
Manipulation of Viral MicroRNAs as a Potential Antiviral Strategy for the Treatment of Cytomegalovirus Infection

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Viral Titers
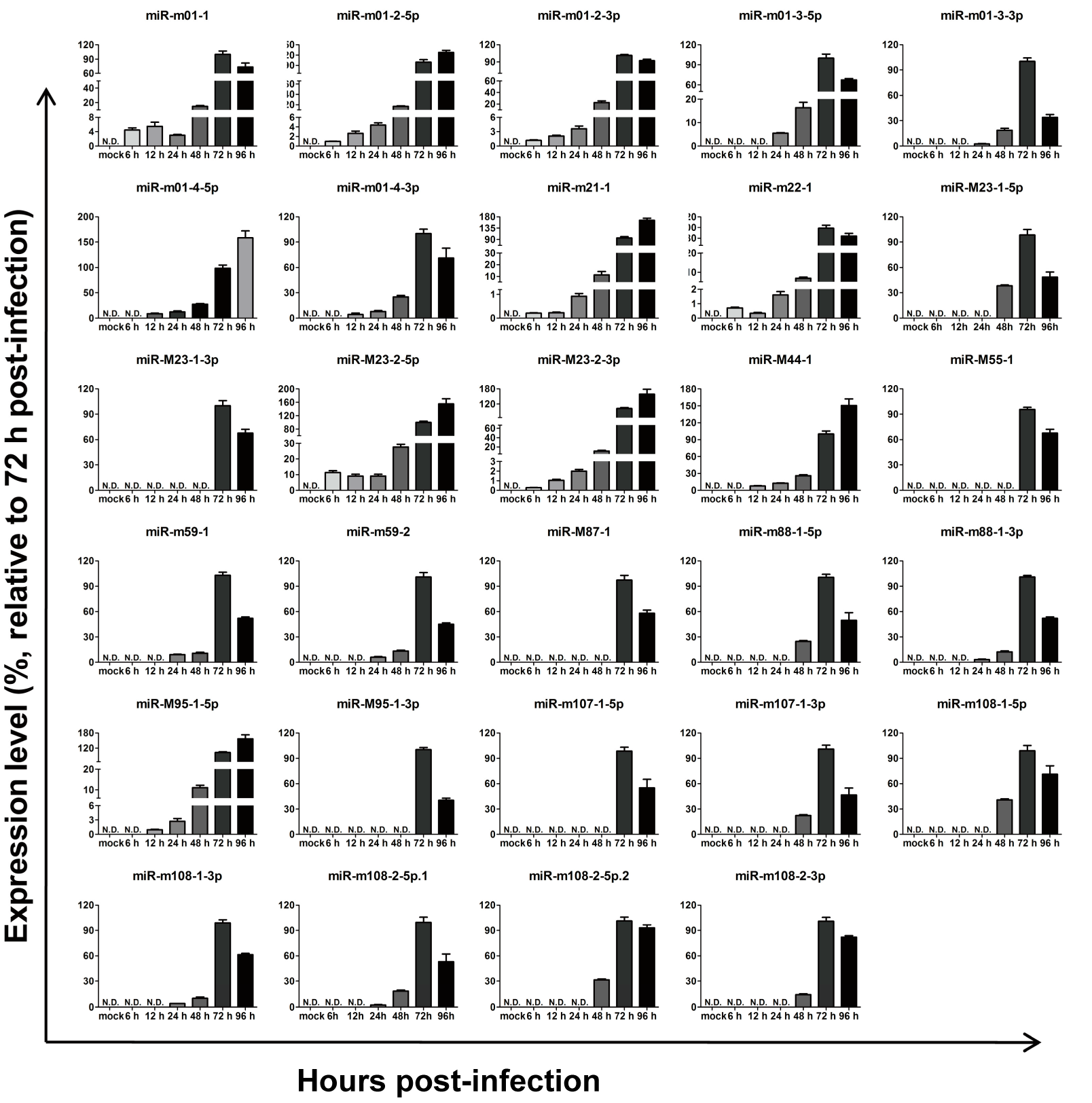
2.2. Detection of MCMV MiRNAs In Vitro
2.3. Examination of MCMV MiRNAs in Different Animal Models
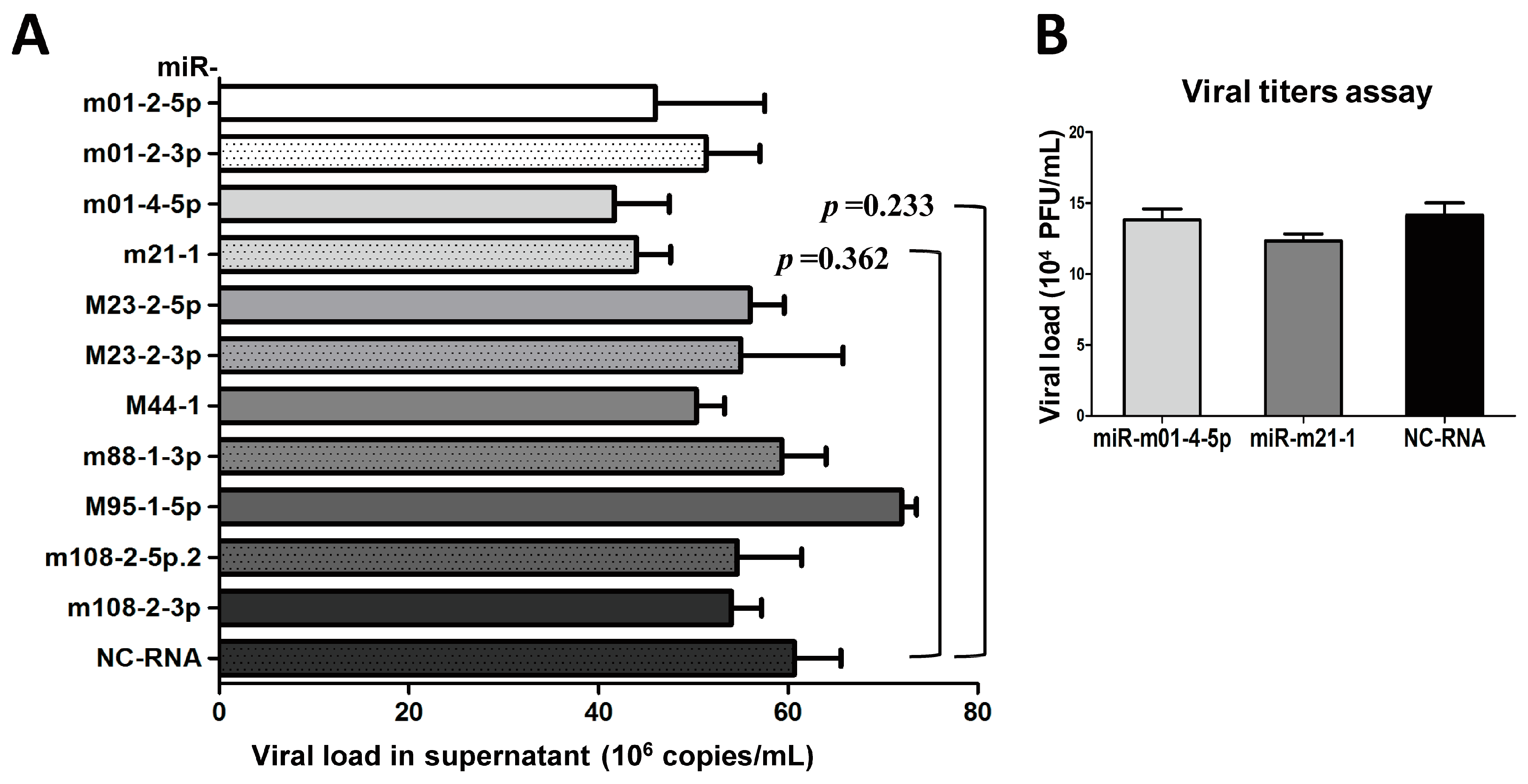
2.4. Transfection and Quantification of Viral Loads for MiRNAs in Group 1
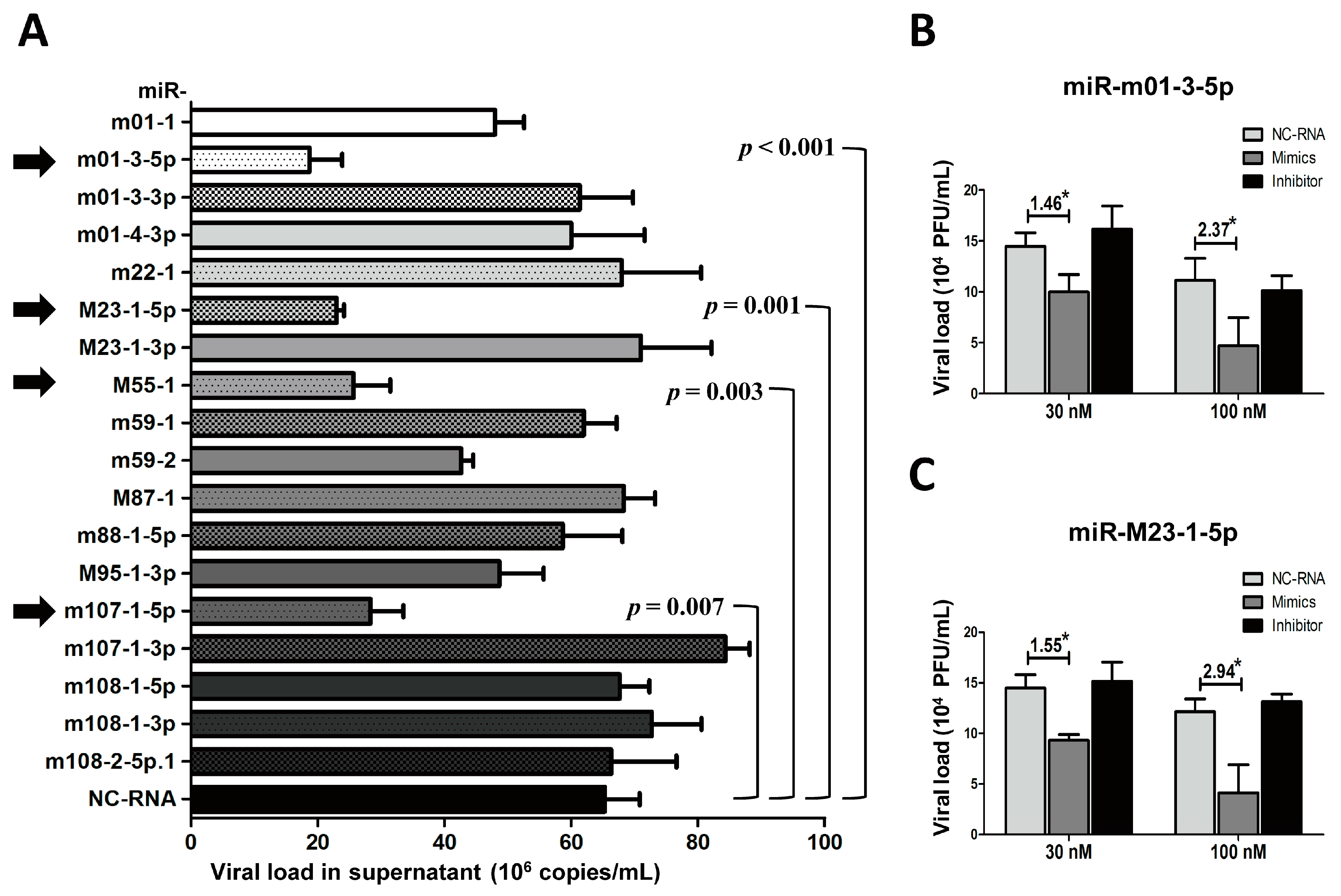
2.5. Transfection and Quantification of Viral Loads for MiRNAs in Group 2
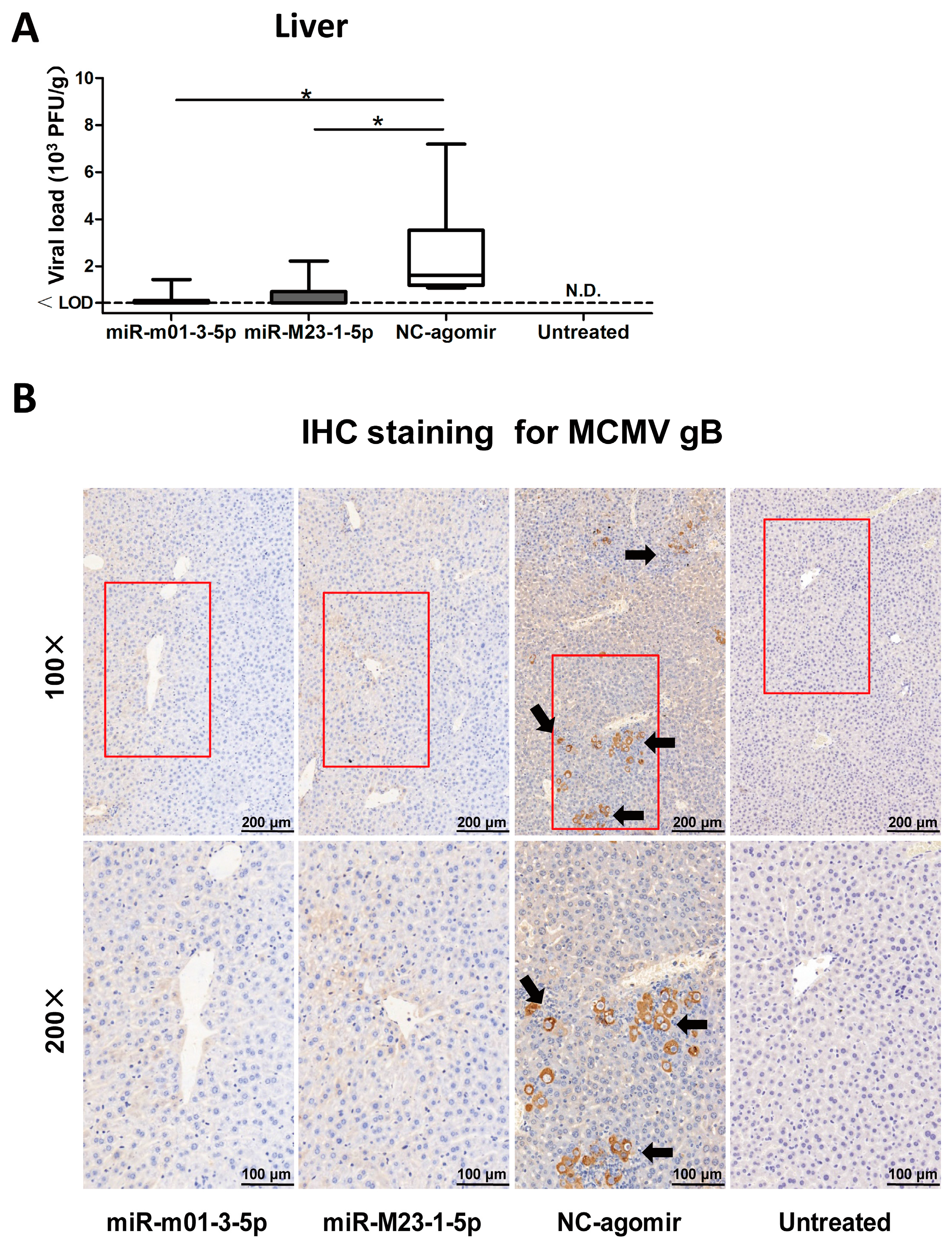
2.6. Effects of Agomir Administration on MCMV Recurrence
2.7. Immunohistochemistry
2.8. Statistical Analysis
3. Results
3.1. MCMV MiRNAExpression during Lytic Infection In Vitro
3.2. In Vivo MiRNA Expression Profiles Differ from In Vitro Profiles
3.3. The Inhibition of Group 1 Viral MiRNAs Exerts Few Effectson MCMV Replication In Vitro
3.4. The Overexpression of Several Viral miRNAs in Group 2 Inhibits MCMV Production In Vitro
3.5. MCMV MiRNA Agomirs Reduce MCMV Recurrence In Vivo
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bate, S.L.; Dollard, S.C.; Cannon, M.J. Cytomegalovirus seroprevalence in the United States: The national health and nutrition examination surveys, 1988–2004. Clin. Infect. Dis. 2010, 50, 1439–1447. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.E.; Mason, G.M.; Wills, M.R. Human cytomegalovirus immunity and immune evasion. Virus Res. 2011, 157, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Goodrich, J.M.; Bowden, R.A.; Fisher, L.; Keller, C.; Schoch, G.; Meyers, J.D. Ganciclovir prophylaxis to prevent cytomegalovirus disease after allogeneic marrow transplant. Ann. Intern. Med. 1993, 118, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Reusser, P.; Einsele, H.; Lee, J.; Volin, L.; Rovira, M.; Engelhard, D.; Finke, J.; Cordonnier, C.; Link, H.; Ljungman, P. Randomized multicenter trial of foscarnet versus ganciclovir for preemptive therapy of cytomegalovirus infection after allogeneic stem cell transplantation. Blood 2002, 99, 1159–1164. [Google Scholar] [CrossRef] [PubMed]
- Valdez, O.; Gaspar, A.; Dickson, J.; Weigert, A.; Machado, D. Cytomegalovirus infection resistant to ganciclovir in a renal transplant patient. Transplant. Proc. 2003, 35, 1081–1082. [Google Scholar] [CrossRef]
- Young, P.G.; Rubin, J.; Angarone, M.; Flaherty, J.; Penugonda, S.; Stosor, V.; Ison, M.G. Ganciclovir-resistant cytomegalovirus infection in solid organ transplant recipients: A single-center retrospective cohort study. Transpl. Infect. Dis. 2016, 18, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Isada, C.M.; Yen-Lieberman, B.; Lurain, N.S.; Schilz, R.; Kohn, D.; Longworth, D.L.; Taege, A.J.; Mossad, S.B.; Maurer, J.; Flechner, S.M.; et al. Clinical characteristics of 13 solid organ transplant recipients with ganciclovir-resistant cytomegalovirus infection. Transpl. Infect. Dis. 2002, 4, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.Z.; Pan, X.; Miao, L.F.; Ye, H.Q.; Chavanas, S.; Davrinche, C.; McVoy, M.; Luo, M.H. Comprehensive analysis of human cytomegalovirus microRNA expression during lytic and quiescent infection. PLoS ONE 2014, 9, e88531. