
Using Next Generation Sequencing to Identify and Quantify the Genetic Composition of Resistance-Breaking Commercial Isolates of Cydia pomonella Granulovirus


 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Viruses
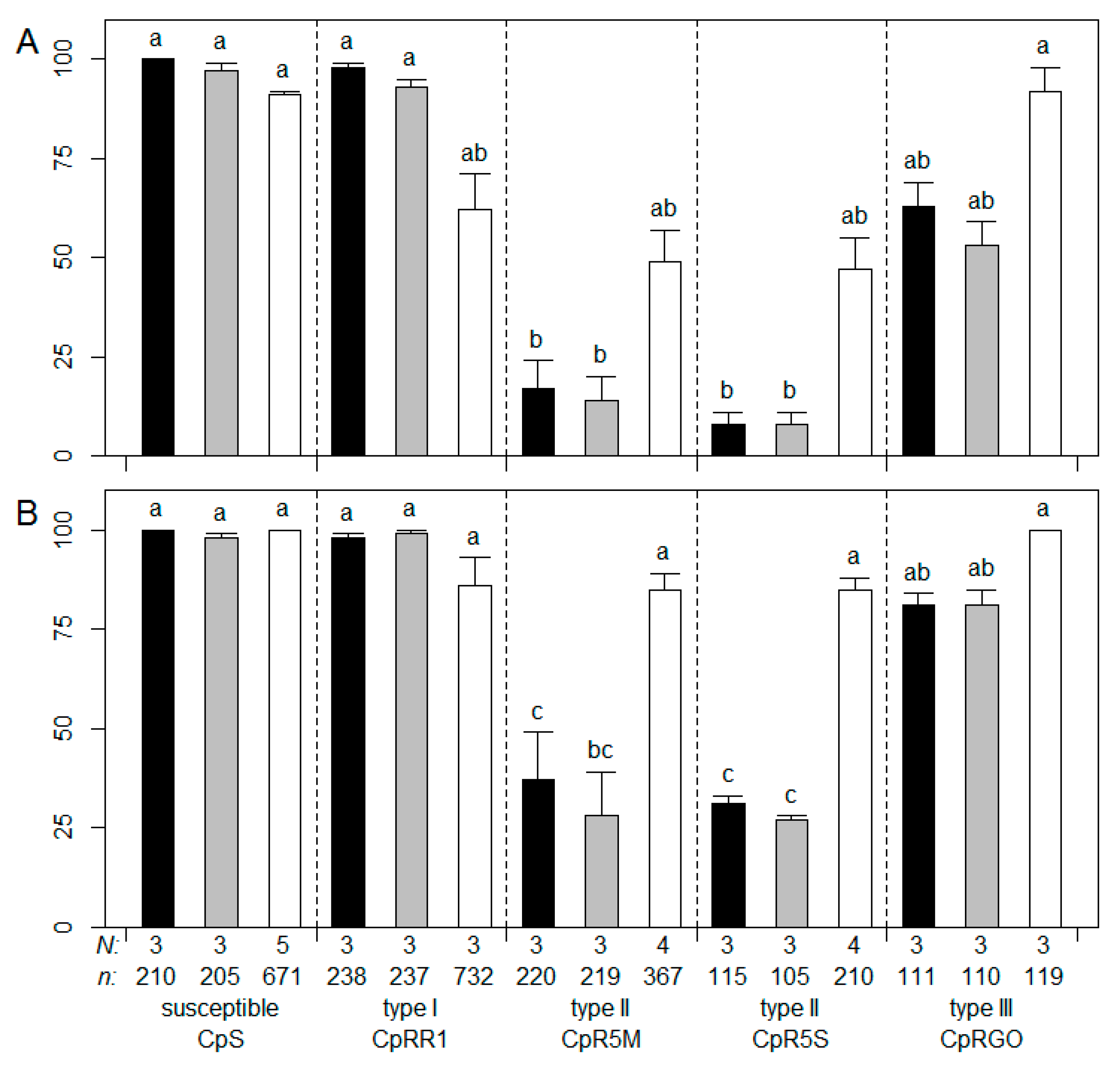
2.3. Resistance Testing
2.4. Alignment of CpGV Isolates
2.5. DNA Extraction & Whole Genome Sequencing
2.6. Detection of Single Nucleotide Polymorphisms
3. Results
3.1. Resistance Testing with Commercial CpGV Products
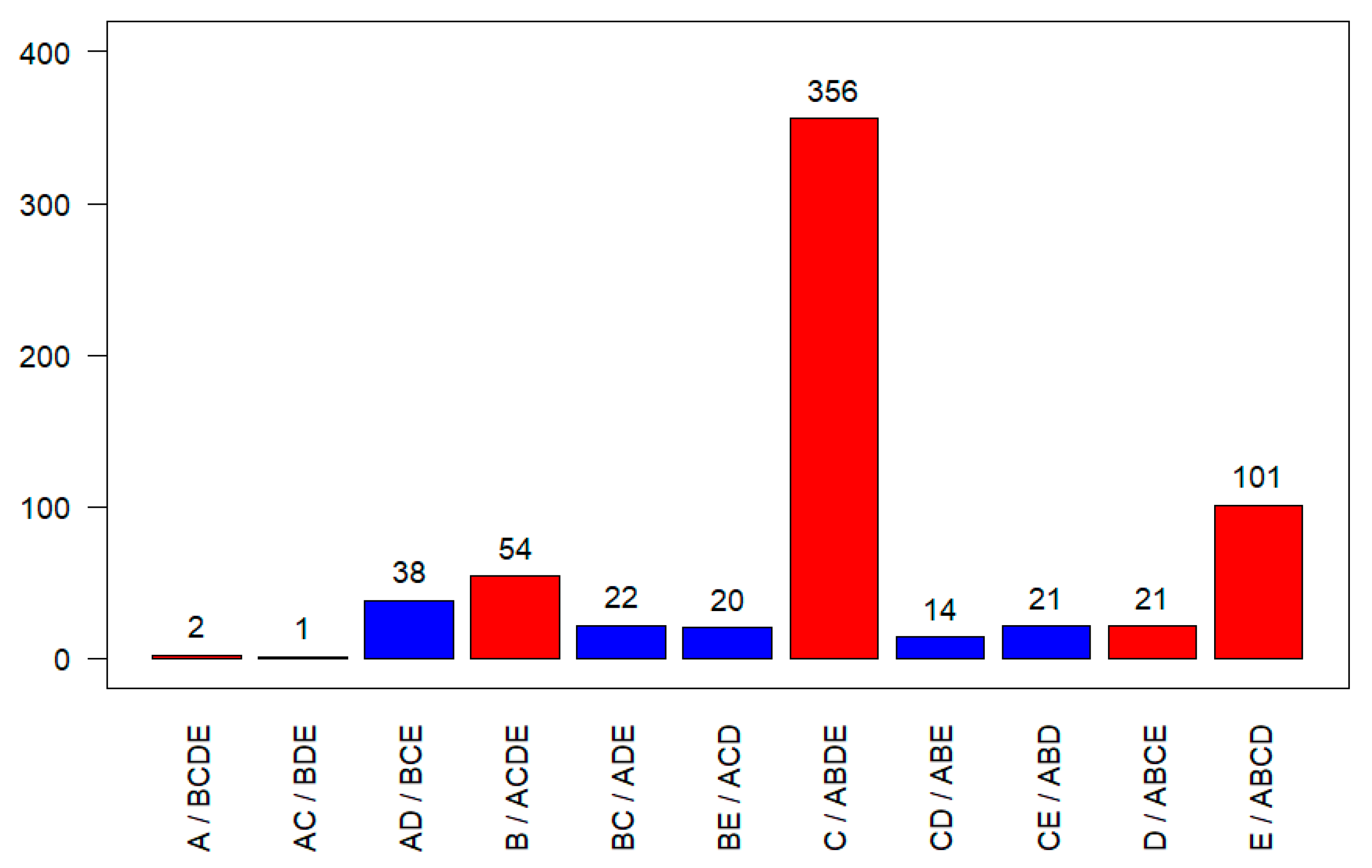
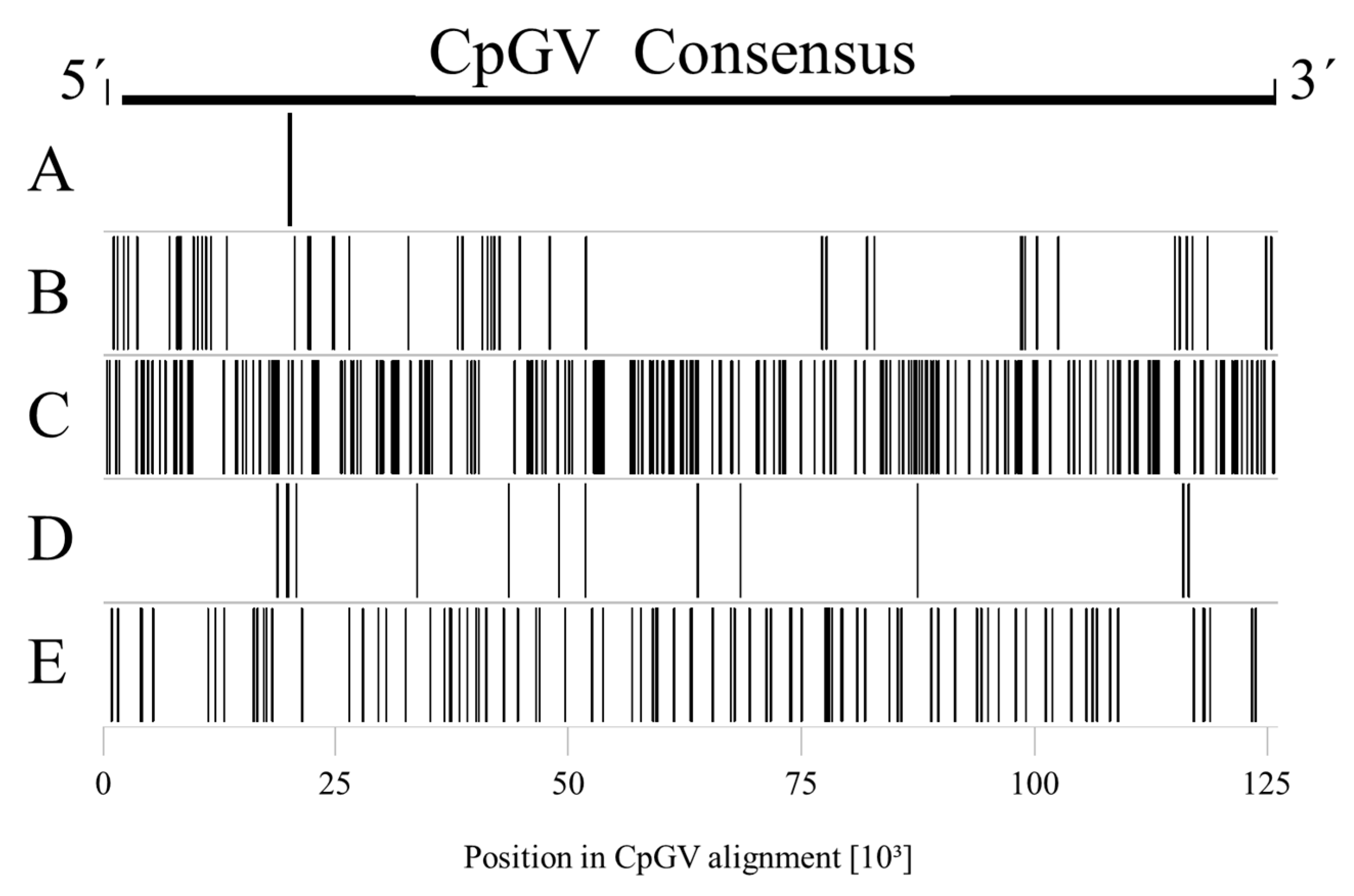
3.2. Detection of Genome Type Specific SNPs in the CpGV Alignment
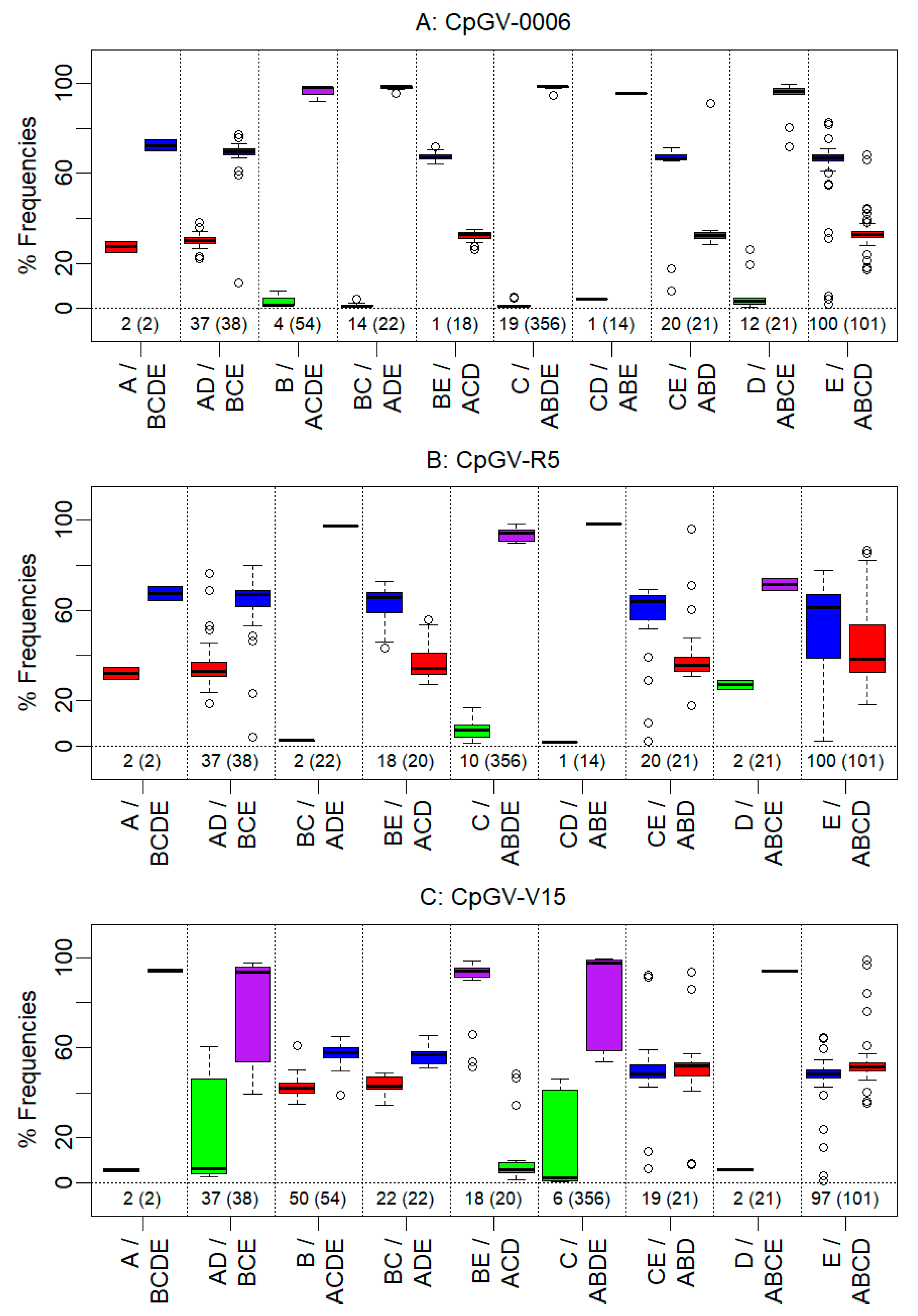
3.3. Genome Group Composition of CpGV-0006, CpGV-R5 and CpGV-V15
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Herniou, E.A.; Arif, B.M.; Bonning, B.C.; Theilmann, D.A.; Blissard, G.W.; Becnel, J.J.; Jehle, J.A.; Harrison, R.L. Baculoviridae. In Virus Taxonomy; Elsevier: Oxford, UK, 2012; Volume 9, pp. 163–173. [Google Scholar]
- Tanada, Y. A granulosis virus of the codling moth, Carpocapsa pomonella (Linnaeus) (Olethreutidae, Lepidoptera). J. Invertebr. Pathol. 1964, 6, 378–380. [Google Scholar]
- Tanada, Y.; Leutenegger, R. Histopathology of a Granulosis-Virus Disease of the Codling Moth, Carpocapsa pomonella. J. Invertebr. Pathol. 1968, 10, 39–47. [Google Scholar] [CrossRef]
- Gebhardt, M.M.; Eberle, K.E.; Radtke, P.; Jehle, J.A. Baculovirus resistance in codling moth is virus isolate-dependent and the consequence of a mutation in viral gene pe38. Proc. Natl. Acad. Sci. USA 2014, 111, 15711–15716. [Google Scholar] [CrossRef] [PubMed]
- Wormleaton, S.L.; Winstanley, D. Phylogenetic analysis of conserved genes within the ecdysteroid UDP-glucosyltransferase gene region of the slow-killing Adoxophyes orana granulovirus. J. Gen. Virol. 2001, 82, 2295–2305. [Google Scholar] [CrossRef] [PubMed]
