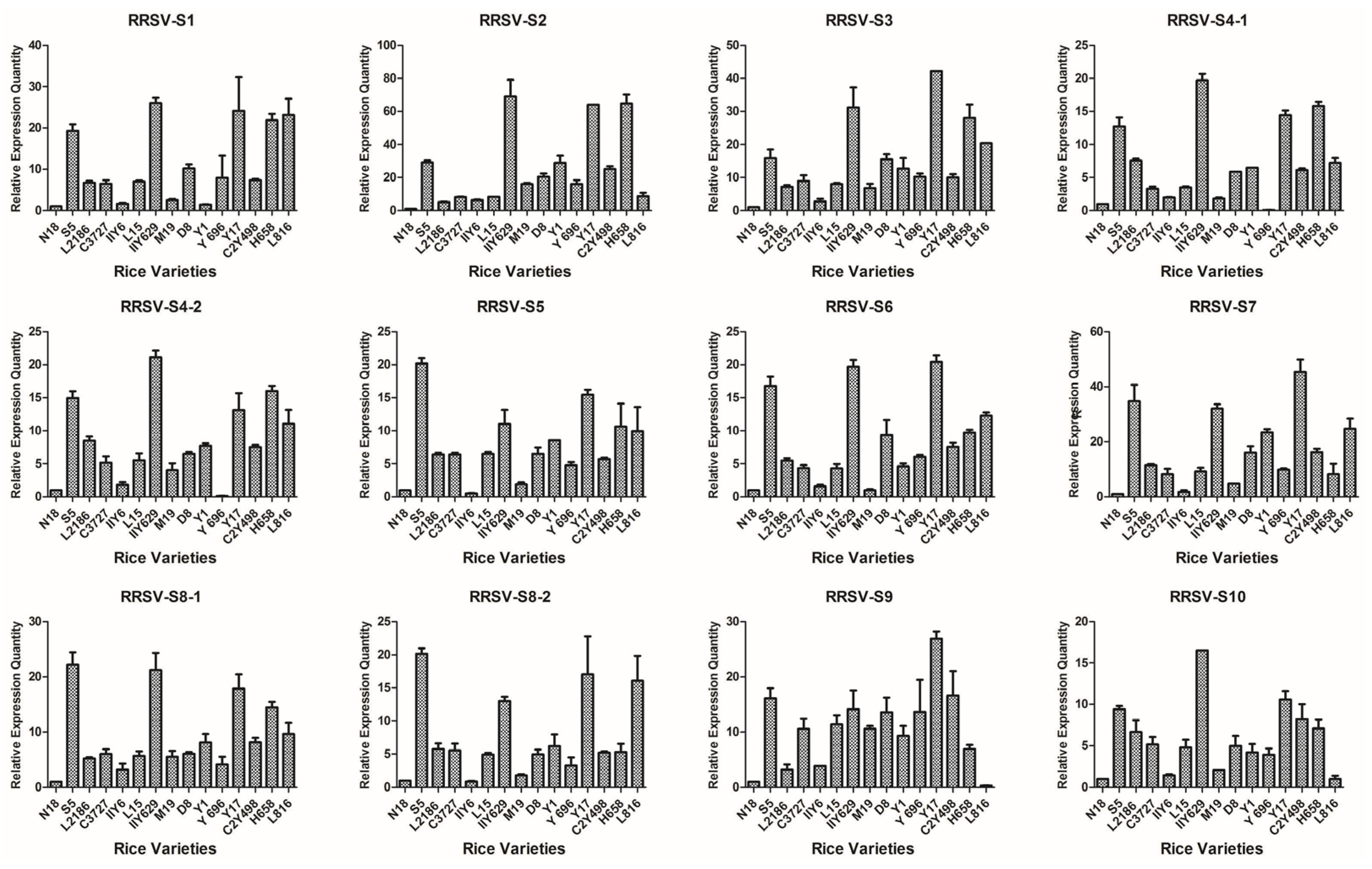
3.2. Analysis of the Relative mRNA Expression Level of SRBSDV and RRSV Genes
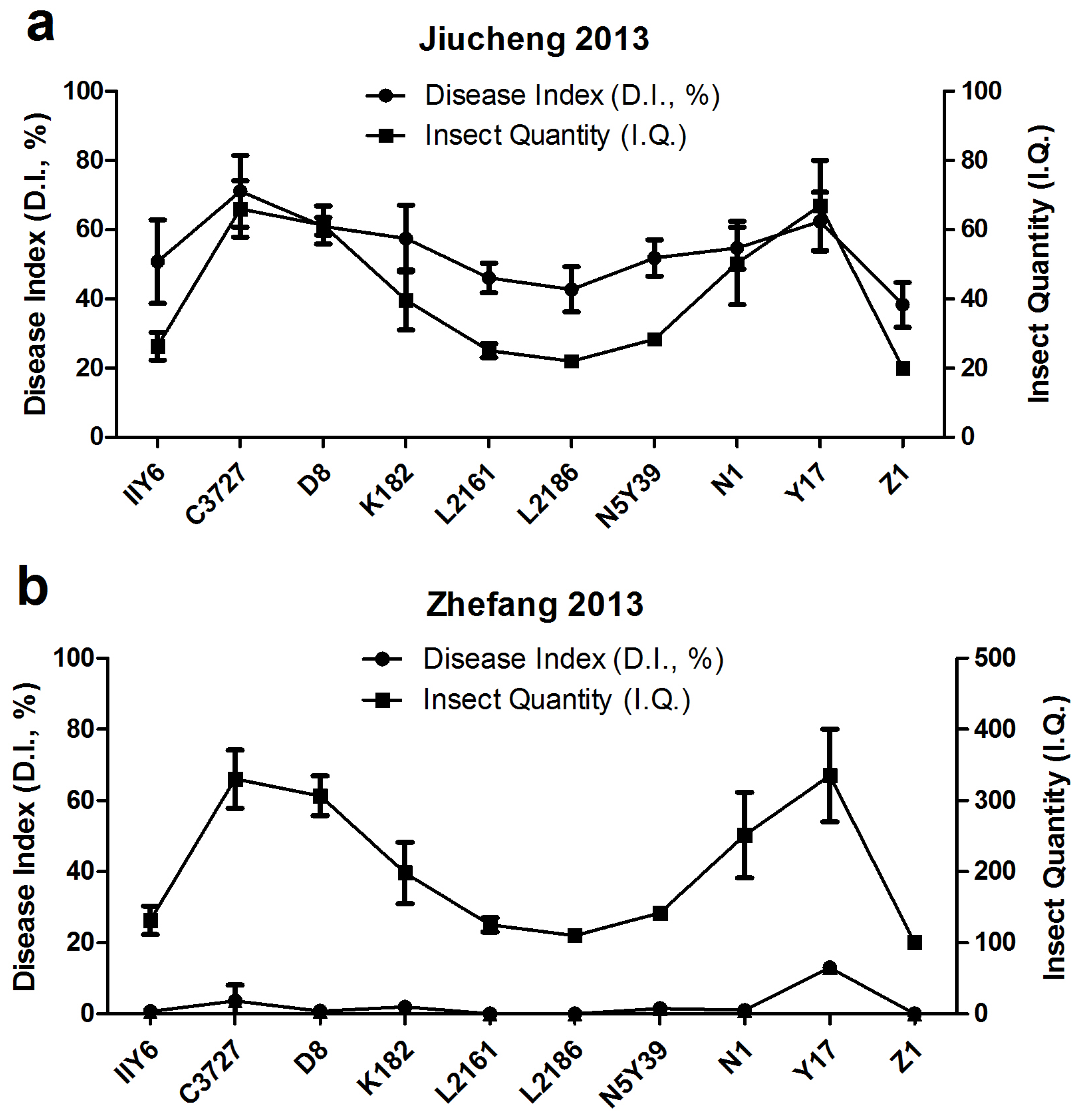
3.3. Field Yields Tests of 10 Major Rice Cultivars in Jiucheng and Zhefang in 2013
3.4. Proteomic Analysis in Three Healthy and SRBSDV-Stressed Rice Cultivars
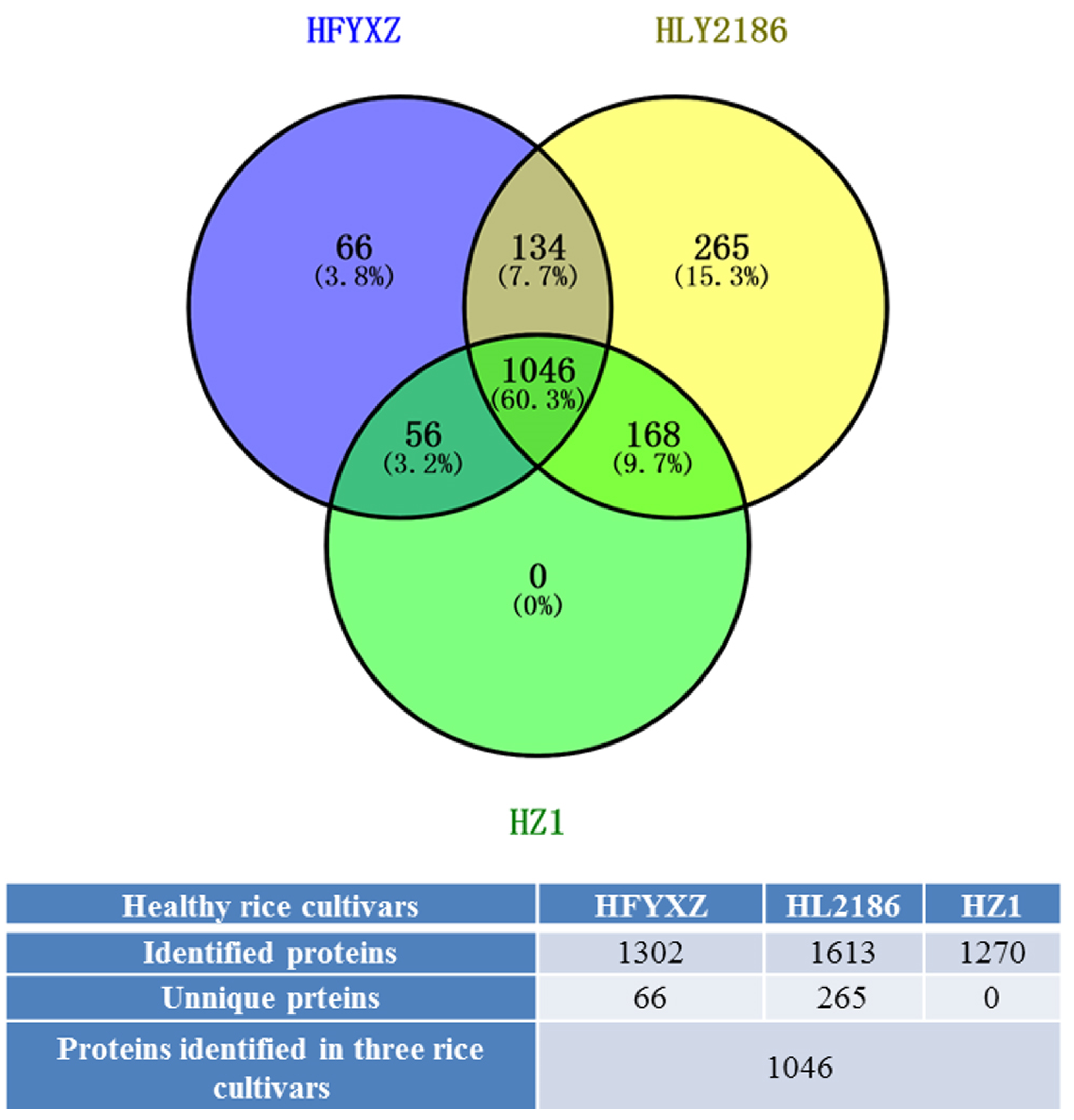
3.4.1. Overview of the Proteomic Analysis of Three Healthy Cultivars
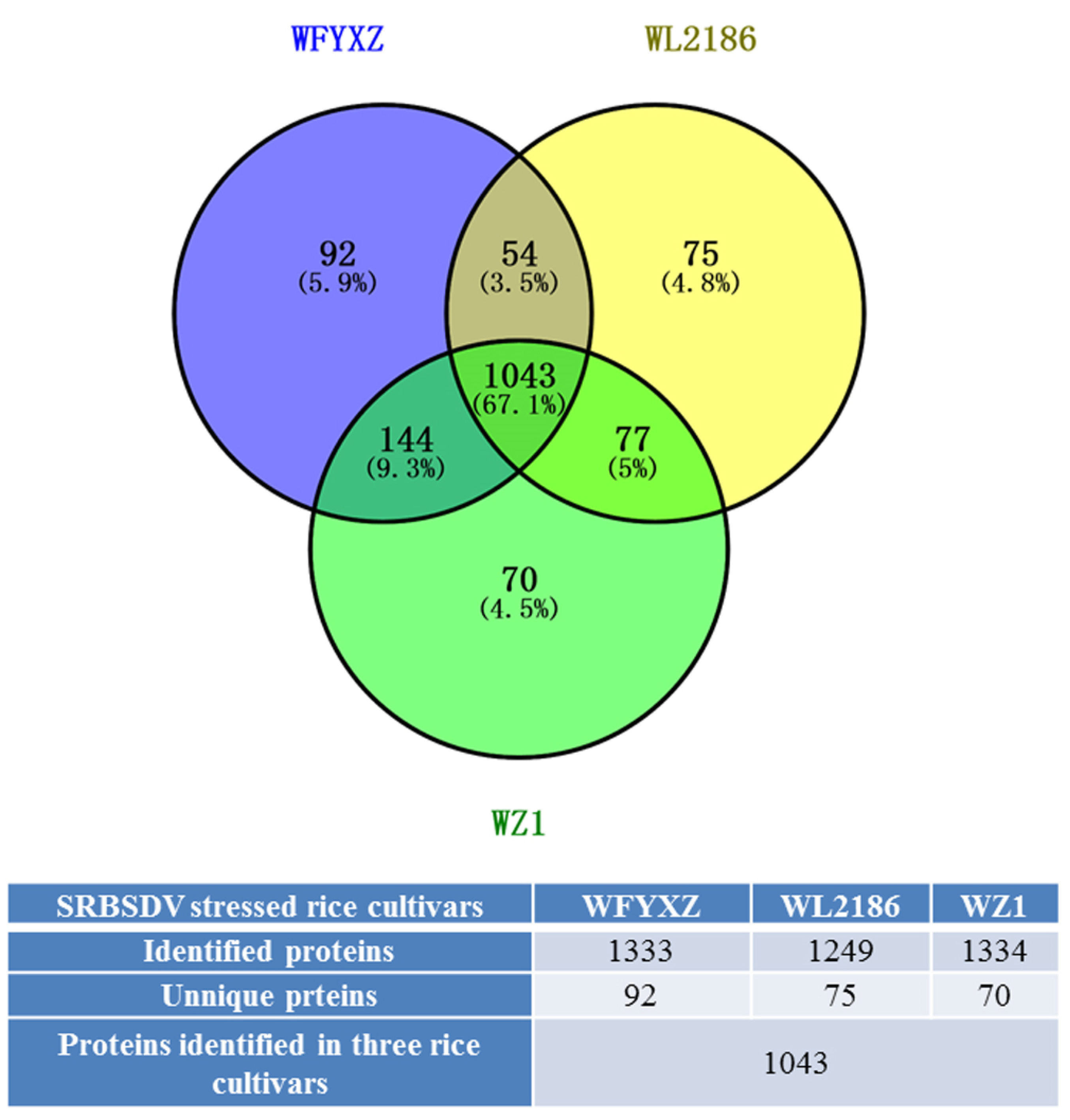
3.4.2. Overview of the Proteomic Analysis in Three SRBSDV-Stressed Cultivars
3.4.3. Systemic Resistance-Related Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yan, J.K.; Shiping, W. Toward an understanding of the molecular basis of quantitative disease resistance in rice. J. Biotechnol. 2012, 159, 283–290. [Google Scholar]
- Wang, Q.; Yang, J.; Zhou, G.H.; Zhang, H.M.; Chen, J.P.; Adams, M.J. The complete genome sequence of two isolates of southern rice black-streaked dwarf virus, a new member of the genus Fijivirus. J. Phytopathol. 2010, 158, 733–737. [Google Scholar] [CrossRef]
- Jiao, D.S.; Guo, N.M.; Chen, H.Y.; Akita, F.; Xie, L.H.; Omura, T.; Wei, T.Y. Assembly of the viroplasm by viral non-structural proteins Pns 10 is essential for persistent infection of rice ragged stunt virus in its insect vector. J. Gen. Virol. 2012, 93, 2299–2309. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.H.; Wen, J.J.; Cai, D.J. Southern rice black-streaked dwarf virus: A new proposed Fijivirus species in the family Reoviridae. Chin. Sci. Bull. 2008, 53, 3677–3685. [Google Scholar] [CrossRef]
- Zhang, H.M.; Yang, J.; Chen, J.P. A black-streaked dwarf disease on rice in China is caused by a novel Fijivirus. Arch. Virol. 2008, 153, 1893–1898. [Google Scholar] [CrossRef] [PubMed]
