
Schrödinger’s Cheshire Cat: Are Haploid Emiliania huxleyi Cells Resistant to Viral Infection or Not?

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Nucleic Acid Extraction
2.3. cDNA Synthesis
2.4. PCR and RT-PCR Amplification
2.5. Sequencing and Alignment
2.6. Tiling Array
3. Results
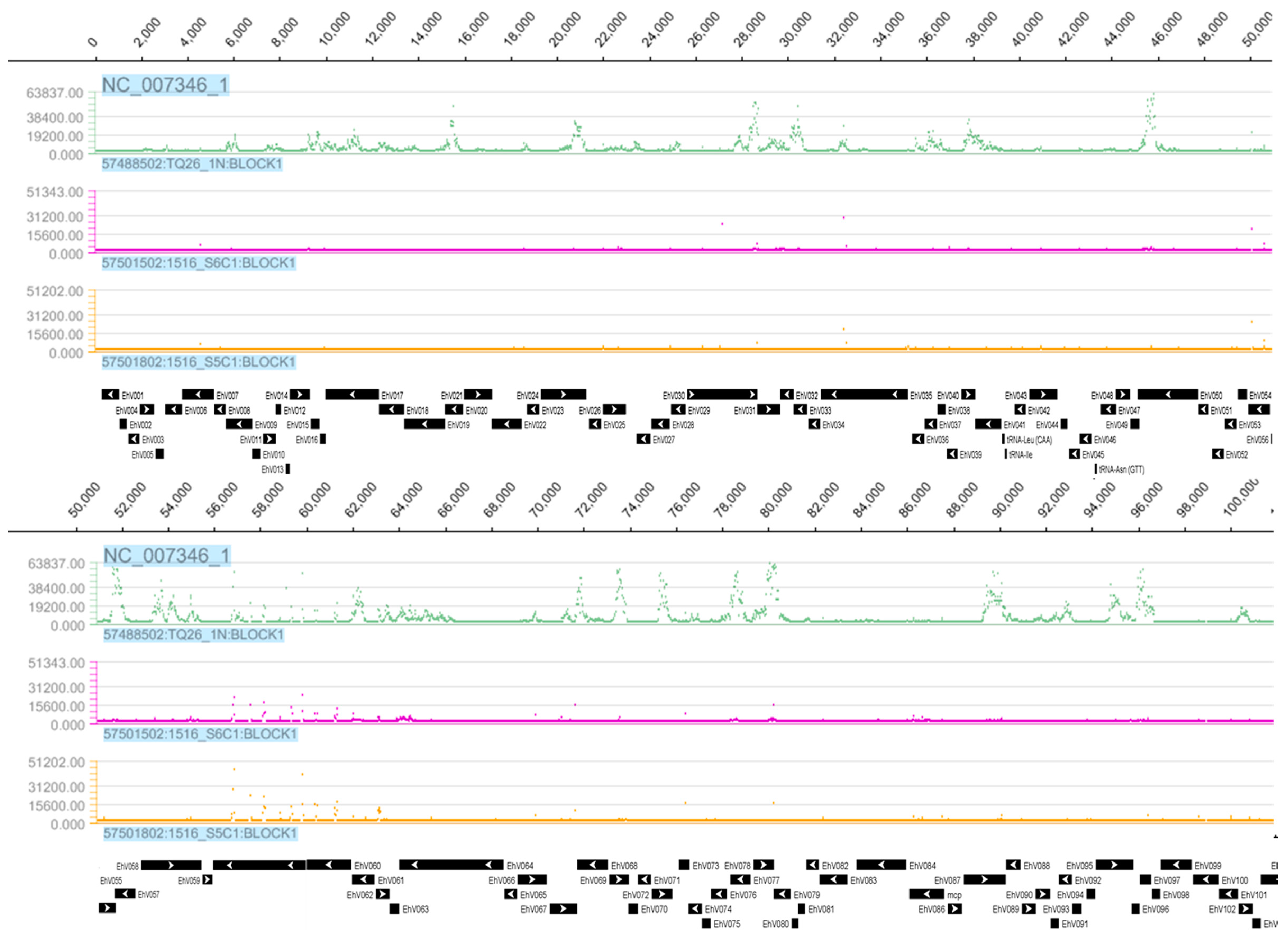
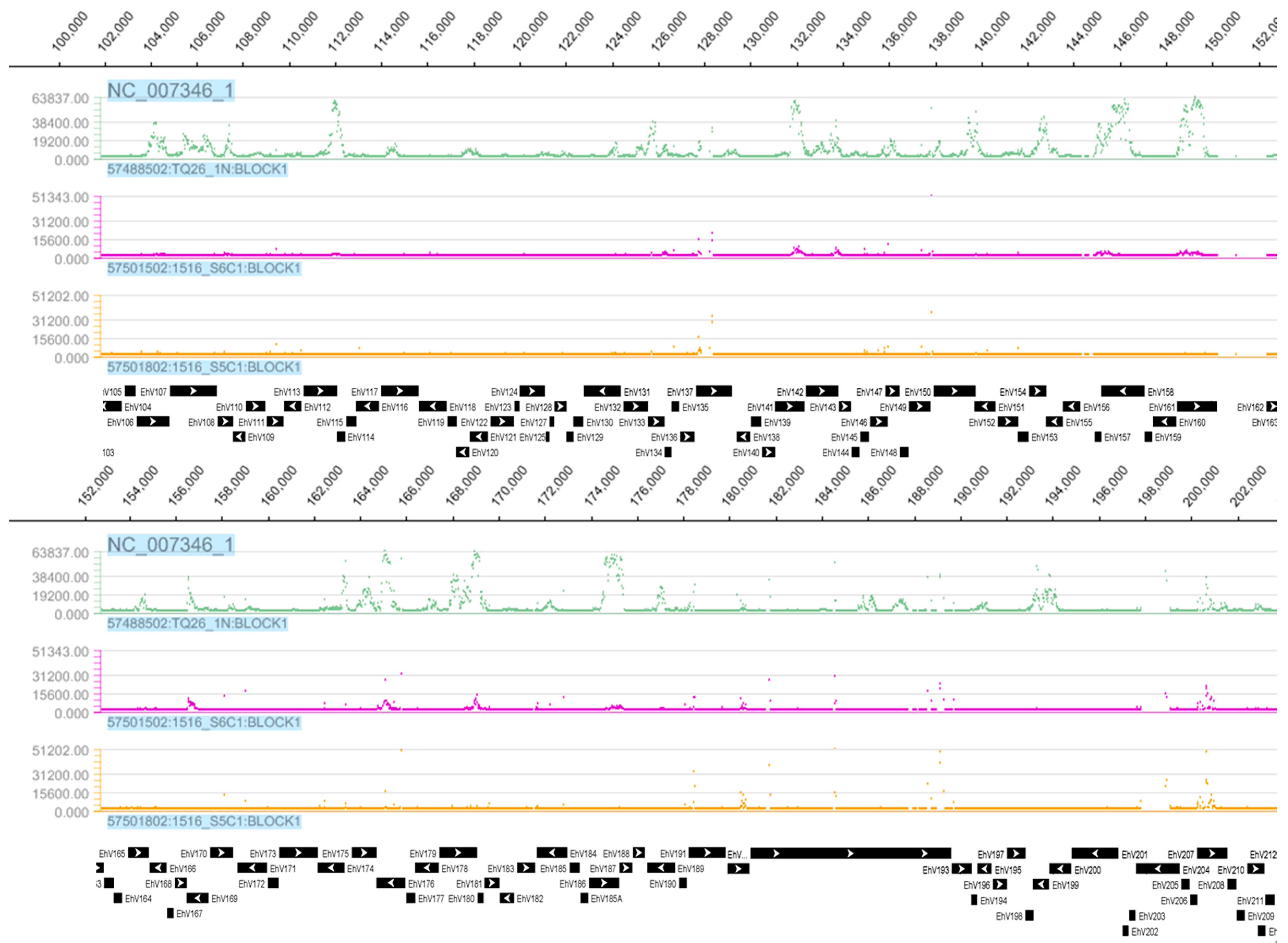
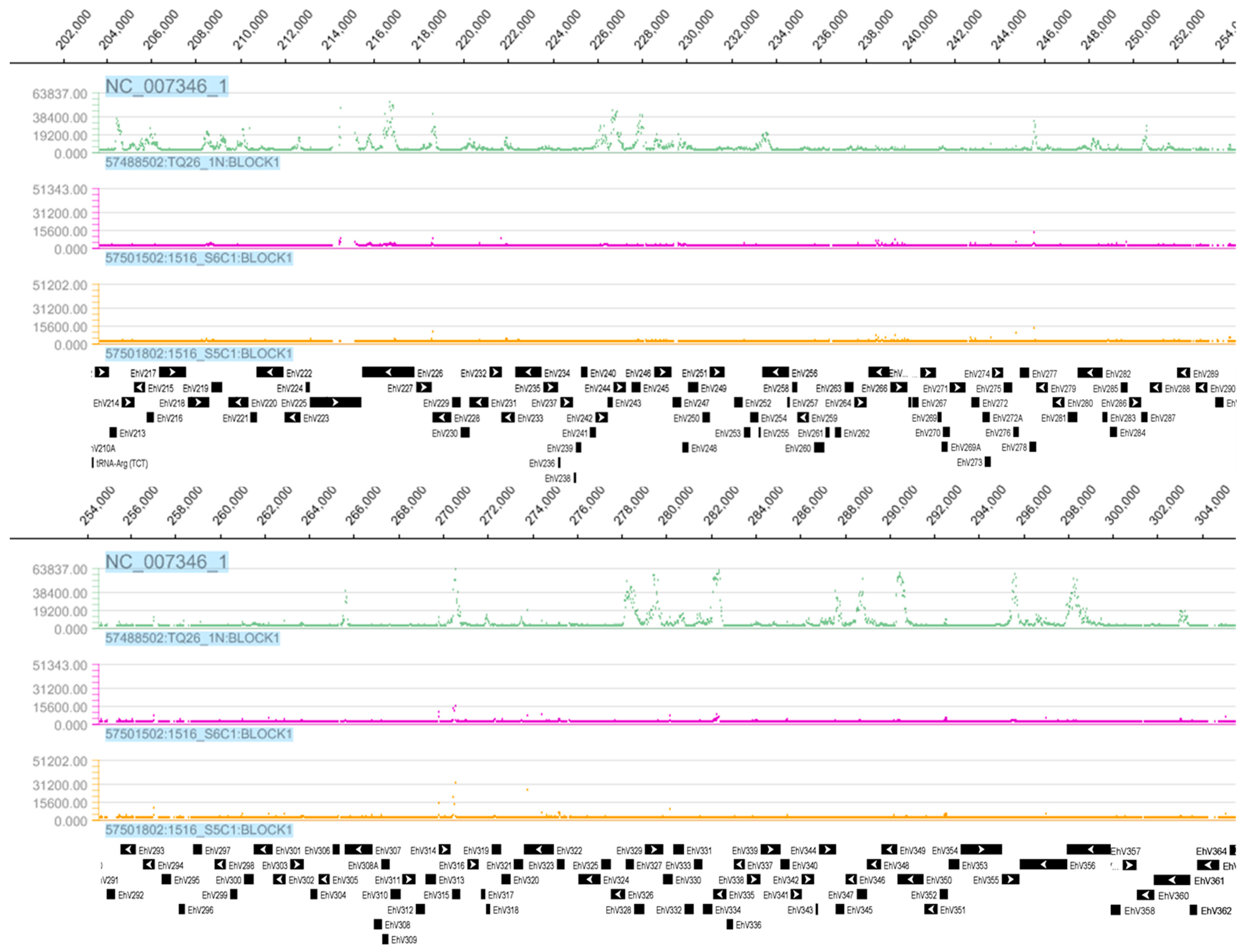
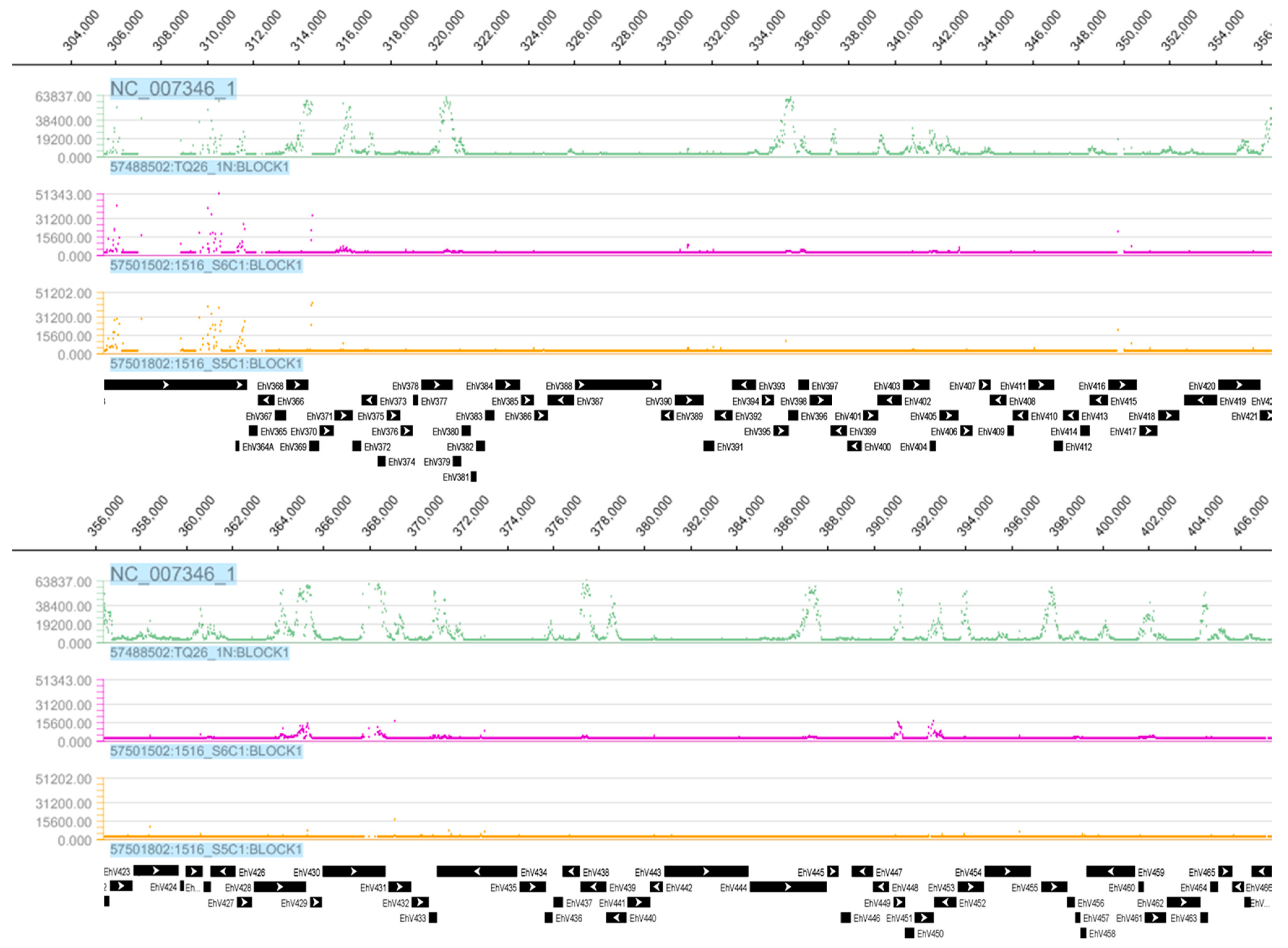
3.1. Tiling Array
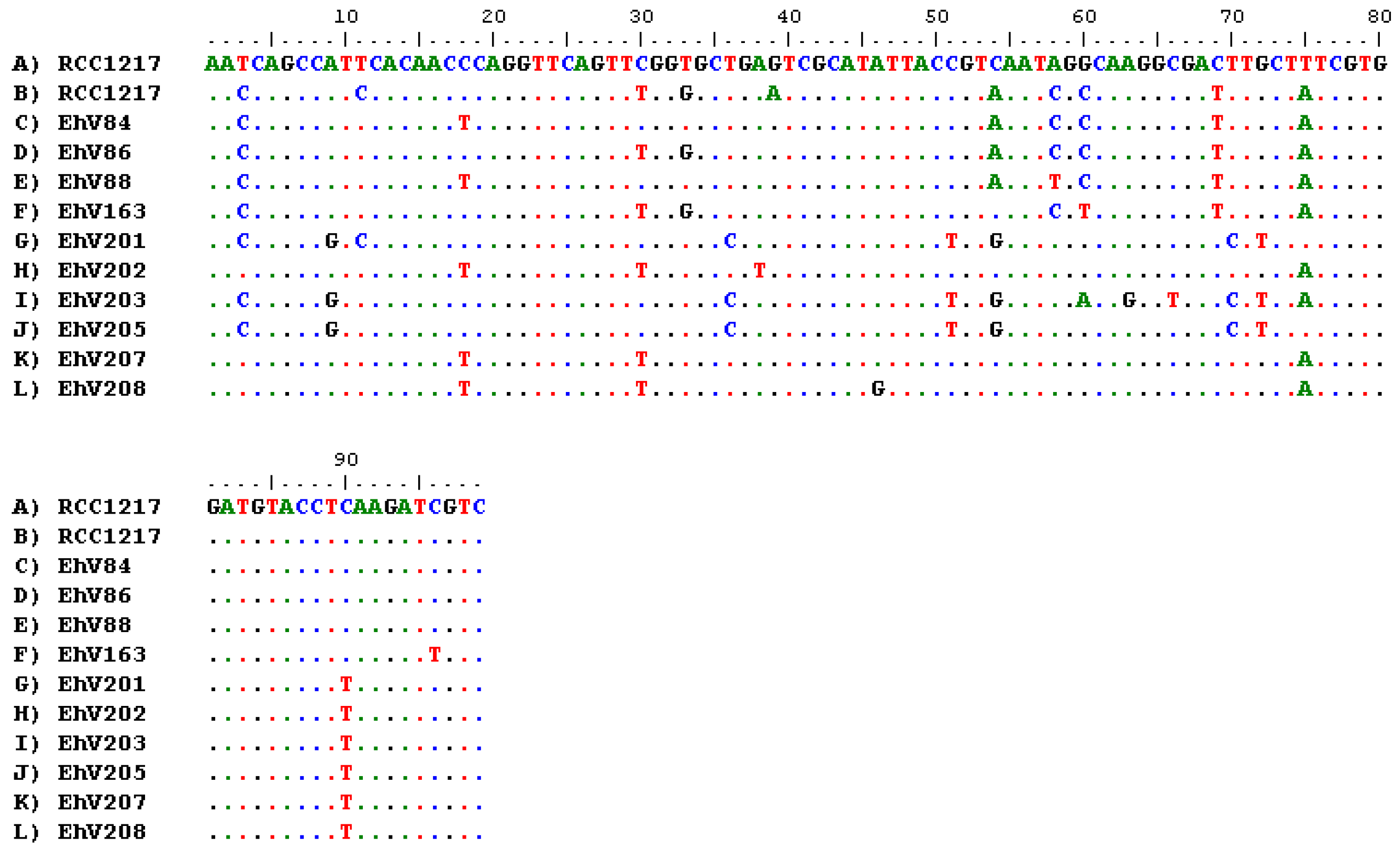
3.2. E. huxleyi Strain Confirmation
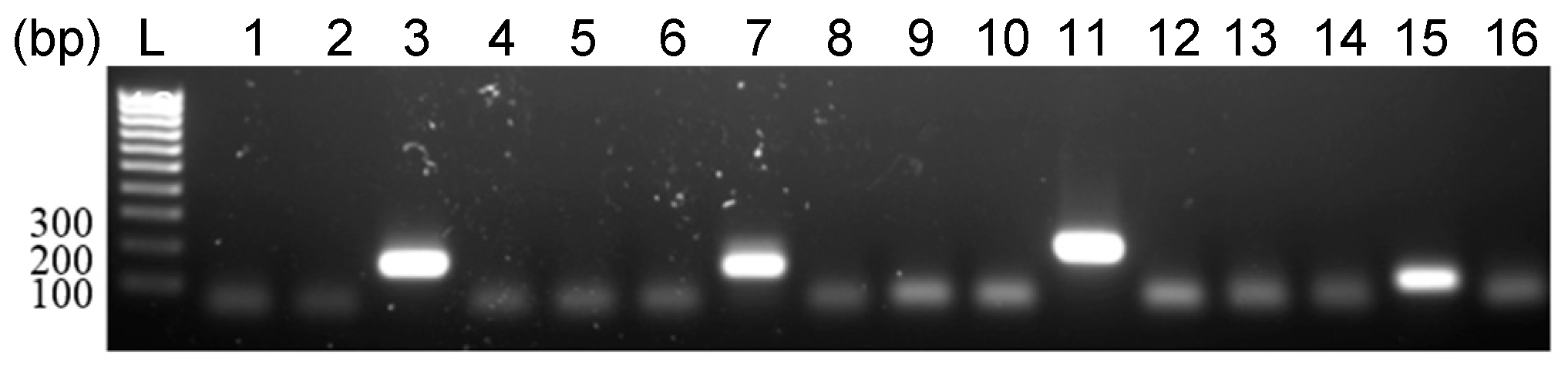
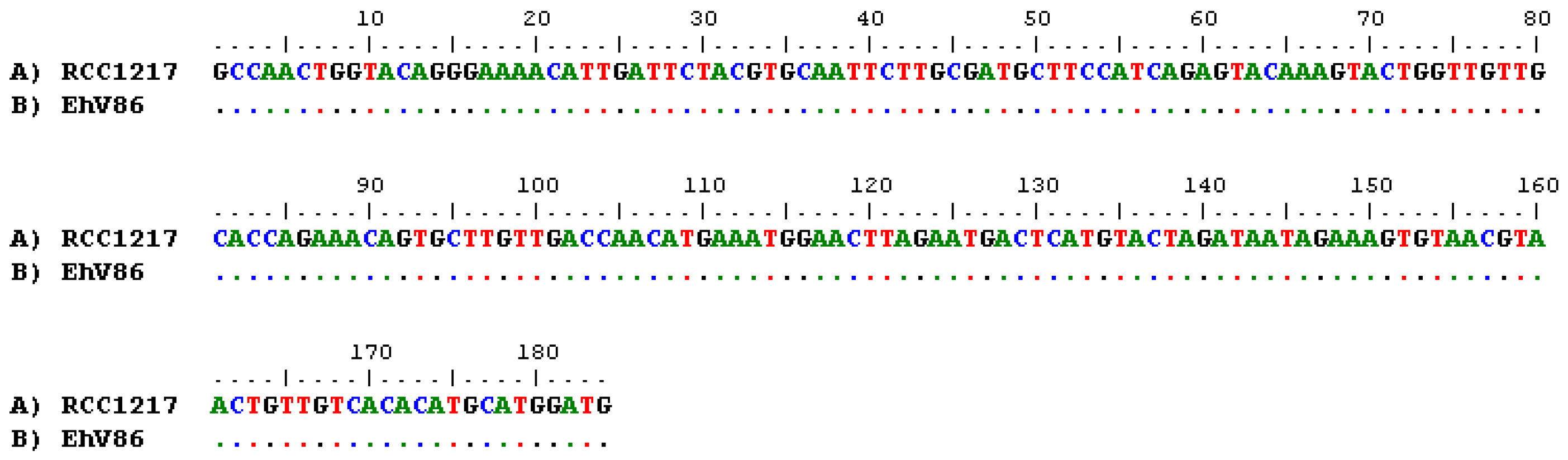
3.3. Detection of EhV DNA
3.4. Detection of EhV RNA
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Paasche, E. A review of the coccolithophorid Emiliania huxleyi (prymnesiophyceae), with particular reference to growth, coccolith formation, and calcification-photosynthesis interactions. Phycologia 2001, 40, 503–529. [Google Scholar] [CrossRef]