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.R.; Li, J.Y.; Gleadle, J.M. Human cytomegalovirus encoded microRNAs: Hitting targets. Expert Rev. Anti-Infect. Ther. 2015, 13, 1469–1479. [Google Scholar] [CrossRef] [PubMed]
- Hook, L.; Hancock, M.; Landais, I.; Grabski, R.; Britt, W.; Nelson, J.A. Cytomegalovirus microRNAs. Curr. Opin. Virol. 2014, 7, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Dhuruvasan, K.; Sivasubramanian, G.; Pellett, P.E. Roles of host and viral microRNAs in human cytomegalovirus biology. Virus Res. 2011, 157, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Qi, Y.; Ma, Y.; He, R.; Ji, Y.; Sun, Z.; Ruan, Q. Down-regulation of human cytomegalovirus UL138, a novel latency-associated determinant, by hcmv-miR-UL36. J. Biosci. 2013, 38, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Stern-Ginossar, N.; Saleh, N.; Goldberg, M.D.; Prichard, M.; Wolf, D.G.; Mandelboim, O. Analysis of human cytomegalovirus-encoded microRNA activity during infection. J. Virol. 2009, 83, 10684–10693. [Google Scholar] [CrossRef] [PubMed]
- Grey, F.; Meyers, H.; White, E.A.; Spector, D.H.; Nelson, J. A human cytomegalovirus-encoded microRNA regulates expression of multiple viral genes involved in replication. PLoS Pathog. 2007, 3, e163. [Google Scholar] [CrossRef] [PubMed]
- Pavelin, J.; Reynolds, N.; Chiweshe, S.; Wu, G.; Tiribassi, R.; Grey, F. Systematic microRNA analysis identifies ATP6V0C as an essential host factor for human cytomegalovirus replication. PLoS Pathog. 2013, 9, e1003820. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.; Vanicek, J.; Robins, H.; Shenk, T.; Levine, A.J. Suppression of immediate-early viral gene expression by herpesvirus-coded microRNAs: Implications for latency. Proc. Natl. Acad. Sci. USA 2008, 105, 5453–5458. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Qi, Y.; He, R.; Huang, Y.; Liu, Z.; Ma, Y.; Guo, X.; Shao, Y.; Sun, Z.; Ruan, Q. Human cytomegalovirus microRNA miR-US25-1-5p inhibits viral replication by targeting multiple cellular genes during infection. Gene 2015, 570, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Qi, Y.; Huang, Y.; Liu, Z.; Ma, Y.; Shao, Y.; Jiang, S.; Sun, Z.; Ruan, Q. Human cytomegalovirus miR-US33-5p inhibits viral DNA synthesis and viral replication by down-regulating expression of the host Syntaxin3. FEBS Lett. 2015, 589, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.; Qi, Y.; Ma, Y.; He, R.; Ji, Y.; Sun, Z.; Ruan, Q. Over-expression of human cytomegalovirus miR-US25-2-3p downregulates eIF4A1 and inhibits HCMV replication. FEBS Lett. 2013, 587, 2266–2271. [Google Scholar] [CrossRef] [PubMed]