- Wennmann, J.T.; Radtke, P.; Eberle, K.E.; Gueli Alletti, G.; Jehle, J.A. Deciphering single nucleotide polymorphisms and evolutionary trends in isolates of the Cydia pomonella granulovirus. Viruses 2017, 9, 227. [Google Scholar] [CrossRef] [PubMed]
- Eberle, K.E.; Sayed, S.; Rezapanah, M.; Shojai-Estabragh, S.; Jehle, J.A. Diversity and evolution of the Cydia pomonella granulovirus. J. Gen. Virol. 2009, 90, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Crook, N.E.; Spencer, R.A.; Payne, C.C.; Leisy, D.J. Variation in Cydia pomonella granulosis virus isolates and physical maps of the DNA from three variants. J. Gen. Virol. 1985, 66, 2423–2430. [Google Scholar] [CrossRef]
- Rezapanah, M.; Shojai-Estabragh, S.; Huber, J.; Jehle, J.A. Molecular and biological characterization of new isolates of Cydia pomonella granulovirus from Iran. J. Pest Sci. 2008, 81, 187–191. [Google Scholar] [CrossRef]
- Garavaglia, M.J.; Miele, S.A.; Iserte, J.A.; Belaich, M.N.; Ghiringhelli, P.D. The ac53, ac78, ac101, and ac103 genes are newly discovered core genes in the family Baculoviridae. J. Virol. 2012, 86, 12069–12079. [Google Scholar] [CrossRef] [PubMed]
- Miele, S.A.; Garavaglia, M.J.; Belaich, M.N.; Ghiringhelli, P.D. Baculovirus: Molecular insights on their diversity and conservation. Int. J. Evol. Biol. 2011, 2011, 379424. [Google Scholar] [CrossRef] [PubMed]
- Lacey, L.A.; Arthurs, S.P.; Headrick, H. Comparative activity of the codling moth granulovirus against Grapholita molesta and Cydia pomonella (Lepidoptera: Tortricidae). J. Entomol. Soc. B. C. 2005, 102, 79–80. [Google Scholar]