- Cuong, H.V.; Nguyen, V.H.; Vu, T.M.; Matsumoto, M. Rice dwarf disease in North Vietnam in 2009 is caused by southern rice black-streaked dwarf virus (SRBSDV). Bull. Inst. Trop. Agric. Kyushu Univ. 2009, 32, 85–92. [Google Scholar]
- Jiang, Y.Y.; Guo, R.; Liu, Y.; Feng, X. The occurrence and prevention profile of rice virus disease in Vietnam. Chin. Plant Prot. 2010, 30, 54–57. [Google Scholar]
- Wang, Z.C.; Yu, D.D.; Li, X.Y.; Zeng, M.J.; Chen, Z.; Bi, L.; Liu, J.J.; Jin, L.H.; Hu, D.Y.; Yang, S.; et al. The development and application of a Dot-ELISA assay for diagnosis of southern rice black-streaked dwarf disease in the field. Viruses 2012, 4, 167–183. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.C.; Li, X.Y.; Wang, W.L.; Zhang, W.Y.; Yu, L.; Hu, D.Y.; Song, B.A. Interaction research on the antiviral molecule Dufulin targeting on southern rice black streaked dwarf virus P9–1 nonstructural protein. Viruses 2015, 7, 1454–1473. [Google Scholar] [CrossRef]
- Bentur, J.S.; Viraktamath, B.C. Rice planthoppers strike back. Curr. Sci. 2008, 95, 441–443. [Google Scholar]
- Chen, Z.; Guo, R.; Zhong, L.; Qiu, G.H.; Chen, M.H.; Song, B.A.; Liu, J.J.; Fan, H.T.; Li, X.Y.; Yang, S. Cause of outbreak of southern rice black-streaked dwarf virus disease at Matian Township of Luxi County, Jiangxi Province. Guizhou Agric. Sci. 2010, 38, 118–120. [Google Scholar]
- Yan, J.; Kudo, H.; Uyeda, I.; Lee, S.Y.; Shikata, E. Conserved terminal sequences of rice ragged stunt virus genomic RNA. J. Gen. Virol. 