- Han, S.J.; Marshall, V.; Barsov, E.; Quinones, O.; Ray, A.; Labo, N.; Trivett, M.; Ott, D.; Renne, R.; Whitby, D. Kaposi’s sarcoma-associated herpesvirus microRNA single-nucleotide polymorphisms identified in clinical samples can affect microRNA processing, level of expression, and silencing activity. J. Virol. 2013, 87, 12237–12248. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Lee, S.; Kim, S.; Kim, D.; Ahn, J.H.; Ahn, K. Human cytomegalovirus clinical strain-specific microRNA miR-UL148D targets the human chemokine RANTES during infection. PLoS Pathog. 2012, 8, e1002577. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, S.; Shin, J.; Kim, Y.; Evnouchidou, I.; Kim, D.; Kim, Y.K.; Kim, Y.E.; Ahn, J.H.; Riddell, S.R.; et al. Human cytomegalovirus microRNA miR-US4-1 inhibits CD8+ T cell responses by targeting the aminopeptidase ERAP1. Nat. Immunol. 2011, 12, 984–991. [Google Scholar] [CrossRef] [PubMed]
- Nachmani, D.; Lankry, D.; Wolf, D.G.; Mandelboim, O. The human cytomegalovirus microRNA miR-UL112 acts synergistically with a cellular microRNA to escape immune elimination. Nat. Immunol. 2010, 11, 806–813. [Google Scholar] [CrossRef] [PubMed]
- Stern-Ginossar, N.; Elefant, N.; Zimmermann, A.; Wolf, D.G.; Saleh, N.; Biton, M.; Horwitz, E.; Prokocimer, Z.; Prichard, M.; Hahn, G.; et al. Host immune system gene targeting by a viral miRNA. Science 2007, 317, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Lisboa, L.F.; Egli, A.; O’Shea, D.; Asberg, A.; Hartmann, A.; Rollag, H.; Pang, X.L.; Tyrrell, D.L.; Kumar, D.; Humar, A. Hcmv-miR-UL22A-5p: A biomarker in transplantation with broad impact on host gene expression and potential immunological implications. Am. J. Transplant. 2015, 15, 1893–1902. [Google Scholar] [CrossRef] [PubMed]
- Dolken, L.; Perot, J.; Cognat, V.; Alioua, A.; John, M.; Soutschek, J.; Ruzsics, Z.; Koszinowski, U.; Voinnet, O.; Pfeffer, S. Mouse cytomegalovirus microRNAs dominate the cellular small RNA profile during lytic infection and show features of posttranscriptional regulation. J. Virol. 2007, 81, 13771–13782. [Google Scholar] [CrossRef] [PubMed]
- Gutermann, A.; Bubeck, A.; Wagner, M.; Reusch, U.; Menard, C.; Koszinowski, U.H. Strategies for the identification and analysis of viral immune-evasive genes–Cytomegalovirus as an example. Curr. Top. Microbiol. Immunol. 2002, 269, 1–22. [Google Scholar] [PubMed]