- Zingg, D.; Kraaz, I.; Wandeler, H.; Züger, M. Madex Twin, a new Cydia pomonella granulovirus isolate for the control of both codling moth Cydia pomonella and oriental fruit moth Grapholita molesta. In Proceedings of the 15th International Conference on Organic Fruit-Growing, Ecofruit, Hohenheim, Germany, 20–22 February 2012; pp. 187–193. [Google Scholar]
- Asser-Kaiser, S.; Fritsch, E.; Undorf-Spahn, K.; Kienzle, J.; Eberle, K.E.; Gund, N.A.; Reineke, A.; Zebitz, C.P.; Heckel, D.G.; Huber, J.; et al. Rapid emergence of baculovirus resistance in codling moth due to dominant, sex-linked inheritance. Science 2007, 317, 1916–1918. [Google Scholar] [CrossRef] [PubMed]
- Briese, D.T. Genetic basis for resistance to a granulosis virus in the potato moth Phthorimaea operculella. J. Invertebr. Pathol. 1982, 39, 215–218. [Google Scholar] [CrossRef]
- Briese, D.T.; Mende, H.A. Selection for increased resistance to a granulosis virus in the potato moth, Phthorimaea operculella (Zeller) (Lepidoptera, Gelichiidae). Bull. Entomol. Res. 1983, 73, 1–9. [Google Scholar] [CrossRef]
- Abot, A.R.; Moscardi, F.; Fuxa, J.R.; Sosa-Gomez, D.R.; Richter, A.R. Development of resistance by Anticarsia gemmatalis from Brazil and the United States to a nuclear polyhedrosis virus under laboratory selection pressure. Biol. Control 1996, 7, 126–130. [Google Scholar] [CrossRef]
- Milks, M.L.; Myers, J.H. The development of larval resistance to a nucleopolyhedrovirus is not accompanied by an increased virulence in the virus. Evol. Ecol. 2000, 14, 654–664. [Google Scholar] [CrossRef]
- Milks, M.L.; Theilmann, D.A. Serial Selection for resistance to a wild-type and to a genetically modified nucleopolyhedrovirus in Trichoplusia ni. Biol. Control 2000, 19, 283–289. [Google Scholar] [CrossRef]