1992, 73, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.G.; Du, Z.G.; Wang, C.Z.; Cai, L.J.; Hu, M.Q.; Lin, Q.Y.; Wu, Z.J.; Li, Y.; Xie, L.H. Identification of Pns6, a putative movement protein of RRSV, as a silencing suppressor. Virol. J. 2010, 7, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Ling, K.C.; Tiongco, E.R.; Aguiero, V.M. Rice ragged stunt, a new virus disease. Plant Dis. Rep. 1978, 62, 701–705. [Google Scholar]
- Ling, K.C.; Tiongco, E.R.; Aguiero, V.M. Host range of rice ragged stunt virus. Int. Rice Res. Newslett. 1978, 3, 8. [Google Scholar]
- Ling, K.C.; Tiongco, E.R.; Aguiero, V.M. Transmission of rice ragged stunt disease. Int. Rice Res. Newslett. 1977, 2, 11–12. [Google Scholar]
- Thanh, D.N.; Séverine, L.; Martine, B.; Hoang, A.T.; Do, N.V.; Pascal, G.; Christophe, B. P2 of Rice grassy stunt virus (RGSV) and p6 and p9 of Rice ragged stunt virus (RRSV) isolates from Vietnam exert suppressor activity on the RNA silencing pathway. Virus Genes. 2015, 51, 267–275. [Google Scholar]
- Hoang, A.T.; Zhang, H.M.; Chen, J.P.; Hébrard, E.; Zhou, G.H.; Vinh, V.N.; Cheng, J.A. Identification, characterization, and distribution of Southern rice black-streaked dwarf virus in Vietnam. Plant Dis. 2011, 95, 1063–1069. [Google Scholar] [CrossRef]
- Xu, H.X.; He, X.C.; Zheng, X.S.; Yang, Y.J.; Tian, J.C.; Lu, Z.X. Southern rice black-streaked dwarf virus (SRBSDV) directly affects the feeding and reproduction behavior of its vector, Sogatella furcifera (Horváth) (Hemiptera: Delphacidae). J. Virol. 2014, 11, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.J.; Bao, Y.Y.; Lao, S.H.; Huang, X.H.; Ye, Y.Z.; Wu, J.X.; Xu, H.J.; Zhou, X.P.; Zhang, C.X. Rice ragged stunt virus-induced apoptosis affects virus transmission from its insect vector, the brown planthopper to the rice plant. Sci. Rep. 2015. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.B.; Li, S.; Gao, R.Z.; Sun, F.; Liu, W.C.; Zhou, G.H.; Wu, J.X.; Zhou, X.P.; Zhou, Y.J. Distribution and genetic diversity of southern rice black-streaked dwarf virus in China. J. Virol. 2013, 10, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Velusamy, R.; Heinrichs, E.A. Electronic monitoring of feeding behavior of Nilaparvatalugens (Homoptera: Delphacidae) on resistant and susceptible rice cultivars. Environ. Entomol. 1986, 15, 678–682. [Google Scholar] [CrossRef]
- Khush, G.S.; Brar, D.S. Genetics of resistance to insects in crop plants. Adv. Agron. 1991, 45, 223–274. [Google Scholar]
- Keiichiro, M.; Tomomi, T.; Kazuhiro, Y.; Junichi, S.; Misturu, O.; Masatoshi, O.; Masaya, M. Quantitative analysis of southern rice black-streaked dwarf virus in Sogatella furcifera and virus threshold for transmission. Phytopathology 2015, 105, 550–554. [Google Scholar]
- Chen, Z.; Song, B.A. The Prevention and Control Technology of Southern Rice Black-Streaked Dwarf Virus Disease, 1st ed.; Chemical Industrial Press: Beijing, China, 2011; pp. 25–48. [Google Scholar]
- Wang, F.; Qin, G.Z.; Sui, Z.H.; Wang, Z.H.; Wang, Z.Y.; Yu, J.L.; Zhang, J.R. Improved method for assaying maize plant resistance to maize rough dwarf disease by artificial inoculation and real-time RT-PCR. Eur. J. Plant Pathol. 2006, 116, 289–300. [Google Scholar] [CrossRef]
- He, P.; Liu, J.J.; He, M.; Wang, Z.C.; Chen, Z.; Guo, R.; Correll, J.C.; Yang, S.; Song, B.A. Quantitative detection of relative expression levels of the whole genome of Southern rice black-streaked dwarf virus and its replication in different hosts. Virol. J. 2013, 10, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.J.; Chen, J.; Liu, T.W.; Simon, M.; Wang, W.H.; Chen, J.; Wu, F.H.; Liu, X.; Shen, Z.J.; Zheng, H.L. Comparative proteomic analysis of differential responses of Pinus massoniana and Taxus wallichiana var. mairei to simulated acid rain. Int. J. Mol. Sci. 2014, 15, 4333–4355. [Google Scholar] [PubMed]
- Fang, W.P.; Xie, D.Y.; Zhu, H.Q.; Li, W.; Xu, Z.Z.; Yang, L.R.; Li, Z.F.; Sun, L.; Wang, J.X.; Nie, L.H.; et al. Comparative proteomic analysis of Gossypium thurberi in response to Verticillium dahliae inoculation. Int. J. Mol. Sci. 2015, 16, 25121–25140. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.P.; Zhang, Q.Y.; Tang, Z.C.; Su, W.A.; Sun, W.N. Comparative proteomic analysis provides new insights into chilling stress responses in rice. Mol. Cell. Proteom. 2006, 5, 484–496. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Hong, H.; Ye, L.C.; Wang, Y.L.; Chen, H.W.; Liu, J.F. Label-free quantitative proteomic analysis of right ventricular remodeling in infant Tetralogy of Fallot patients. J. Proteom. 2013, 84, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Guo, Q.; Chen, B.H.; Li, X.Y.; Wang, Z.C.; He, P.; Yan, F.; Hu, D.Y.; Yang, S. Development of proteomic technology of shotgun and label free combined with multiple reaction monitoring to simultaneously detect southern rice black-streaked dwarf virus and rice ragged stunt virus. Virus Dis. 2013, 25, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Neuhauser, N.; Michalski, A.; Scheltema, R.A.; Olsen, J.V.; Mann, M. Andromeda: A peptide search engine integrated into the MaxQuant environment. J. Proteome Res. 2011, 10, 1794–1805. [Google Scholar] [CrossRef] [PubMed]