- Deng, J.; Xiao, J.; Lv, L.; Ma, P.; Song, X.; Gao, B.; Gong, F.; Zhang, Y.; Xu, J. Immunosuppressive therapy alleviates murine cytomegalovirus recurrence by reducing TNF-α post cell transplantation with lethal GVHD. Antiviral Res. 2016, 133, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Busk, P.K. A tool for design of primers for microRNA-specific quantitative RT-qPCR. BMC Bioinform. 2014, 15, 29. [Google Scholar] [CrossRef] [PubMed]
- Balcells, I.; Cirera, S.; Busk, P.K. Specific and sensitive quantitative RT-PCR of miRNAs with DNA primers. BMC Biotechnol. 2011, 11, 70. [Google Scholar] [CrossRef] [PubMed]
- Reichenstein, I.; Aizenberg, N.; Goshen, M.; Bentwich, Z.; Avni, Y.S. A novel qPCR assay for viral encoded microRNAs. J. Virol. Methods 2010, 163, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Palaniyandi, S.; Radhakrishnan, S.V.; Karlsson, F.J.; Stokes, K.Y.; Kittan, N.; Huber, E.; Hildebrandt, G.C. Murine cytomegalovirus immediate-early 1 gene expression correlates with increased GVHD after allogeneic hematopoietic cell transplantation in recipients reactivating from latent infection. PLoS ONE 2013, 8, e61841. [Google Scholar] [CrossRef] [PubMed]
- Gosselin, J.; Borgeat, P.; Flamand, L. Leukotriene B4 protects latently infected mice against murine cytomegalovirus reactivation following allogeneic transplantation. J. Immunol. 2005, 174, 1587–1593. [Google Scholar] [CrossRef] [PubMed]
- Ni, D.; Yu, H.; Zhang, W.; Gan, L.; Zhao, J.; Wang, M.; Chen, J. A mouse model of interstitial pneumonitis induced by murine cytomegalovirus infection after allogeneic skin transplantation. Biomed. Res. Int. 2013, 2013, 341387. [Google Scholar] [CrossRef] [PubMed]
- Cook, C.H.; Trgovcich, J.; Zimmerman, P.D.; Zhang, Y.; Sedmak, D.D. Lipopolysaccharide, tumor necrosis factor α, or interleukin-1β triggers reactivation of latent cytomegalovirus in immunocompetent mice. J. Virol. 2006, 80, 9151–9158. [Google Scholar] [CrossRef] [PubMed]
- Seckert, C.K.; Renzaho, A.; Tervo, H.M.; Krause, C.; Deegen, P.; Kühnapfel, B.; Reddehase, M.J.; Grzimek, N.K. Liver sinusoidal endothelial cells are a site of murine cytomegalovirus latency and reactivation. J. Virol. 2009, 83, 8869–8884. [Google Scholar] [CrossRef] [PubMed]
- Slobedman, B.; Cao, J.Z.; Avdic, S.; Webster, B.; McAllery, S.; Cheung, A.K.; Tan, J.C.; Abendroth, A. Human cytomegalovirus latent infection and associated viral gene expression. Future Microbiol. 2010, 5, 883–900. [Google Scholar] [CrossRef] [PubMed]
- Reddehase, M.J.; Podlech, J.; Grzimek, N.K. Mouse models of cytomegalovirus latency: Overview. J. Clin. Virol. 2002, 25 (Suppl. 2), S23–S36. [Google Scholar] [CrossRef]
- Meyer, C.; Grey, F.; Kreklywich, C.N.; Andoh, T.F.; Tirabassi, R.S.; Orloff, S.L.; Streblow, D.N. Cytomegalovirus microRNA expression is tissue specific and is associated with persistence. J. Virol. 2011, 85, 378–389. [Google Scholar] [CrossRef] [PubMed]
