Isolation and Chemical Characterization of a Toxin Isolated from the Venom of the Sea Snake, Hydrophis torquatus aagardi

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation procedure
2.2. Toxicity test
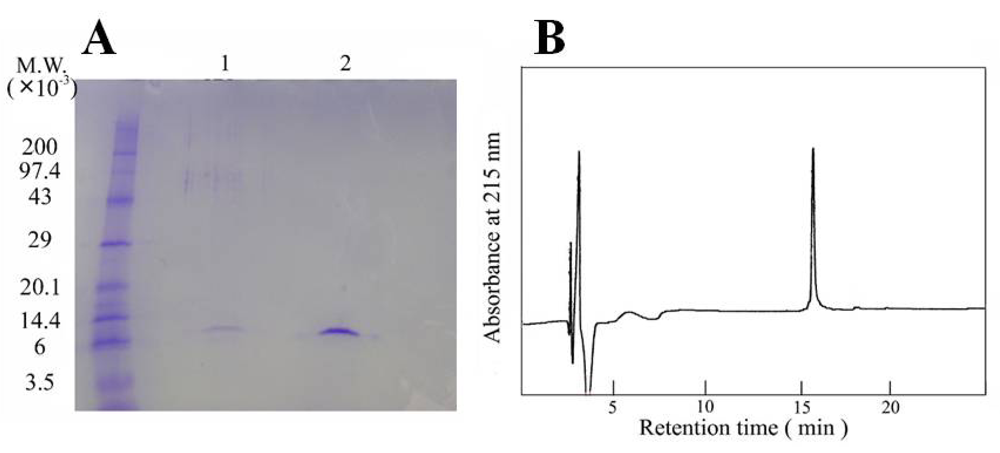
2.3. Homogeneity
2.4. Enzymatic cleavage
2.5. Primary structure analysis
2.6. Molecular mass
2.7. Isoelectric focusing/polyacrylamide gel electrophoresis
2.8. Other activities
3. Results
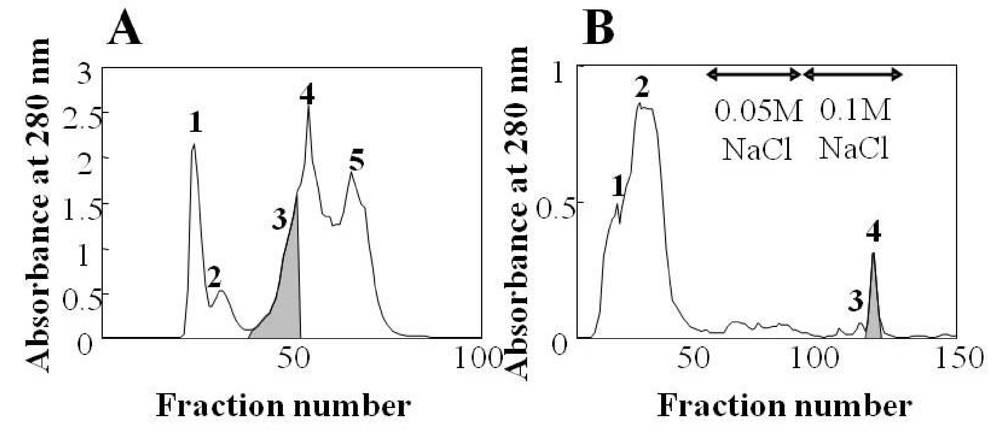