- Luber, C.A.; Cox, J.; Lauterbach, H.; Fancke, B.; Selbach, M.; Tschopp, J.; Akira, S.; Wiegand, M.; Hochrein, H.; O’Keeffe, M.; et al. Quantitative proteomics reveals subset-specific viral recognition in dendritic cells. Immunity 2010, 32, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Sta, C.F.C.; Hull, R.; Azzam, O. Changes in level of virus accumulation and incidence of infection are critical in the characterization of Rice tungro bacilliform virus (RTBV) resistance in rice. Arch. Virol. 2003, 148, 1465–1483. [Google Scholar]
- Fang, X.P.; Chen, J.P.; Dai, L.Y.; Ma, H.S.; Zhang, H.M.; Yang, J.; Wang, F.; Yan, C.Q. Proteomic dissection of plant responses to various pathogens. Proteomics 2015, 15, 1525–1543. [Google Scholar] [CrossRef] [PubMed]
- Ryals, J.; Uknes, S.; Ward, E. Systemic acquired resistance. Plant Physiol. 1994, 104, 1109–1112. [Google Scholar] [CrossRef] [PubMed]
- Luna, E.; Bruce, T.J.A.; Roberts, M.R.; Flors, V.; Ton, J. Next-Generation Systemic Acquired Resistance. Plant Physiol. 2012, 158, 844–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durrant, W.E.; Dong, X. Systemic Acquired Resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, A.M.; Doerner, P.; Dixon, R.A.; Lamb, C.J.; Cameron, R.K. A putative lipid transfer protein involved in systemic resistance signalling in Arabidopsis. Nature 2002, 419, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Sun, T.J.; Zhang, Y.L. ER quality control components UGGT and STT3a are required for activation of defense responses in Bir1–1. PLoS ONE 2015, 10, e0120245. [Google Scholar] [CrossRef] [PubMed]
