
A Preliminary Study about the Role of Reactive Oxygen Species and Inflammatory Process after COVID-19 Vaccination and COVID-19 Disease

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Ethical Statement
2.3. Analysis of Antibodies
2.4. Analysis of ROS Levels
2.5. Statistical Analysis
3. Results
3.1. First Study
3.2. Second Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weiss, S.R.; Navas-Martin, S. Coronavirus Pathogenesis and the Emerging Pathogen Severe Acute Respiratory Syndrome Coronavirus. Microbiol. Mol. Biol. Rev. 2005, 69, 635–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsang, H.F.; Chan, L.W.C.; Cho, W.C.S.; Yu, A.C.S.; Yim, A.K.Y.; Chan, A.K.C.; Ng, L.P.W.; Wong, Y.K.E.; Pei, X.M.; Li, M.J.W.; et al. An update on COVID-19 pandemic: The epidemiology, pathogenesis, prevention and treatment strategies. Expert Rev. Anti-Infect. Ther. 2020, 19, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Wölfel, R.; Corman, V.M.; Guggemos, W.; Seilmaier, M.; Zange, S.; Müller, M.A.; Niemeyer, D.; Jones, T.C.; Vollmar, P.; Rothe, C.; et al. Virological assessment of hospitalized patients with COVID-2019. Nature 2020, 581, 465–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Leary, V.B.; Ovsepian, S.V. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2). Trends Genet. 2020, 36, 892–893. [Google Scholar] [CrossRef] [PubMed]
- SoRelle, J.A.; Frame, I.; Falcon, A.; Jacob, J.; Wagenfuehr, J.; Mitui, M.; Park, J.Y.; Filkins, L. Clinical Validation of a SARS-CoV-2 Real-Time Reverse Transcription PCR Assay Targeting the Nucleocapsid Gene. J. Appl. Lab. Med. 2020, 5, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, W.; Zhang, Q.; Xu, K.; Ye, G.; Wu, W.; Sun, Z.; Liu, F.; Wu, K.; Zhong, B.; et al. RNA based mNGS approach identifies a novel human coronavirus from two individual pneumonia cases in 2019 Wuhan outbreak. Emerg. Microbes Infect. 2020, 9, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.-Y.; Wang, P.; Mok, B.W.-Y.; Zhang, A.J.; Chu, H.; Lee, A.C.-Y.; Deng, S.; Chen, P.; Chan, K.-H.; Song, W.; et al. Attenuated SARS-CoV-2 variants with deletions at the S1/S2 junction. Emerg. Microbes Infect. 2020, 9, 837–842. [Google Scholar] [CrossRef]
- Parlakpinar, H.; Gunata, M. SARS-CoV-2 (COVID-19): Cellular and biochemical properties and pharmacological insights into new therapeutic developments. Cell Biochem. Funct. 2021, 39, 10–28. [Google Scholar] [CrossRef]
- Dashraath, P.; Wong, J.L.J.; Lim, M.X.K.; Lim, L.M.; Li, S.; Biswas, A.; Choolani, M.; Mattar, C.; Su, L.L. Coronavirus disease 2019 (COVID-19) pandemic and pregnancy. Am. J. Obstet. Gynecol. 2020, 222, 521–531. [Google Scholar] [CrossRef]
- Terpos, E.; Stellas, D.; Rosati, M.; Sergentanis, T.N.; Hu, X.; Politou, M.; Pappa, V.; Ntanasis-Stathopoulos, I.; Karaliota, S.; Bear, J.; et al. SARS-CoV-2 antibody kinetics eight months from COVID-19 onset: Persistence of spike antibodies but loss of neutralizing antibodies in 24% of convalescent plasma donors. Eur. J. Intern. Med. 2021, 89, 87–96. [Google Scholar] [CrossRef]