- Iwata, K.; Haas-Stapleton, E.J.; Kunimi, Y.; Inoue, M.N.; Nakai, M. Midgut based resistance to oral infection by a nucleopolyhedrovirus in the laboratory-selected strain of the small tea tortrix, Adoxophyses honmai (Lepidoptera: Totridicidae). J. Gen. Virol. 2017, 98, 296–304. [Google Scholar] [PubMed]
- Nakai, M.; Takahashi, K.; Iwata, K.; Koyanagi, J.; Ookuma, A.; Takatsuka, J.; Okuno, S.; Kunimi, Y. Acquired resistance to a nucleopolyhedrovirus in the smaller tea tortrix Adoxophyes honmai (Lepidoptera: Tortricidae) after selection by serial viral administration. J. Invertebr. Pathol. 2017, 145, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Fritsch, E.; Undorf-Spahn, K.; Kienzle, J.; Zebitz, C.P.; Huber, J. Apfelwickler-Granulovirus: Erste Hinweise auf Unterschiede in der Empfindlichkeit lokaler Apfelwickler-Populationen. Nachrichtenbl. Deut. Pflanzenschutzd. 2005, 57, 29–34. [Google Scholar]
- Sauphanor, B.; Berling, M.; Toubon, J.-F.; Reyes, M.; Delnatte, J. Carpocapse des pommes cas de resistance aux virus de la granulose dans le sud-est. Phytoma 2006, 590, 24–27. [Google Scholar]
- Schmitt, A.; Bisutti, I.L.; Ladurner, E.; Benuzzi, M.; Sauphanor, B.; Kienzle, J.; Zingg, D.; Undorf-Spahn, K.; Fritsch, E.; Huber, J.; et al. The occurrence and distribution of resistance of codling moth to Cydia pomonella granulovirus in Europe. J. Appl. Entomol. 2013, 137, 641–649. [Google Scholar] [CrossRef]
- Asser-Kaiser, S.; Heckel, D.G.; Jehle, J.A. Sex linkage of CpGV resistance in a heterogeneous field strain of the codling moth Cydia pomonella (L.). J. Invertebr. Pathol. 2010, 103, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Asser-Kaiser, S.; Radtke, P.; El-Salamouny, S.; Winstanley, D.; Jehle, J.A. Baculovirus resistance in codling moth (Cydia pomonella L.) caused by early block of virus replication. Virology 2011, 410, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Zichová, T.; Stará, J.; Kundu, J.K.; Eberle, K.E.; Jehle, J.A. Resistance to Cydia pomonella granulovirus follows a geographically widely distributed inheritance type within Europe. BioControl 2013, 58, 525–534. [Google Scholar] [CrossRef]