- Noutoshi, Y.; Kuromori, T.; Wada, T.; Hirayama, T.; Kamiya, A.; Imura, Y.; Yasuda, M.; Nakashita, H.; Shirasu, K.; Shinozaki, K. Loss of necrotic spotted lesions 1 associates with cell death and defense responses in Arabidopsis thaliana. Plant Mol. Biol. 2006, 62, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Morita-Yamamuro, C.; Tsutsui, T.; Sato, M.; Yoshioka, H.; Tamaoki, M.; Ogawa, D.; Matsuura, H.; Yoshihara, T.; Ikeda, A.; Uyeda, I.; et al. The Arabidopsis gene CAD1 controls programmed cell death in the plant immune system and encodes a protein containing a MACPF domain. Plant Cell Physiol. 2005, 46, 902–912. [Google Scholar] [CrossRef] [PubMed]
- Elvira, M.I.; Galdeano, M.M.; Gilardi, P.; Garcia, L.; Serra, M.T. Proteomic analysis of pathogenesisrelated proteins (PRs) induced by compatible and incompatible interactions of pepper mild mottle virus (PMMoV) in Capsicum chinense L3 plants. J. Exp. Bot. 2008, 59, 1253–1265. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.J.; Lu, S.; Xu, Y.H.; Wang, J.W.; Chen, X.Y. A cotton cDNA (GaPR-10) encoding a pathogenesis-related 10 protein with in vivo ribonuclease activity. Plant Sci. 2002, 162, 629–636. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Number | Abbreviation | Name of Variety | Infected with SRBSDV (+/−) | Infected with RRSV (+/−) |
|---|---|---|---|---|
| Group 1 | H658 | Huxiang658 | + | + |
| Z1 | Zhongzheyou1 | + | − | |
| Group 2 | N18 | Neixiangyou18 | + | + |
| L2161 | Liangyou2161 | + | − | |
| K182 | Kefeng182 | + | − | |
| N5Y39 | Nei5You39 | + | − | |
| L2186 | Liangyou2186 | + | + | |
| Y696 | YLiangyou696 | + | + | |
| C2Y498 | Chuannong2 You 498 | + | + | |
| Group 3 | Y17 | Yunguang17 | + | + |
| N1 | Neixiangyou1 | − | + | |
| L816 | Liangyou816 | + | + | |
| IIY629 | IIYou629 | + | + | |
| IIY58 | IIYou58 | + | − | |
| L15 | Liangyou15 | + | + | |
| IIY6 | IIYou6 | + | + | |