- Westbroek, P.; Brown, C.W.; Vanbleijswijk, J.; Brownlee, C.; Brummer, G.J.; Conte, M.; Egge, J.; Fernandez, E.; Jordan, R.; Knappertsbusch, M. A model system approach to biological climate forcing—The example of Emiliania huxleyi. Glob. Planet. Chang. 1993, 8, 27–46. [Google Scholar] [CrossRef]
- Holligan, P.M.; Viollier, M.; Harbour, D.S.; Camus, P.; Champagnephilippe, M. Satellite and ship studies of coccolithophore production along a continental-shelf edge. Nature 1983, 304, 339–342. [Google Scholar] [CrossRef]
- Elderfield, H. Climate change: Carbonate mysteries. Science 2002, 296, 1618–1621. [Google Scholar] [CrossRef] [PubMed]
- Riebesell, U.; Schulz, K.G.; Bellerby, R.G.J.; Botros, M.; Fritsche, P.; Meyerhofer, M.; Neill, C.; Nondal, G.; Oschlies, A.; Wohlers, J.; et al. Enhanced biological carbon consumption in a high CO2 ocean. Nature 2007, 450, 545–548. [Google Scholar] [CrossRef] [PubMed]
- Green, J.C.; Course, P.A.; Tarran, G.A. The life-cycle of Emiliania huxleyi: A brief review and a study of relative ploidy levels analysed by flow cytometry. J. Mar. Syst. 1996, 9, 33–44. [Google Scholar] [CrossRef]