3.1. Isolation and purification
3.2. Toxicity
3.3. Chemical properties
3.4. Reduction of disulfide bonds
3.5. Enzymatic and biological assays
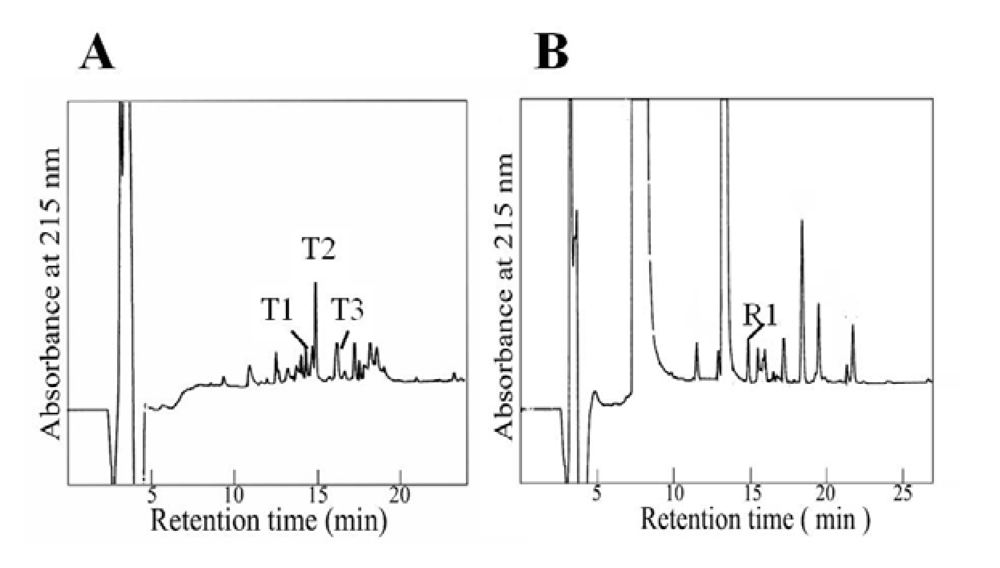
3.6. Fragments
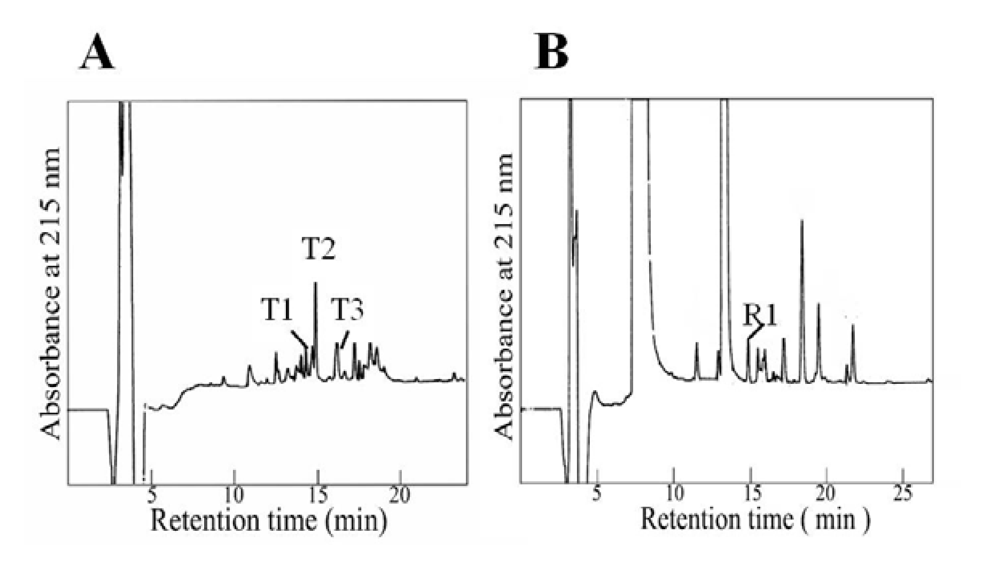
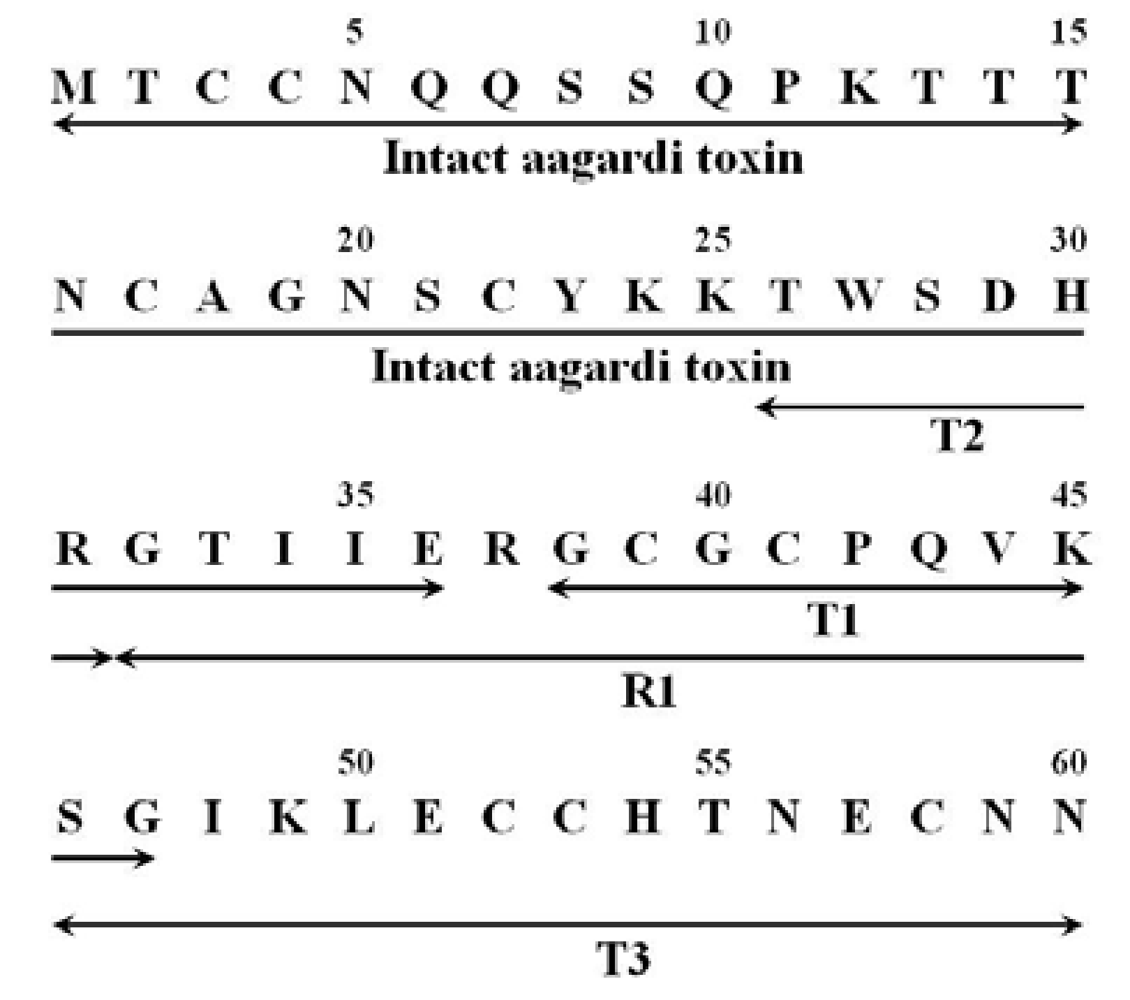
4. Discussion
4.1. Stability of sea snake toxins
4.2. Non-enzymatic nature of sea snake toxins
4.3. Comparison of primary structure within Hydrophiinae toxins
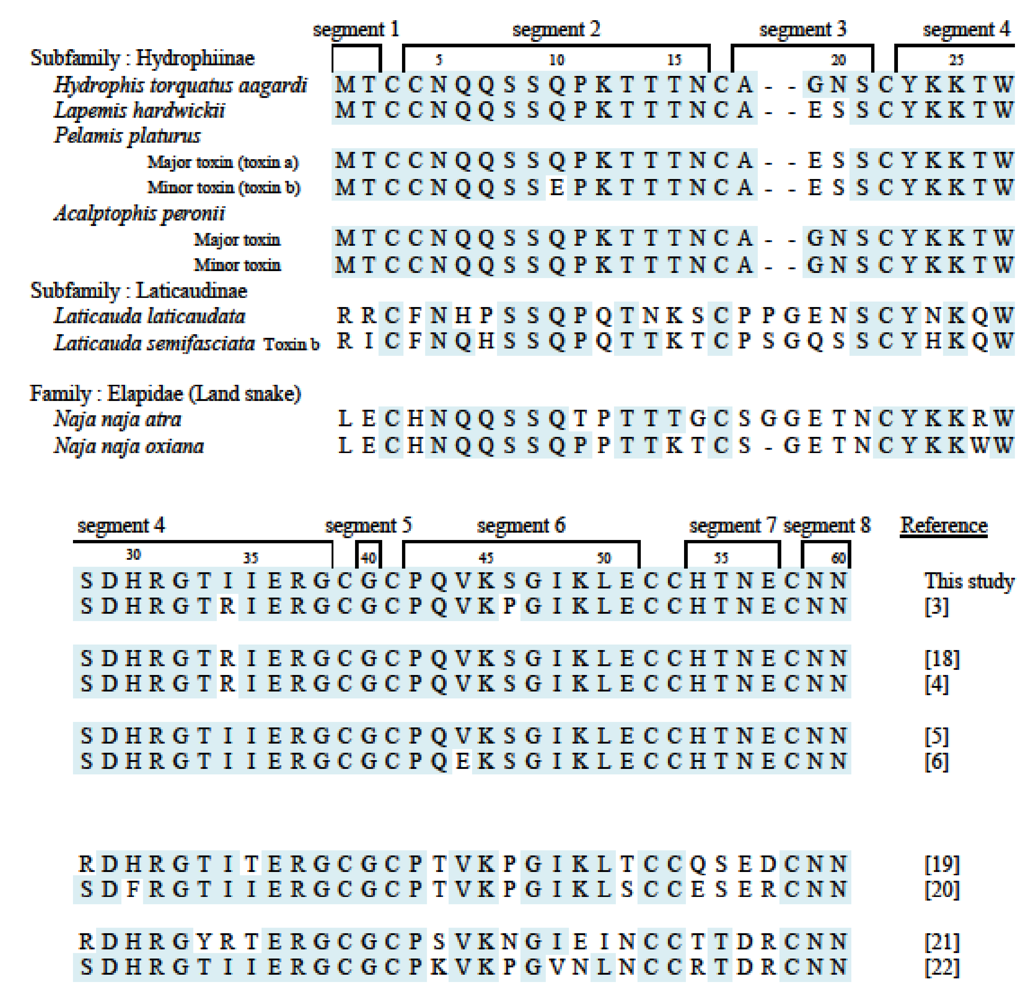
- 1. Position 10: Most toxins show Q (glutamine), with only Pelamis platurus minor toxin having E (glutamic acid).
- 2. By comparing the amino acid residues it is apparent that the amino acid residue at 19 is E for Pelamis minor toxin and G for aagardi toxin (Figure 6). At position 34, R for Pelamis major toxin becomes I for aagardi toxin. It is premature to conclude definitely, however the differences at 19 and 34 may be correlated with toxicity difference.
- 3. As compared to the venoms of land snakes, enzymatic activity of sea snake venoms has rarely been studied. Therefore, we really don’t know the enzyme activity until we actually test. Presynaptic toxins are known to have phospholipase A2 activity; it is for this reason that it’s worthwhile to investigate such activities.
- 4. Position 44: Most toxins have V (valine), and only one toxin shows E.
- 5. Position 46: Most are V; only Lapemis hardwickii shows P (proline).