- Fathizadeh, H.; Afshar, S.; Masoudi, M.R.; Gholizadeh, P.; Asgharzadeh, M.; Ganbarov, K.; Köse, Ş.; Yousefi, M.; Kafil, H.S. SARS-CoV-2 (COVID-19) vaccines structure, mechanisms and effectiveness: A review. Int. J. Biol. Macromol. 2021, 188, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.C.; Tortorici, M.A.; Snijder, J.; Xiong, X.; Bosch, B.-J.; Rey, F.A.; Veesler, D. Tectonic conformational changes of a coronavirus spike glycoprotein promote membrane fusion. Proc. Natl. Acad. Sci. USA 2017, 114, 11157–11162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sindona, C.; Schepici, G.; Contestabile, V.; Bramanti, P.; Mazzon, E. Nox2 activation in COVID-19: Possible implications for neurodegenerative diseases. Medicina 2021, 57, 604. [Google Scholar] [CrossRef]
- Ulhaq, Z.S.; Soraya, G.V. Interleukin-6 as a potential biomarker of COVID-19 progression. Med. Mal. Infect. 2020, 50, 382–383. [Google Scholar] [CrossRef]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef]
- Wieczfinska, J.; Kleniewska, P.; Pawliczak, R. Oxidative Stress-Related Mechanisms in SARS-CoV-2 Infections. Oxidative Med. Cell. Longev. 2022, 2022, 5589089. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.C.; Uhl, S.; Hoagland, D.; Møller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045.e9. [Google Scholar] [CrossRef]
- Finkel, T. Signal transduction by reactive oxygen species. J. Cell Biol. 2011, 194, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Bazhin, A.V.; Werner, J.; Karakhanova, S. Reactive Oxygen Species in the Immune System. Int. Rev. Immunol. 2013, 32, 249–270. [Google Scholar] [CrossRef]
- Schönrich, G.; Raftery, M.J.; Samstag, Y. Devilishly radical NETwork in COVID-19: Oxidative stress, neutrophil extracellular traps (NETs), and T cell suppression. Adv. Biol. Regul. 2020, 77, 100741. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Norouzi, P.; Aazami, H.; Moosavi-Movahedi, A.A. Review on oxidative stress relation on COVID-19: Biomolecular and bioanalytical approach. Int. J. Biol. Macromol. 2021, 189, 802–818. [Google Scholar] [CrossRef]
- To, E.E.; Broughton, B.R.S.; Hendricks, K.S.; Vlahos, R.; Selemidis, S. Influenza A virus and TLR7 activation potentiate NOX2 oxidase-dependent ROS production in macrophages. Free Radic. Res. 2014, 48, 940–947. [Google Scholar] [CrossRef]
- Fink, K.; Duval, A.; Martel, A.; Soucy-Faulkner, A.; Grandvaux, N. Dual Role of NOX2 in Respiratory Syncytial Virus- and Sendai Virus-Induced Activation of NF-κB in Airway Epithelial Cells. J. Immunol. 2008, 180, 6911–6922. [Google Scholar] [CrossRef]