- Klaveness, D. Coccolithus huxleyi (lohm.) kamptn ii. The flagellate cell, aberrant cell types, vegetative propagation and life cycles. Eur. J. Phycol. 1972, 7, 309–318. [Google Scholar] [CrossRef]
- Schroeder, D.C.; Oke, J.; Hall, M.; Malin, G.; Wilson, W. Virus succession observed during an Emiliania huxleyi bloom. Appl. Environ. Microbiol. 2003, 69, 2484–2490. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, D.C.; Oke, J.; Malin, G.; Wilson, W.H. Coccolithovirus (phycodnaviridae): Characterisation of a new large dsdna algal virus that infects Emiliania huxleyi. Arch. Virol. 2002, 147, 1685–1698. [Google Scholar] [CrossRef] [PubMed]
- Wilson, W.H.; Schroeder, D.C.; Allen, M.J.; Holden, M.T.; Parkhill, J.; Barrell, B.G.; Churcher, C.; Hamlin, N.; Mungall, K.; Norbertczak, H. Complete genome sequence and lytic phase transcription profile of a coccolithovirus. Science 2005, 309. [Google Scholar] [CrossRef] [PubMed]
- Frada, M.; Probert, I.; Allen, M.J.; Wilson, W.H.; de Vargas, C. The “cheshire cat” escape strategy of the coccolithophore Emiliania huxleyi in response to viral infection. Proc. Natl. Acad. Sci. USA 2008, 105, 15944–15949. [Google Scholar] [CrossRef] [PubMed]