- Berling, M.; Blachere-Lopez, C.; Soubabere, O.; Lery, X.; Bonhomme, A.; Sauphanor, B.; Lopez-Ferber, M. Cydia pomonella granulovirus genotypes overcome virus resistance in the codling moth and improve virus efficiency by selection against resistant hosts. Appl. Environ. Microbiol. 2009, 75, 925–930. [Google Scholar] [CrossRef] [PubMed]
- Graillot, B.; Blachere-Lopez, C.; Besse, S.; Siegwart, M.; Lopez-Ferber, M. Variations in the susceptibility to CpGV in populations of the codling moth, Cydia pomonella. IOBC Bull. 2013, 90, 149–153. [Google Scholar]
- Berling, M.; Sauphanor, B.; Bonhomme, A.; Siegwart, M.; Lopez-Ferber, M. A single sex-linked dominant gene does not fully explain the codling moth’s resistance to granulovirus. Pest Manag. Sci. 2013, 69, 1261–1266. [Google Scholar] [CrossRef] [PubMed]
- Jehle, J.A.; Schulze-Bopp, S.; Undorf-Spahn, K.; Fritsch, E. Evidence for a second type of resistance against Cydia pomonella granulovirus in field populations of codling moths. Appl. Environ. Microbiol. 2017, 83, e02330-16. [Google Scholar] [CrossRef] [PubMed]
- Eberle, K.E.; Asser-Kaiser, S.; Sayed, S.M.; Nguyen, H.T.; Jehle, J.A. Overcoming the resistance of codling moth against conventional Cydia pomonella granulovirus (CpGV-M) by a new isolate CpGV-I12. J. Invertebr. Pathol. 2008, 98, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Sauer, A.J.; Fritsch, E.; Undorf-Spahn, K.; Nguyen, H.T.; Frantisek, M.; Heckel, D.G.; Jehle, J.A. Novel resistance to Cydia pomonella granulovirus (CpGV) in codling moth shows autosomal and dominant inheritance and confers cross-resistance to different CpGV genome groups. PLoS ONE 2017, 12, e0179157. [Google Scholar] [CrossRef] [PubMed]