- Kaul, P.; Biagioli, M.C.; Singh, I.; Turner, R.B. Rhinovirus-Induced Oxidative Stress and Interleukin-8 Elaboration Involves p47-phox but Is Independent of Attachment to Intercellular Adhesion Molecule-1 and Viral Replication. J. Infect. Dis. 2000, 181, 1885–1890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado-Roche, L.; Mesta, F. Oxidative Stress as Key Player in Severe Acute Respiratory Syndrome Coronavirus (SARS-CoV) Infection. Arch. Med. Res. 2020, 51, 384–387. [Google Scholar] [CrossRef]
- Sies, H.; Parnham, M.J. Potential therapeutic use of ebselen for COVID-19 and other respiratory viral infections. Free Radic. Biol. Med. 2020, 156, 107–112. [Google Scholar] [CrossRef]
- Barzegar, A.; Moosavi-Movahedi, A.A. Intracellular ROS Protection Efficiency and Free Radical-Scavenging Activity of Curcumin. PLoS ONE 2011, 6, e26012. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Gu, L.; Su, Y.; Wang, Q.; Zhao, Y.; Chen, X.; Deng, H.; Li, W.; Wang, G.; Li, K. Inhibition of curcumin on influenza A virus infection and influenzal pneumonia via oxidative stress, TLR2/4, p38/JNK MAPK and NF-κB pathways. Int. Immunopharmacol. 2018, 54, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Hashemi Gahruie, H.; Niakousari, M. Antioxidant, antimicrobial, cell viability and enzymatic inhibitory of antioxidant polymers as biological macromolecules. Int. J. Biol. Macromol. 2017, 104, 606–617. [Google Scholar] [CrossRef]
- El-Kenawi, A.; Ruffell, B. Inflammation, ROS, and Mutagenesis. Cancer Cell 2017, 32, 727–729. [Google Scholar] [CrossRef] [Green Version]
- Hosakote, Y.M.; Rayavara, K.; Hosakote, Y.M.; Rayavara, K. Respiratory Syncytial Virus-Induced Oxidative Stress in Lung Pathogenesis. In Oxidative Stress in Lung Diseases; Springer: Singapore, 2020; Volume 2, pp. 297–330. [Google Scholar] [CrossRef]
- Poprac, P.; Jomova, K.; Simunkova, M.; Kollar, V.; Rhodes, C.J.; Valko, M. Targeting Free Radicals in Oxidative Stress-Related Human Diseases. Trends Pharmacol. Sci. 2017, 38, 592–607. [Google Scholar] [CrossRef] [PubMed]
- Czarny, P.; Wigner, P.; Galecki, P.; Sliwinski, T. The interplay between inflammation, oxidative stress, DNA damage, DNA repair and mitochondrial dysfunction in depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 80, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Heine, A.; Juranek, S.; Brossart, P. Clinical and immunological effects of mRNA vaccines in malignant diseases. Mol. Cancer 2021, 20, 52. [Google Scholar] [CrossRef]
- Amanat, F.; Krammer, F. SARS-CoV-2 Vaccines: Status Report. Immunity 2020, 52, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Obermoser, G.; Presnell, S.; Domico, K.; Xu, H.; Wang, Y.; Anguiano, E.; Thompson-Snipes, L.A.; Ranganathan, R.; Zeitner, B.; Bjork, A.; et al. Systems Scale Interactive Exploration Reveals Quantitative and Qualitative Differences in Response to Influenza and Pneumococcal Vaccines. Immunity 2013, 38, 831–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Athale, S.; Banchereau, R.; Thompson-Snipes, L.; Wang, Y.; Palucka, K.; Pascual, V.; Banchereau, J. Influenza vaccines differentially regulate the interferon response in human dendritic cell subsets. Sci. Transl. Med. 2017, 9, eaaf9194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoshi, T.; Koyama, S.; Kobiyama, K.; Akira, S.; Ishii, K.J. Innate and adaptive immune responses to viral infection and vaccination. Curr. Opin. Virol. 2011, 1, 226–232. [Google Scholar] [CrossRef]