- VanValen, L. A new evolutionary law. Evolut. Theory 1973, 1. [Google Scholar]
- Rosenwasser, S.; Mausz, M.A.; Schatz, D.; Sheyn, U.; Malitsky, S.; Aharoni, A.; Weinstock, E.; Tzfadia, O.; Ben-Dor, S.; Feldmesser, E.; et al. Rewiring host lipid metabolism by large viruses determines the fate of Emiliania huxleyi, a bloom-forming alga in the ocean. Plant Cell 2014, 26, 2689–2707. [Google Scholar] [CrossRef] [PubMed]
- Read, B.A.; Kegel, J.; Klute, M.J.; Kuo, A.; Lefebvre, S.C.; Maumus, F.; Mayer, C.; Miller, J.; Monier, A.; Salamov, A.; et al. Pan genome of the phytoplankton Emiliania underpins its global distribution. Nature 2013, 499, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Monier, A.; Pagarete, A.; de Vargas, C.; Allen, M.J.; Read, B.; Claverie, J.-M.; Ogata, H. Horizontal gene transfer of an entire metabolic pathway between a eukaryotic alga and its DNA virus. Genome Res. 2009, 19, 1441–1449. [Google Scholar] [CrossRef] [PubMed]
- Bidle, K.D. The molecular ecophysiology of programmed cell death in marine phytoplankton. Annu. Rev. Mar. Sci. 2015, 7, 1–596. [Google Scholar] [CrossRef] [PubMed]