- Sauer, A.J.; Gebhardt, M.M.; Fritsch, E.; Undorf-Spahn, K.; Iwata, K.; Kleespies, R.G.; Nakai, M.; Jehle, J.A. Cross-resistance of coding moth against different isolates of Cydia pomonella granulovirus is caused by two different but genetically linked resistance mechanisms. J. Virol. 2017, in press. [Google Scholar]
- Sauer, A.J.; Schulze-Bopp, S.; Fritsch, E.; Undorf-Spahn, K.; Jehle, J.A. A third type of resistance of codling moth against Cydia pomonella granulovirus (CpGV) shows a mixture of a Z-linked and autosomal inheritance pattern. Appl. Environ. Microbiol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Zingg, D. Madex Plus and Madex I12 overcome virus resistance of codling moth. In Ecofruit, Proceedings of the 13th International Conference on Cultivation Technique and Phytsopathological Problems in Organic Fruit-Growing, Weinsberg, Germany, 18–20 February 2008; Föko e.V.: Weinsberg, Germany, 2010; pp. 256–260. [Google Scholar]
- Brand, G.; Zingg, D.; Querrioux, A. Virus de la granulose du carpocapse: Le bal des isolats. Phytoma 2017, 702, 47–49. [Google Scholar]
- European Union. Regulation (EC) No 1107/2009 of the European Parliament and of the Council of 21 October 2009 Concerning the Placing of Plant Protection Products on the Market and Repealing Council Directives 79/117/EEC and 91/414/EEC. OJ NO l309/1. Off. J. Eur. Union 2009, L 309/1, 52–101. [Google Scholar]
- European Union. Commission Regulation (EU) No 283/2013 of 1 March 2013 setting out the data requirements for active substances, in accordance with regulation (EC) no 1107/2009 of the European Parliament and of the Council concerning the placing of plant protection products on the market. Off. J. Eur. Union 2013, L 93/1, 44–127. [Google Scholar]