- Ntouros, P.A.; Vlachogiannis, N.I.; Pappa, M.; Nezos, A.; Mavragani, C.P.; Tektonidou, M.G.; Souliotis, V.L.; Sfikakis, P.P. Effective DNA damage response after acute but not chronic immune challenge: SARS-CoV-2 vaccine versus Systemic Lupus Erythematosus. Clin. Immunol. 2021, 229, 108765. [Google Scholar] [CrossRef]
- Li, S.C.; Yang, J.; Zeng, X.C.; Gao, Y.; Liang, X.; Duan, J.; Li, X.; Zhu, X.; Chen, Y.; Wang, X.; et al. Showcasing research from Tianjin Innovation and Promotion Plan Key Innovation Team of Immunoreactive Biomaterials Improved vaccine-induced immune responses via a ROS-triggered nanoparticle-based antigen delivery system. Nanoscale 2018, 10, 9489. [Google Scholar] [CrossRef]
- Trougakos, I.P.; Terpos, E.; Zirou, C.; Sklirou, A.D.; Apostolakou, F.; Gumeni, S.; Charitaki, I.; Papanagnou, E.D.; Bagratuni, T.; Liacos, C.I.; et al. Comparative kinetics of SARS-CoV-2 anti-spike protein RBD IgGs and neutralizing antibodies in convalescent and naïve recipients of the BNT162b2 mRNA vaccine versus COVID-19 patients. BMC Med. 2021, 19, 208. [Google Scholar] [CrossRef]
- Hoppe, A.; Güldal, N.S.; Boccaccini, A.R. A review of the biological response to ionic dissolution products from bioactive glasses and glass-ceramics. Biomaterials 2011, 32, 2757–2774. [Google Scholar] [CrossRef] [PubMed]
- Mouffak, S.; Shubbar, Q.; Saleh, E.; El-Awady, R. Recent advances in management of COVID-19: A review. Biomed. Pharmacother. 2021, 143, 112107. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Li, X.; Yang, G.; Fan, J.; Tang, Y.; Zhao, J.; Long, X.; Guo, S.; Zhao, Z.; Liu, Y.; et al. Serological immunochromatographic approach in diagnosis with SARS-CoV-2 infected COVID-19 patients. J. Infect. 2020, 81, e28–e32. [Google Scholar] [CrossRef]
- Tsamesidis, I.; Pantaleo, A.; Pekou, A.; Gusani, A.; Iliadis, S.; Makedou, K.; Manca, A.; Carruale, A.; Lymperaki, E.; Fozza, C. Correlation of oxidative stress biomarkers and hematological parameters in blood cancer patients from sardinia, Italy. Int. J. Hematol. Stem Cell Res. 2019, 13, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Tsamesidis, I.; Egwu, C.O.; Samara, D.; Vogiatzi, D.; Lettas, A.; Lymperaki, E. Effects of Greek Honey and Propolis on Oxidative Stress and Biochemical Parameters in Regular Blood Donors. J. Xenobiot. 2022, 12, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Lauring, A.S.; Tenforde, M.W.; Chappell, J.D.; Gaglani, M.; Ginde, A.A.; Mcneal, T.; Ghamande, S.; Douin, D.J.; Talbot, H.K.; Casey, J.D.; et al. Clinical severity of, and effectiveness of mRNA vaccines against, COVID-19 from omicron, delta, and alpha SARS-CoV-2 variants in the United States: Prospective observational study. BMJ Br. Med. J. 2022, 376, e069761. [Google Scholar] [CrossRef] [PubMed]