- Vardi, A.; Van Mooy, B.A.S.; Fredricks, H.F.; Popendorf, K.J.; Ossolinski, J.E.; Haramaty, L.; Bidle, K.D. Viral glycosphingolipids induce lytic infection and cell death in marine phytoplankton. Science 2009, 326, 861–865. [Google Scholar] [CrossRef] [PubMed]
- Hannun, Y.A.; Obeid, L.M. Principles of bioactive lipid signalling: Lessons from sphingolipids. Nat. Rev. Mol. Cell Biol. 2008, 9, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Bidle, K.D.; Haramaty, L.; Barcelos e Ramos, J.; Falkowski, P. Viral activation and recruitment of metacaspases in the unicellular coccolithophore, Emiliania huxleyi. Proc. Natl. Acad. Sci. USA 2007, 104, 6049–6054. [Google Scholar] [CrossRef] [PubMed]
- Brussaard, C.P.D.; Kempers, R.S.; Kop, A.J.; Riegman, R.; Heldal, M. Virus-like particles in a summer bloom of Emiliania huxleyi in the north sea. Aquat. Microb. Ecol. 1996, 10, 105–113. [Google Scholar] [CrossRef]
- Hunter, J.E.; Frada, M.J.; Fredricks, H.F.; Vardi, A.; Van Mooy, B.A.S. Targeted and untargeted lipidomics of Emiliania huxleyi viral infection and life cycle phases highlights molecular biomarkers of infection, susceptibility and ploidy. Front. Mar. Sci. 2015, 2, 81. [Google Scholar] [CrossRef]
- Jacquet, S.; Heldal, M.; Iglesias-Rodriguez, D.; Larsen, A.; Wilson, W.H.; Bratbak, G. Flow cytometric analysis of an Emiliana huxleyi bloom terminated by viral infection. Aquat. Microb. Ecol. 2002, 27, 111–124. [Google Scholar] [CrossRef]
- Schroeder, D.C.; Biggi, G.F.; Hall, M.; Davy, J.; Martinez-Martinez, J.; Richardson, A.J.; Malin, G.; Wilson, W.H. A genetic marker to separate Emiliania huxleyi (Prymnesiophyceae) morphotypes. J. Phycol. 2005, 41, 874–879. [Google Scholar] [CrossRef]
- Krueger-Hadfield, S.A.; Balestreri, C.; Schroeder, J.; Highfield, A.; Helaouët, P.; Allum, J.; Moate, R.; Lohbeck, K.T.; Miller, P.I.; Riebesell, U.; et al. Genotyping an Emiliania huxleyi (Prymnesiophyceae) bloom event in the north sea reveals evidence of asexual reproduction. Biogeosciences 2014, 11, 5215–5234. [Google Scholar] [CrossRef]
- Bioedit. Available online: http://www.mbio.ncsu.edu/bioedit/bioedit.html (accessed 14 March 2017).