- Giladi, H.; Ketzinel-Gilad, M.; Rivkin, L.; Felig, Y.; Nussbaum, O.; Galun, E. Small interfering RNA inhibits hepatitis B virus replication in mice. Mol. Ther. 2003, 8, 769–776. [Google Scholar] [CrossRef]
- Zender, L.; Hutker, S.; Liedtke, C.; Tillmann, H.L.; Zender, S.; Mundt, B.; Waltemathe, M.; Gosling, T.; Flemming, P.; Malek, N.P.; et al. Caspase 8 small interfering RNA prevents acute liver failure in mice. Proc. Natl. Acad. Sci. USA 2003, 100, 7797–7802. [Google Scholar] [CrossRef] [PubMed]
- Grey, F.; Nelson, J. Identification and function of human cytomegalovirus microRNAs. J. Clin. Virol. 2008, 41, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Hancock, M.H.; Tirabassi, R.S.; Nelson, J.A. Rhesus cytomegalovirus encodes seventeen microRNAs that are differentially expressed in vitro and in vivo. Virology 2012, 425, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, A.A.; Rahbar, A.; Lui, W.O.; Davoudi, B.; Catrina, A.; Stragliotto, G.; Mellbin, L.; Hamsten, A.; Rydén, L.; Yaiw, K.C.; et al. Detection of circulating hcmv-miR-UL112-3p in patients with glioblastoma, rheumatoid arthritis, diabetes mellitus and healthy controls. PLoS ONE 2014, 9, e113740. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhu, J.; Zhang, W.; Chen, Y.; Zhang, K.; Popescu, L.M.; Ma, X.; Lau, W.B.; Rong, R.; Yu, X.; et al. Signature microRNA expression profile of essential hypertension and its novel link to human cytomegalovirus infection. Circulation 2011, 124, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Poole, E.; Sinclair, J. Sleepless latency of human cytomegalovirus. Med. Microbiol. Immunol. 2015, 204, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Piedade, D.; Azevedo-Pereira, J.M. The Role of microRNAs in the Pathogenesis of Herpesvirus Infection. Viruses 2016, 8, 156. [Google Scholar] [CrossRef] [PubMed]
- Meshesha, M.K.; Bentwich, Z.; Solomon, S.A.; Avni, Y.S. In vivo expression of human cytomegalovirus (HCMV) microRNAs during latency. Gene 2016, 575, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Bowman, L.J.; Melaragno, J.I.; Brennan, D.C. Letermovir for the management of cytomegalovirus infection. Expert Opin. Investig. Drugs 2017, 26, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Asberg, A.; Humar, A.; Rollag, H.; Jardine, A.G.; Mouas, H.; Pescovitz, M.D.; Sgarabotto, D.; Tuncer, M.; Noronha, I.L.; Hartmann, A. Oral valganciclovir is noninferior to intravenous ganciclovir for the treatment of cytomegalovirus disease in solid organ transplant recipients. Am. J. Transplant. 2007, 7, 2106–2113. [Google Scholar] [CrossRef] [PubMed]