- Arbel, R.; Sergienko, R.; Friger, M.; Peretz, A.; Beckenstein, T.; Yaron, S.; Netzer, D.; Hammerman, A. Effectiveness of a second BNT162b2 booster vaccine against hospitalization and death from COVID-19 in adults aged over 60 years. Nat. Med. 2022, 28, 1486–1490. [Google Scholar] [CrossRef] [PubMed]
- Bergamaschi, C.; Terpos, E.; Rosati, M.; Angel, M.; Bear, J.; Stellas, D.; Karaliota, S.; Apostolakou, F.; Bagratuni, T.; Patseas, D.; et al. Systemic IL-15, IFN-γ, and IP-10/CXCL10 signature associated with effective immune response to SARS-CoV-2 in BNT162b2 mRNA vaccine recipients. Cell Rep. 2021, 36, 109504. [Google Scholar] [CrossRef] [PubMed]
- Saeed, U. Evaluation of SARS-CoV-2 spike antibody levels among Sputnik V rst dose vaccinated people in Pakistan: Formulation of national anti-COVID-19 mass vaccination strategy. Res. Sq. 2021, 1–9. [Google Scholar]
- Notarte, K.I.; Ver, A.T.; Velasco, J.V.; Pastrana, A.; Catahay, J.A.; Salvagno, G.L.; Yap, E.P.H.; Martinez-Sobrido, L.; Torrelles, J.B.; Lippi, G.; et al. Effects of age, sex, serostatus, and underlying comorbidities on humoral response post-SARS-CoV-2 Pfizer-BioNTech mRNA vaccination: A systematic review. Crit. Rev. Clin. Lab. Sci. 2022. [Google Scholar] [CrossRef]
- Terpos, E.; Trougakos, I.P.; Apostolakou, F.; Charitaki, I.; Sklirou, A.D.; Mavrianou, N.; Papanagnou, E.D.; Liacos, C.I.; Gumeni, S.; Rentziou, G.; et al. Age-dependent and gender-dependent antibody responses against SARS-CoV-2 in health workers and octogenarians after vaccination with the BNT162b2 mRNA vaccine. Am. J. Hematol. 2021, 96, E257–E259. [Google Scholar] [CrossRef] [PubMed]


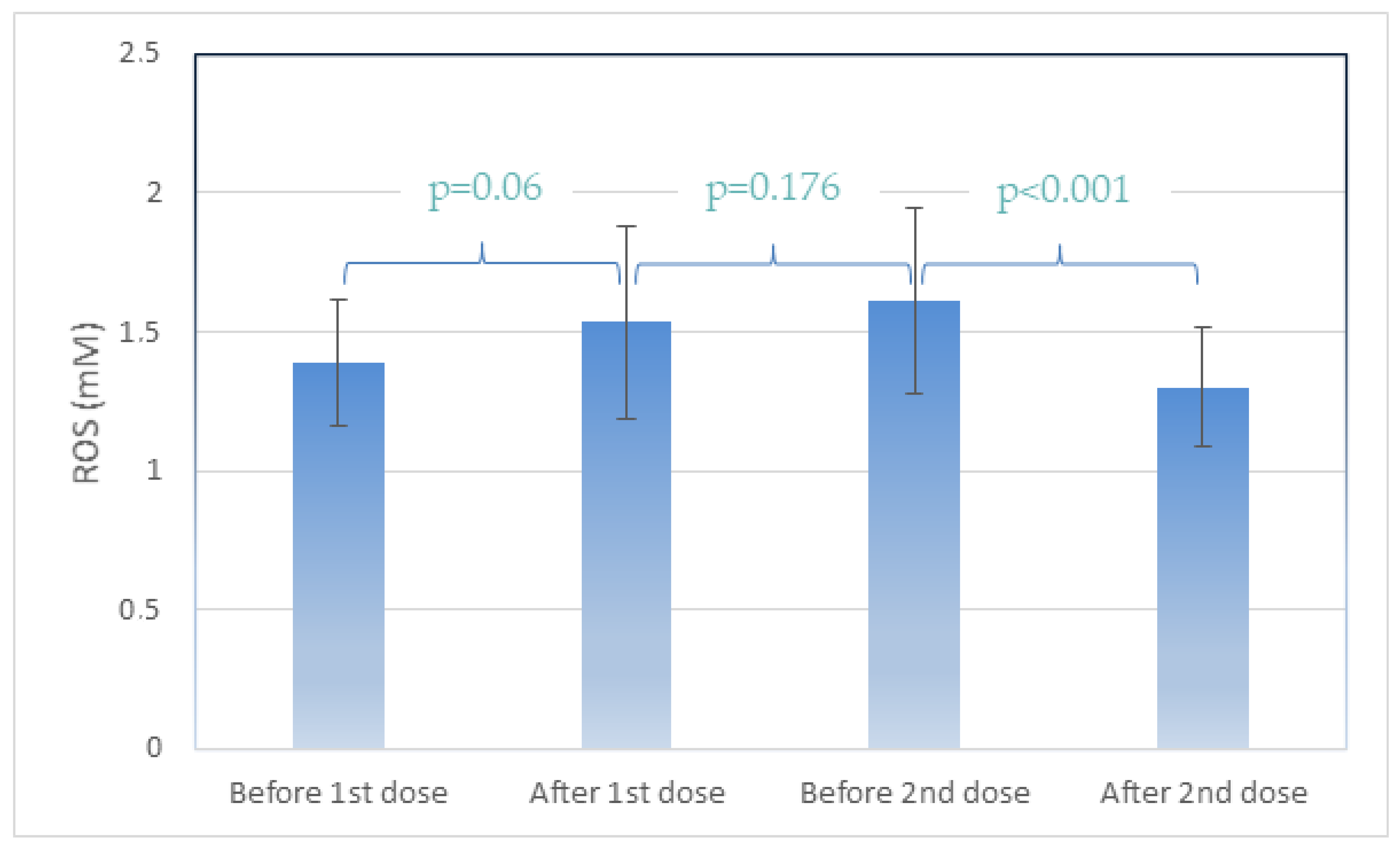{kind=link}
| 1st Study | 2nd Study | |||
|---|---|---|---|---|
| Vaccinated (2 Doses) | Infected and Unvaccinated Group A | Vaccinated (2 Doses) Group Β | Healthy Control Group C | |
| SEX | ||||
| Male | 9 | 10 | 79 | 5 |
| Female | 11 | 18 | 82 | 8 |
| Age | ||||
| 25–40 | 12 | 8 | 51 | 4 |
| 40–66 | 8 | 20 | 110 | 9 |
| Total participants | 20 | 28 | 161 | 13 |
| Before First Dose | After First Dose | Before Second Dose | After Second Dose | |||||
|---|---|---|---|---|---|---|---|---|
| Time Point 1 | Time Point 2 | Time Point 3 | Time Point 4 | |||||
| Participant | Reactive Oxygen Species (ROS) [mM] | Antibodies | Reactive Oxygen Species (ROS) [mM] | Antibodies | Reactive Oxygen Species (ROS) [mM] | Antibodies | Reactive Oxygen Species (ROS) [mM] | Antibodies |
| U/mL | U/mL | U/mL | U/mL | |||||
| ID1 | 1.634 | 40 | 2.146 | 50 | 2.127 | 5660 | 1.714 | 6330 |
| ID2 | 1.137 | 6.3 | 1.521 | 13 | 1.502 | 13470 | 1.236 | 13,250 |
| ID3 | 1.489 | 4.6 | 0.603 | 4.5 | 1.110 | 12780 | 1.285 | 13,250 |
| ID4 | 1.089 | 3.8 | 1.455 | 4.3 | 1.436 | 9500 | 1.547 | 9400 |