- Gledhill, M.; Devez, A.; Highfield, A.; Singleton, C.; Achterberg, E.P.; Schroeder, D. Effect of metals on the lytic cycle of the coccolithovirus, EhV86. Front. Microbiol. 2012, 3, 155. [Google Scholar] [CrossRef] [PubMed]
- von Dassow, P.; Ogata, H.; Probert, I.; Wincker, P.; Da Silva, C.; Audic, S.; Claverie, J.-M.; de Vargas, C. Transcriptome analysis of functional differentiation between haploid and diploid cells of Emiliania huxleyi, a globally significant photosynthetic calcifying cell. Genome Biol. 2009, 10, R114. [Google Scholar] [CrossRef] [PubMed]
- Kegel, J.; Blaxter, M.; Allen, M.J.; Metfies, K.; Wilson, W.; Valentin, K. Transcriptional host-virus interaction of Emiliania huxleyi (Haptophyceae) and EhV-86 deduced from combined analysis of expressed sequence tags and microarrays. Eur. J. Phycol. 2010, 45, 1–12. [Google Scholar] [CrossRef]
- Allen, M.J.; Forster, T.; Schroeder, D.C.; Hall, M.; Roy, D.; Ghazal, P.; Wilson, W.H. Locus-specific gene expression pattern suggests a unique propagation strategy for a giant algal virus. J. Virol. 2006, 80, 7699–7705. [Google Scholar] [CrossRef] [PubMed]
- Pagarete, A.; Allen, M.J.; Wilson, W.H.; Kimmance, S.A.; De Vargas, C. Host–virus shift of the sphingolipid pathway along an Emiliania huxleyi bloom: Survival of the fattest. Environ. Microbiol. 2009, 11, 2840–2848. [Google Scholar] [CrossRef] [PubMed]
- The National Center for Biotechnology Information . Available online: https://www.ncbi.nlm.nih.gov/ (accessed 14 March 2017).
- Hsia, K.-C.; Chak, K.-F.; Liang, P.-H.; Cheng, Y.-S.; Ku, W.-Y.; Yuan, H.S. DNA binding and degradation by the hnh protein cole7. Structure 2004, 12, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, D.C.; Park, Y.; Yoon, H.-M.; Lee, Y.S.; Kang, S.W.; Meints, R.H.; Ivey, R.G.; Choi, T.-J. Genomic analysis of the smallest giant virus—Feldmannia sp. Virus 158. Virology 2009, 384, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Senkevich, T.G.; White, C.L.; Koonin, E.V.; Moss, B. A viral member of the ERV1/ALR protein family participates in a cytoplasmic pathway of disulfide bond formation. Proc. Natl. Acad. Sci. USA 2000, 97, 12068–12073. [Google Scholar] [CrossRef] [PubMed]
- Rosenwasser, S.; Graff van Creveld, S.; Schatz, D.; Malitsky, S.; Tzfadia, O.; Aharoni, A.; Levin, Y.; Gabashvili, A.; Feldmesser, E.; Vardi, A. Mapping the diatom redox-sensitive proteome provides insight into response to nitrogen stress in the marine environment. Proc. Natl. Acad. Sci. USA 2014, 111, 2740–2745. [Google Scholar] [CrossRef] [PubMed]
- Frada, M.J.; Bidle, K.D.; Probert, I.; de Vargas, C. In situ survey of life cycle phases of the coccolithophore Emiliania huxleyi (Haptophyta). Environ. Microbiol. 2012, 14, 1558–1569. [Google Scholar] [CrossRef] [PubMed]
- Mackinder, L.C.M.; Worthy, C.A.; Biggi, G.; Hall, M.; Ryan, K.P.; Varsani, A.; Harper, G.M.; Wilson, W.H.; Brownlee, C.; Schroeder, D.C. A unicellular algal virus, Emiliania huxleyi virus 86, exploits an animal-like infection strategy. J. Gen. Virol. 2009, 90, 2306–2316. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.J.; Schroeder, D.C.; Wilson, W.H. Preliminary characterisation of repeat families in the genome of EhV-86, a giant algal virus that infects the marine microalga Emiliania huxleyi. Arch. Virol. 2006, 151, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Mackinder, L.; Wheeler, G.; Schroeder, D.; Riebesell, U.; Brownlee, C. Molecular mechanisms underlying calcification in coccolithophores. Geomicrobiol. J. 2010, 27, 585–595. [Google Scholar] [CrossRef]
- Kawakami, S.K.; Gledhill, M.; Achterberg, E.P. Production of phytochelatins and glutathione by marine phytoplankton in response to metal stress1. J. Phycol. 2006, 42, 975–989. [Google Scholar] [CrossRef]
- Schatz, D.; Shemi, A.; Rosenwasser, S.; Sabanay, H.; Wolf, S.G.; Ben-Dor, S.; Vardi, A. Hijacking of an autophagy-like process is critical for the life cycle of a DNA virus infecting oceanic algal blooms. New Phytol. 2014, 204, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Malitsky, S.; Ziv, C.; Rosenwasser, S.; Zheng, S.; Schatz, D.; Porat, Z.; Ben-Dor, S.; Aharoni, A.; Vardi, A. Viral infection of the marine alga Emiliania huxleyi triggers lipidome remodeling and induces the production of highly saturated triacylglycerol. New Phytol. 2016, 210, 88–96. [Google Scholar] [CrossRef]
- Iyer, L.M.; Tahiliani, M.; Rao, A.; Aravind, L. Prediction of novel families of enzymes involved in oxidative and other complex modifications of bases in nucleic acids. Cell Cycle 2009, 8, 1698–1710. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.N.; Burbank, D.E.; Uher, L.; Rabussay, D.; Van Etten, J.L. Restriction endonuclease activity induced by PBCV-1 virus infection of a chlorella-like green alga. Mol. Cell. Biol. 1986, 6, 1430–1439. [Google Scholar] [CrossRef] [PubMed]
- Wilson, W.H. Coccolithovirus-emiliania huxleyi dynamics: An introduction to the coccolithovirocell. Perspect. Phycol. 2015, 2, 91–103. [Google Scholar]
- ViralZone. Available online: http://viralzone.expasy.org/ (accessed 14 March 2017).