- Paya, C.; Humar, A.; Dominguez, E.; Washburn, K.; Blumberg, E.; Alexander, B.; Freeman, R.; Heaton, N.; Pescovitz, M.D. Efficacy and safety of valganciclovir vs. oral ganciclovir for prevention of cytomegalovirus disease in solid organ transplant recipients. Am. J. Transplant. 2004, 4, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Winston, D.J.; Busuttil, R.W. Randomized controlled trial of oral ganciclovir versus oral acyclovir after induction with intravenous ganciclovir for long-term prophylaxis of cytomegalovirus disease in cytomegalovirus-seropositive liver transplant recipients. Transplantation 2003, 75, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Hodson, E.M.; Jones, C.A.; Webster, A.C.; Strippoli, G.F.; Barclay, P.G.; Kable, K.; Vimalachandra, D.; Craig, J.C. Antiviral medications to prevent cytomegalovirus disease and early death in recipients of solid-organ transplants: A systematic review of randomised controlled trials. Lancet 2005, 365, 2105–2115. [Google Scholar] [CrossRef]
- Gane, E.; Saliba, F.; Valdecasas, G.J.; O’Grady, J.; Pescovitz, M.D.; Lyman, S.; Robinson, C.A. Randomised trial of efficacy and safety of oral ganciclovir in the prevention of cytomegalovirus disease in liver-transplant recipients. The Oral Ganciclovir International Transplantation Study Group [corrected]. Lancet 1997, 350, 1729–1733. [Google Scholar] [CrossRef]
- Gilbert, C.; Boivin, G. Human cytomegalovirus resistance to antiviral drugs. Antimicrob. Agents Chemother. 2005, 49, 873–883. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, V.; Biron, K.K.; Talarico, C.; Stanat, S.C.; Davis, M.; Pozzi, L.M.; Coen, D.M. A point mutation in the human cytomegalovirus DNA polymerase gene confers resistance to ganciclovir and phosphonylmethoxyalkyl derivatives. Antimicrob. Agents Chemother. 1993, 37, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Jopling, C.L.; Yi, M.; Lancaster, A.M.; Lemon, S.M.; Sarnow, P. Modulation of hepatitis C virus RNA abundance by a liver-specific microRNA. Science 2005, 309, 1577–1581. [Google Scholar] [CrossRef] [PubMed]
- Thibault, P.A.; Wilson, J.A. Targeting miRNAs to treat hepatitis C virus infections and liver pathology: Inhibiting the virus and altering the host. Pharmacol. Res. 2013, 75, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Thi, E.P.; Mire, C.E.; Lee, A.C.; Geisbert, J.B.; Zhou, J.Z.; Agans, K.N.; Snead, N.M.; Deer, D.J.; Barnard, T.R.; Fenton, K.A.; et al. Lipid nanoparticle siRNA treatment of Ebola-virus-Makona-infected nonhuman primates. Nature 2015, 521, 362–365. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MiRNA Name | Accession Number | Melting Temperature (°C) | Primer F (5′–3′) | Primer R (5′–3′) | Group |
|---|---|---|---|---|---|
| miR-m01-1 | MIMAT0005533 | 71.5 | GCAGAGAGGAGAATAACGTC | GTCCAGTTTTTTTTTTTTTTTCCGT | 2 |
| miR-m01-2-5p | MIMAT0005534 | 72 | CAGGAAGAGAATCGGGTTG | GTCCAGTTTTTTTTTTTTTTTACCGT | 1 |
| miR-m01-2-3p | MIMAT0005535 | 72.5 | GCGTTCGACACGGTTTCC | GTCCAGTTTTTTTTTTTTTTTCGAAG | 1 |