| ID5 | 1.588 | 7.5 | 1.714 | 8.5 | 2.081 | 6780 | 1.314 | 6800 |
| ID6 | 1.072 | 6.5 | 1.252 | 6.2 | 1.269 | 8200 | 0.847 | 8500 |
| ID7 | 1.318 | 2.8 | 1.787 | 3.2 | 1.870 | 5800 | 1.299 | 5300 |
| ID8 | 1.574 | 3.7 | 2.112 | 2.7 | 2.272 | 4500 | 1.53 | 5400 |
| ID9 | 1.354 | 37 | 1.471 | 19 | 1.498 | 7150 | 1.285 | 7200 |
| ID10 | 1.235 | 15 | 1.293 | 14.5 | 1.356 | 12000 | 1.225 | 11,980 |
| ID11 | 1.872 | 1.8 | 1.890 | 2.7 | 1.990 | 5400 | 1.446 | 5600 |
| ID12 | 1.428 | 2.8 | 1.618 | 2.9 | 1.656 | 6800 | 1.300 | 6700 |
| ID13 | 1.307 | 0.6 | 1.291 | 0.6 | 1.302 | 12700 | 1.056 | 11,900 |
| ID14 | 1.137 | 0.7 | 1.317 | 0.9 | 1.373 | 11150 | 1.147 | 11,650 |
| ID15 | 1.737 | 16 | 1.861 | 10.3 | 1.821 | 6750 | 1.490 | 6700 |
| ID16 | 1.291 | 2.8 | 1.360 | 14 | 1.389 | 8500 | 1.228 | 8400 |
| ID17 | 1.152 | 6.3 | 1.302 | 10.5 | 1.323 | 10.900 | 1.005 | 11,110 |
| ID18 | 1.433 | 8.5 | 1.656 | 9.5 | 1.589 | 7200 | 1.384 | 7300 |
| ID19 | 1.325 | 9.5 | 1.426 | 9.4 | 1.375 | 11.500 | 1.080 | 11,150 |
| ID20 | 1.574 | 28 | 1.590 | 17.8 | 1.887 | 7200 | 1.618 | 7150 |
| Average | 1.387 | 23.7 | 1.533 | 24 | 1.611 | 8697 | 1.302 | 8735 |
| SD | 0.224 | 0.346 | 0.331 | 0.216 | ||||
| ROS (mM) | Antibodies (U/mL) | r-Value | p-Value | ||
|---|---|---|---|---|---|
| Mean | |||||
| Group A (n = 28) | 1.213 | 6584 | −0.55 | 0.001 | |
| Male | 10 | 1.373 | 10,642 | −0.88 | 0.001 |
| Female | 18 | 1.434 | 5076 | −0.23 | 0.001 |
| Group B (n = 161) | 1.243 | 12,179 | 0.01 | <0.001 | |
| Male | 79 | 1.419 | 9668 | 0.1 | <0.001 |
| Female | 82 | 1.194 | 13,178 | −0.00044 | 0.01 |
| Group C (n = 13) | 0.885 | 6.26 | −0.24 | <0.0001 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lymperaki, E.; Kazeli, K.; Tsamesidis, I.; Nikza, P.; Poimenidou, I.; Vagdatli, E. A Preliminary Study about the Role of Reactive Oxygen Species and Inflammatory Process after COVID-19 Vaccination and COVID-19 Disease. Clin. Pract. 2022, 12, 599-608. https://doi.org/10.3390/clinpract12040063
Lymperaki E, Kazeli K, Tsamesidis I, Nikza P, Poimenidou I, Vagdatli E. A Preliminary Study about the Role of Reactive Oxygen Species and Inflammatory Process after COVID-19 Vaccination and COVID-19 Disease. Clinics and Practice. 2022; 12(4):599-608. https://doi.org/10.3390/clinpract12040063
Chicago/Turabian StyleLymperaki, Evgenia, Konstantina Kazeli, Ioannis Tsamesidis, Polykseni Nikza, Irini Poimenidou, and Eleni Vagdatli. 2022. "A Preliminary Study about the Role of Reactive Oxygen Species and Inflammatory Process after COVID-19 Vaccination and COVID-19 Disease" Clinics and Practice 12, no. 4: 599-608. https://doi.org/10.3390/clinpract12040063
APA StyleLymperaki, E., Kazeli, K., Tsamesidis, I., Nikza, P., Poimenidou, I., & Vagdatli, E. (2022). A Preliminary Study about the Role of Reactive Oxygen Species and Inflammatory Process after COVID-19 Vaccination and COVID-19 Disease. Clinics and Practice, 12(4), 599-608. https://doi.org/10.3390/clinpract12040063