- Stevens, K.; Weynberg, K.; Bellas, C.; Brown, S.; Brownlee, C.; Brown, M.T.; Schroeder, D.C. A novel evolutionary strategy revealed in the phaeoviruses. PLoS ONE 2014, 9, e86040. [Google Scholar] [CrossRef] [PubMed]
- Suttle, C.A.; Feng, C. Mechanisms and rates of decay of marine viruses in seawater. Appl. Environ. Microbiol. 1992, 58, 3721–3729. [Google Scholar] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Origin | Date of Isolation |
|---|---|---|
| Emiliania huxleyi RCC1216 (TQ26 2N) | Tasman Sea | October 1998 |
| E. huxleyi RCC1217 (TQ26 1N) | from strain E. huxleyi RCC1216 | July 1999 |
| E. huxleyi CCMP1516 | South Pacific | July 1991 |
| E. huxleyi Virus 86 | English Channel [9] | July 1999 |
| Primer | Sequence (5′ to 3′) | Target (CDS) | Ta (°C) | Reference |
|---|---|---|---|---|
| qCBP_F | AGTCTCTCGACGCTGCCTC | GPA | 60 | [24] |
| qCBP_R | TGGCCTAGCACCAGTCTTTGG | |||
| MCP_F2 | TTCGCGCTCGAGTCGATC | MCP (ehv085) | 60 | [8] |
| MCP_R2 | GACCTTTAGGCCAGGGAG | |||
| EhVhel_F | GCCAACTGGTACAGGGAAAA | Helicase (ehv104) | 54 | [26] |
| EhVhel_R | CATCCATGCATGTGTCACAA | |||
| EhVPCNA_F | GGGCATTTCATTTGCCATAC | PCNA (ehv440) | 54 | [26] |
| EhVPCNA_R | ATTCTCCGTCGACAAACGC | |||
| EhVpol_F | TATAATGCACGCCAACTTGC | DNA pol (ehv030) | 54 | [26] |
| EhVpol_R | GCAATTGCACCAAGTGGATA | |||
| DHC1b_F | GCTTTCTCACTGCGCTCAT | Flagellar inner DHC | 55 | [27] |
| DHC1b_R | GTAGAGCGGGCACGAGTACA | |||
| DHCb_F | TGAACCTCGTCCTCAACACA | Flagellar outer DHC | 55 | [27] |
| DHCb_R | GAATCATCGGCATCACTGG |
| CHIP_ID | Image Name | Dye | Sample Description * | Virus Inoculation |
|---|---|---|---|---|
| 57501502 | 57501502_532.tif | Cy3 | 1516 S6C1 | 3.5 h p.i. |
| 57501502_635.tif | Cy5 | 1516 S5C1 | 0.5 h p.i. | |
| 57501802 | 57501802_532.tif | Cy3 | 1516 S5C1 | 0.5 h p.i. |
| 57501802_635.tif | Cy5 | 1516 S5C4 | No | |
| 58219802 | 58219802_532.tif | Cy3 | 1516 S6C4 | No |
| 58219802_635.tif | Cy5 | 1516 S6C1 | 3.5 h p.i. | |
| 57488502 | 57488502_532.tif | Cy3 | TQ26 1N | No |
| 57488502_635.tif | Cy5 | TQ26 2N | No |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mordecai, G.J.; Verret, F.; Highfield, A.; Schroeder, D.C. Schrödinger’s Cheshire Cat: Are Haploid Emiliania huxleyi Cells Resistant to Viral Infection or Not? Viruses 2017, 9, 51. https://doi.org/10.3390/v9030051
Mordecai GJ, Verret F, Highfield A, Schroeder DC. Schrödinger’s Cheshire Cat: Are Haploid Emiliania huxleyi Cells Resistant to Viral Infection or Not? Viruses. 2017; 9(3):51. https://doi.org/10.3390/v9030051
Chicago/Turabian StyleMordecai, Gideon J., Frederic Verret, Andrea Highfield, and Declan C. Schroeder. 2017. "Schrödinger’s Cheshire Cat: Are Haploid Emiliania huxleyi Cells Resistant to Viral Infection or Not?" Viruses 9, no. 3: 51. https://doi.org/10.3390/v9030051
APA StyleMordecai, G. J., Verret, F., Highfield, A., & Schroeder, D. C. (2017). Schrödinger’s Cheshire Cat: Are Haploid Emiliania huxleyi Cells Resistant to Viral Infection or Not? Viruses, 9(3), 51. https://doi.org/10.3390/v9030051