| miR-m01-3-5p | MIMAT0005536 | 74 | AGCGGTGAAGCGACTGTTGC | GTCCAGTTTTTTTTTTTTTTTCGAG | 2 |
| miR-m01-3-3p | MIMAT0005537 | 75 | AGCGAGGAACGCTCGCTTCAC | GGTCCAGTTTTTTTTTTTTTTTGCC | 2 |
| miR-m01-4-5p | MIMAT0005538 | 75 | CGCAGTCCTATGCTAACAC | GGTCCAGTTTTTTTTTTTTTTTCAC | 1 |
| miR-m01-4-3p | MIMAT0005539 | 74 | AGCGCCGCGTGGTAGCAT | GGTCCAGTTTTTTTTTTTTTTTGTTCT | 2 |
| miR-m21-1 | MIMAT0005540 | 72.5 | GCAGATAGGGGACACGTTC | CAGTTTTTTTTTTTTTTTCGGCTTG | 1 |
| miR-m22-1 | MIMAT0005541 | 73.5 | CAGTTCCCGTCCGTACCGA | CCAGTTTTTTTTTTTTTTTGGCCT | 2 |
| miR-M23-1-5p | MIMAT0005542 | 73 | ACTCGGTACGGACGGGGAA | GTCCAGTTTTTTTTTTTTTTTACGGT | 2 |
| miR-M23-1-3p | MIMAT0005543 | 75 | CTCCTGCGTCGGCCCGAG | GTCCAGTTTTTTTTTTTTTTTGGC | 2 |
| miR-M23-2-5p | MIMAT0005544 | 73 | CAGTGAACGTGTCCCCTATC | GGTCCAGTTTTTTTTTTTTTTTCCA | 1 |
| miR-M23-2-3p | MIMAT0005545 | 74.5 | AGCAGATGGGGGCCTCGGT | AGTTTTTTTTTTTTTTTCCGCTTGA | 1 |
| miR-M44-1 | MIMAT0005546 | 74 | CGCAGTATCTTTTTCCAGAG | AGGTCCAGTTTTTTTTTTTTTTTACC | 1 |
| miR-M55-1 | MIMAT0005547 | 75.5 | GGTGATCGGCGTGCTA | GTCCAGTTTTTTTTTTTTTTTACGG | 2 |
| miR-m59-1 | MIMAT0005548 | 73.5 | TTAGCAGTGCCTCGACCGT | GGTCCAGTTTTTTTTTTTTTTTCTGA | 2 |
| miR-m59-2 | MIMAT0005549 | 73.5 | GCCCGAAGAGCCCTC | AGTTTTTTTTTTTTTTTGGCTCTGT | 2 |
| miR-M87-1 | MIMAT0005550 | 74.5 | CCGTCGGCAGCG | GGTCCAGTTTTTTTTTTTTTTTGCT | 2 |
| miR-m88-1-5p | MIMAT0005551 | 73 | CAGATGACCGACCCCCTGA | CCAGTTTTTTTTTTTTTTTCCGATGT | 2 |
| miR-m88-1-3p | MIMAT0005552 | 74 | AGCAGCAGAAGTCGATGT | GTCCAGTTTTTTTTTTTTTTTAGACC | 1 |
| miR-M95-1-5p | MIMAT0005553 | 73.5 | AGGGTCGTGGGCTTGTGT | CAGTTTTTTTTTTTTTTTCAAGCGA | 1 |
| miR-M95-1-3p | MIMAT0005554 | 75 | AGCGACGTCGGACCGCGA | CCAGTTTTTTTTTTTTTTTGCCGT | 2 |
| miR-m107-1-5p | MIMAT0005555 | 75.5 | CGGTCACTCGTCTCGA | CCAGTTTTTTTTTTTTTTTGGTGACT | 2 |
| miR-m107-1-3p | MIMAT0005556 | 75.5 | AGTGCTCGCGTCGAGTGACC | GGTCCAGTTTTTTTTTTTTTTTGAG | 2 |
| miR-m108-1-5p | MIMAT0005557 | 73 | CAGTCACGAGCAACCGCCC | TCCAGTTTTTTTTTTTTTTTCATTTC | 2 |
| miR-m108-1-3p | MIMAT0005558 | 74 | CAGTTTCTGACGGTGGCT | GTCCAGTTTTTTTTTTTTTTTCGAC | 2 |
| miR-m108-2-5p.1 | MIMAT0005560 | 75 | GGCGGTCACTCGAC | AGTTTTTTTTTTTTTTTCGGTGCT | 2 |
| miR-m108-2-5p.2 | MIMAT0005559 | 74 | TCACTCGTCGCGAGCGGT | GGTCCAGTTTTTTTTTTTTTTTGTGA | 1 |
| miR-m108-2-3p | MIMAT0005561 | 73.5 | GTGACTCGAGACGAGTGA | GGTCCAGTTTTTTTTTTTTTTTACCG | 1 |
| Acute Infection Group (N = 15), n (%) | Viral Reactivation Group (N = 17), n (%) | |||||
|---|---|---|---|---|---|---|
| Livers | Lungs | Blood | Livers | Lungs | Blood | |
| miR-m01-1 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| miR-m01-2-5p | 15 (100) | 15 (100) | 15 (100) | 17 (100) | 17 (100) | 15 (88) |
| miR-m01-2-3p | 15 (100) | 15 (100) | 3 (20) | 17 (100) | 14 (82) | 1 (6) |
| miR-m01-3-5p | 0 (0) | 3 (20) | 1 (7) | 0 (0) | 0 (0) | 0 (0) |