| D8 | Deyou8 | + | + | |
| Group 4 | YX1 | Yingxuan1 | + | − |
| S5 | Shenzhou5 | + | + | |
| C3727 | Chuanyou3727 | + | + | |
| M19 | Mengyou19 | + | + | |
| Y1 | Yingxiang1 | + | + | |
| Control | FYXZ | Fengyouxiangzhan | + | − |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Yu, L.; Jin, L.; Wang, W.; Zhao, Q.; Ran, L.; Li, X.; Chen, Z.; Guo, R.; Wei, Y.; et al. Evaluation of Rice Resistance to Southern Rice Black-Streaked Dwarf Virus and Rice Ragged Stunt Virus through Combined Field Tests, Quantitative Real-Time PCR, and Proteome Analysis. Viruses 2017, 9, 37. https://doi.org/10.3390/v9020037
Wang Z, Yu L, Jin L, Wang W, Zhao Q, Ran L, Li X, Chen Z, Guo R, Wei Y, et al. Evaluation of Rice Resistance to Southern Rice Black-Streaked Dwarf Virus and Rice Ragged Stunt Virus through Combined Field Tests, Quantitative Real-Time PCR, and Proteome Analysis. Viruses. 2017; 9(2):37. https://doi.org/10.3390/v9020037
Chicago/Turabian StyleWang, Zhenchao, Lu Yu, Linhong Jin, Wenli Wang, Qi Zhao, Longlu Ran, Xiangyang Li, Zhuo Chen, Rong Guo, Yongtian Wei, and et al. 2017. "Evaluation of Rice Resistance to Southern Rice Black-Streaked Dwarf Virus and Rice Ragged Stunt Virus through Combined Field Tests, Quantitative Real-Time PCR, and Proteome Analysis" Viruses 9, no. 2: 37. https://doi.org/10.3390/v9020037
APA StyleWang, Z., Yu, L., Jin, L., Wang, W., Zhao, Q., Ran, L., Li, X., Chen, Z., Guo, R., Wei, Y., Yang, Z., Liu, E., Hu, D., & Song, B. (2017). Evaluation of Rice Resistance to Southern Rice Black-Streaked Dwarf Virus and Rice Ragged Stunt Virus through Combined Field Tests, Quantitative Real-Time PCR, and Proteome Analysis. Viruses, 9(2), 37. https://doi.org/10.3390/v9020037