- Ivaldi-Sender, C. A simple technique for the permanent rearing of the oriental fruit moth, Grapholita molesta (Lepidoptera. Tortricidae) on an artificial medium. Techniques simples pour un elevage permanent de la tordeuse orientale, Grapholita molesta (Lepidoptera: Tortricidae) sur milieu artificiel. Ann. Zool. Ecol. Anim. 1974, 6, 337–343. [Google Scholar]
- Zingg, D.; Züger, M.; Bollhalder, F.; Andermatt, M. Use of resistance overcoming CpGV isolates and CpGV resistance situation of the codling moth in Europe seven years after the first discovery of resistance to CpGV-M. IOBC-WPRS Bull. 2011, 66, 401–404. [Google Scholar]
- Graillot, B.; Berling, M.; Blachere-Lopez, C.; Siegwart, M.; Besse, S.; Lopez-Ferber, M. Progressive Adaptation of a CpGV Isolate to Codling Moth Populations Resistant to CpGV-M. Viruses 2014, 6, 5135–5144. [Google Scholar] [CrossRef] [PubMed]
- Jehle, J.A.; Backhaus, H.; Fritsch, E.; Huber, J. Physical map of the Cryptophlebia leucotreta granulosis virus genome and its relationship to the genome of Cydia pomoneila granulosis virus. J. Gen. Virol. 1992, 73, 1621–1626. [Google Scholar] [CrossRef] [PubMed]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Arends, H.M.; Jehle, J.A. Sequence analysis and quantification of transposase cDNAs of transposon tcp3.2 in Cydia pomonella larvae. Arch. Insect Biochem. Physiol. 2006, 63, 135–145. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, D.R.; Miller, L.K.; Luckov, V.A. Baculovirus Expression Vectors: A Laboratory Manual; Oxford University Press: New York, NY, USA, 1994. [Google Scholar]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Gordon, A. Fastx Toolkit. 2009. Available online: http://hannonlab.cshl.edu/fastx_toolkit/ (accessed on 1 September 2016).