| miR-m01-3-3p | 0 (0) | 1 (7) | 0 (0) | 0 (0) | 0 (0) | 1 (6) |
| miR-m01-4-5p | 15 (100) | 15 (100) | 9 (60) | 17 (100) | 17 (100) | 7 (41) |
| miR-m01-4-3p | 3 (20) | 0 (0) | 4 (27) | 3 (18) | 0 (0) | 3 (18) |
| miR-m21-1 | 15 (100) | 15 (100) | 6 (40) | 17 (100) | 17 (100) | 9 (53) |
| miR-m22-1 | 6 (40) | 3 (20) | 3 (20) | 3 (18) | 3 (18) | 2 (12) |
| miR-M23-1-5p | 2 (13) | 1 (7) | 0 (0) | 3 (18) | 0 (0) | 1 (6) |
| miR-M23-1-3p | 0 (0) | 0 (0) | 1 (7) | 0 (0) | 2 (12) | 0 (0) |
| miR-M23-2-5p | 14 (93) | 15 (100) | 8 (53) | 12 (71) | 14 (82) | 9 (53) |
| miR-M23-2-3p | 15 (100) | 15 (100) | 8 (53) | 17 (100) | 14 (82) | 13 (76) |
| miR-M44-1 | 15 (100) | 12 (80) | 2 (13) | 17 (100) | 14 (82) | 4 (24) |
| miR-M55-1 | 1 (7) | 1 (7) | 0 (0) | 0 (0) | 0 (0) | 2 (12) |
| miR-m59-1 | 0 (0) | 0 (0) | 1 (7) | 2 (12) | 0 (0) | 0 (0) |
| miR-m59-2 | 3 (20) | 0 (0) | 0 (0) | 0 (0) | 3 (18) | 1 (6) |
| miR-M87-1 | 0 (0) | 2 (13) | 1 (7) | 0 (0) | 0 (0) | 0 (0) |
| miR-m88-1-5p | 1 (7) | 0 (0) | 0 (0) | 0 (0) | 3 (18) | 0 (0) |
| miR-m88-1-3p | 15 (100) | 15 (100) | 8 (53) | 17 (100) | 17 (100) | 12 (71) |
| miR-M95-1-5p | 15 (100) | 15 (100) | 3 (20) | 17 (100) | 17 (100) | 7 (41) |
| miR-M95-1-3p | 0 (0) | 2 (13) | 0 (0) | 0 (0) | 2 (12) | 2 (12) |
| miR-m107-1-5p | 3 (20) | 0 (0) | 1 (7) | 0 (0) | 0 (0) | 1 (6) |
| miR-m107-1-3p | 0 (0) | 1 (7) | 3 (20) | 0 (0) | 0 (0) | 0 (0) |
| miR-m108-1-5p | 3 (20) | 0 (0) | 0 (0) | 2 (12) | 2 (12) | 0 (0) |
| miR-m108-1-3p | 3 (20) | 2 (13) | 4 (27) | 1 (6) | 1 (6) | 1 (6) |
| miR-m108-2-5p.1 | 2 (13) | 3 (20) | 1 (7) | 0 (0) | 0 (0) | 0 (0) |
| miR-m108-2-5p.2 | 12 (80) | 9 (60) | 10 (67) | 17 (100) | 12 (71) | 11 (65) |
| miR-m108-2-3p | 12 (80) | 15 (100) | 7 (47) | 17 (100) | 17 (100) | 15 (88) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, J.; Xiao, J.; Ma, P.; Gao, B.; Gong, F.; Lv, L.; Zhang, Y.; Xu, J. Manipulation of Viral MicroRNAs as a Potential Antiviral Strategy for the Treatment of Cytomegalovirus Infection. Viruses 2017, 9, 118. https://doi.org/10.3390/v9050118
Deng J, Xiao J, Ma P, Gao B, Gong F, Lv L, Zhang Y, Xu J. Manipulation of Viral MicroRNAs as a Potential Antiviral Strategy for the Treatment of Cytomegalovirus Infection. Viruses. 2017; 9(5):118. https://doi.org/10.3390/v9050118
Chicago/Turabian StyleDeng, Jiang, Jun Xiao, Ping Ma, Bo Gao, Feng Gong, Liping Lv, Yanyu Zhang, and Jinbo Xu. 2017. "Manipulation of Viral MicroRNAs as a Potential Antiviral Strategy for the Treatment of Cytomegalovirus Infection" Viruses 9, no. 5: 118. https://doi.org/10.3390/v9050118
APA StyleDeng, J., Xiao, J., Ma, P., Gao, B., Gong, F., Lv, L., Zhang, Y., & Xu, J. (2017). Manipulation of Viral MicroRNAs as a Potential Antiviral Strategy for the Treatment of Cytomegalovirus Infection. Viruses, 9(5), 118. https://doi.org/10.3390/v9050118