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Chateigner, A.; Bezier, A.; Labrousse, C.; Jiolle, D.; Barbe, V.; Herniou, E.A. Ultra deep sequencing of a baculovirus population reveals widespread genomic variations. Viruses 2015, 7, 3625–3646. [Google Scholar] [CrossRef] [PubMed]
- Graillot, B.; Bayle, S.; Blachere-Lopez, C.; Besse, S.; Siegwart, M.; Lopez-Ferber, M. Biological characteristics of experimental genotype mixtures of Cydia pomonella granulovirus (CpGV): Ability to control susceptible and resistant pest populations. Viruses 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Clavijo, G.; Williams, T.; Munoz, D.; Caballero, P.; Lopez-Ferber, M. Mixed genotype transmission bodies and virions contribute to the maintenance of diversity in an insect virus. Proc. R. Soc. B 2009, 277, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Ferber, M.; Simon, O.; Williams, T.; Caballero, P. Defective or effective? Mutualistic interactions between virus genotypes. Proc. R. Soc. B 2003, 270, 2249–2255. [Google Scholar] [CrossRef] [PubMed]
- Simon, O.; Williams, T.; Lopez-Ferber, M.; Caballero, P. Functional importance of deletion mutant genotypes in an insect nucleopolyhedrovirus population. Appl. Environ. Microbiol. 2005, 71, 4254–4262. [Google Scholar] [CrossRef] [PubMed]
- Simon, O.; Williams, T.; Possee, R.D.; Lopez-Ferber, M.; Caballero, P. Stability of a Spodoptera frugiperda nucleopolyhedrovirus deletion recombinant during serial passage in insects. Appl. Environ. Microbiol. 2010, 76, 803–809. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CpGV Isolate | Reads Pairs ≥ Q30 | Number and Percentage of Read Pairs Assembled to CpGV Consensus | Number and Percentage of Read Pairs Not Assembled to CpGV Consensus | Mean Coverage ± SD |
|---|---|---|---|---|
| 0006 | 736,927 | 729,391 (99%) | 7536 (1%) | 1690 ± 515 |
| R5 | 1,517,362 | 240,501 (16%) | 1,273,861 (84%) * | 565 ± 180 |
| V15 | 1,232,807 | 1,231,217 (>99%) | 1590 (<1%) | 2848 ± 557 |
| CpGV Isolate | Genome Groups | Combined Group | Data Size (n) | Mean (95%-CL) (%) | Median (5–95%) (%) | Weight. Median (%) |
|---|---|---|---|---|---|---|
| 0006 | A, AD, ACD, ABD, ABCD | A | 160 | 33 (32–34) | 32 (27–38) | 32 |
| BCDE; BCE; BE, CE, E | E | 160 | 65 (63–67) | 67 (55–72) | 68 | |
| B, BC, C, CD, D | BCD | 50 | 3 (2–4) | 1 (0.6–7) | 0 | |
| R5 | A, AD, ABD, ACD, ABCD | A | 177 | 41 (39–43) | 36 (26–77) | 36 |
| BCDE, BCE, BE, CE, E | E | 177 | 57 (54–60) | 64 (18–74) | 64 | |
| BC, C, CD, D | CD 1 | 15 | 9 (5–13) | 5 (1–26) | 0 | |
| V15 | ABD, ABCD, B, BC | B | 188 | 48 (47–50) | 49 (36–57) | 48 * |
| ACDE, ADE, CE, E | E | 188 | 52 (51–54) | 51 (43–64) | 50 * | |
| A, AD, ACD, C, D | ACD | 65 | 15 (11–20) | 6 (2–50) | 2 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alletti, G.G.; Sauer, A.J.; Weihrauch, B.; Fritsch, E.; Undorf-Spahn, K.; Wennmann, J.T.; Jehle, J.A. Using Next Generation Sequencing to Identify and Quantify the Genetic Composition of Resistance-Breaking Commercial Isolates of Cydia pomonella Granulovirus. Viruses 2017, 9, 250. https://doi.org/10.3390/v9090250
Alletti GG, Sauer AJ, Weihrauch B, Fritsch E, Undorf-Spahn K, Wennmann JT, Jehle JA. Using Next Generation Sequencing to Identify and Quantify the Genetic Composition of Resistance-Breaking Commercial Isolates of Cydia pomonella Granulovirus. Viruses. 2017; 9(9):250. https://doi.org/10.3390/v9090250
Chicago/Turabian StyleAlletti, Gianpiero Gueli, Annette J. Sauer, Birgit Weihrauch, Eva Fritsch, Karin Undorf-Spahn, Jörg T. Wennmann, and Johannes A. Jehle. 2017. "Using Next Generation Sequencing to Identify and Quantify the Genetic Composition of Resistance-Breaking Commercial Isolates of Cydia pomonella Granulovirus" Viruses 9, no. 9: 250. https://doi.org/10.3390/v9090250
APA StyleAlletti, G. G., Sauer, A. J., Weihrauch, B., Fritsch, E., Undorf-Spahn, K., Wennmann, J. T., & Jehle, J. A. (2017). Using Next Generation Sequencing to Identify and Quantify the Genetic Composition of Resistance-Breaking Commercial Isolates of Cydia pomonella Granulovirus. Viruses, 9(9), 250. https://doi.org/10.3390/v9090250