4.4. Comparison of Hydrophiinae and Laticaudinae toxins
4.5. Comparison with land snake (Elapidae) toxins
References
- Tu, A.T. Neurotoxins of animal venoms: Snakes. Ann. Rev. Biochem. 1973, 42, 235. [Google Scholar] [CrossRef]
- Tu, A.T. Venoms. Chemistry and Molecular Biology; John Wiley and Sons: New York, NY, USA, 1977; pp. 151–177. [Google Scholar]
- Fox, J.W.; Elzinga, M.; Tu, A.T. Amino acid sequence of a snake neurotoxin from the venom of Lapemis hardwickii and the detection of a sulfhydryl group by laser Raman spectroscopy. FEBS Lett. 1977, 80, 210–240. [Google Scholar]
- Mori, N.; Ishizaki, H.; Tu, A.T. Isolation and characterization of Pelamis platurus (yellow-bellied sea snake) postsynaptic neurotoxin. J. Pharm. Pharmacol. 1989, 41, 331–334. [Google Scholar] [PubMed]
- Mori, N.; Tu, A.T. Isolation and primary structure of the major toxin from sea snake, Acalyptophis peronii, venom. Arch. Biochem. Biophys. 1988, 260, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Mori, N.; Tu, A.T. Amino-acid sequence of the minor neurotoxin from Acalyptophis peronii venom. Biol. Chem. Hoppe. Seyler 1988, 369, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Litchfield, J.T.; Wilcoxon, F. A simplified method of evaluating dose-effect experiments. J. Pharmacol. Exp. Therap. 1949, 96, 99–113. [Google Scholar]
- Weber, K.; Osborn, M. The reliability of molecular weight determinations by sulfate-polyacrylamide gel electrophoresis. J. Biol. Chem. 1969, 244, 4406–4412. [Google Scholar] [PubMed]
- Hendon, R.A.; Fraenkel-Conrat, H. Biological roles of two components of crotoxin. Proc. Natl. Acad. Sci. USA 1971, 68, 1560–1563. [Google Scholar] [CrossRef]
- Michl, H.; Molzer, H. The occurrence of L-leucyl-b-naphthylamide (LNA) splitting enzymes in some amphibia and reptile venoms. Toxicon 1965, 2, 281–282. [Google Scholar] [CrossRef]
- Murata, Y.; Satake, M.; Suzuki, T. Studies on snake venom. XII. Distribution of proteinase activities among Japanese and Formosan snake venoms. J. Biochem. Tokyo 1963, 53, 431–437. [Google Scholar]
- Roberts, P.S. Measurement of the rate of plasmin action on synthetic substrates. J. Biochem. 1958, 232, 285–291. [Google Scholar]
- Chenxi, O.; Che-Ming, T. Fiblinogenolytic enzymes of Trimeresurus mucrosquamatus venom. Biochim. Biophys. Acta 1976, 420, 298–308. [Google Scholar] [PubMed]
- Okumura, Y.; Ogawa, K.; Uchiya, K.; Komori, Y.; Nonogaki, T.; Nikai, T. Biological properties of elastase inhibitor, AFLEI from Aspergillus flavus. Jpn. J. Med. Mycol. 2008, 49, 87–93. [Google Scholar] [CrossRef]
- Bjarnason, J.B.; Tu, A.T. Hemorrhagic toxins from western diamondback rattlesnake (Crotalus atrox) venom: isolation and characterization of five toxins and the role of zinc in hemorrhagic toxin e. Biochemistry 1978, 17, 3395–3404. [Google Scholar] [PubMed]
- Tamiya, N.; Maeda, N.; Cogger, H.G. Neurotoxins from the venom of the sea snakes Hydrophis ornatus and Hydrophis lapemoides. Biochem. J. 1983, 213, 31–38. [Google Scholar] [PubMed]
- Tu, A.T. Sea snakes and their venoms. In Handbook of Natural of Toxins: Marine Toxins and venoms; Marcel Dekker: New York, NY, USA, 1988; Volume 3, pp. 379–444. [Google Scholar]
- Wang, C.L.; Liu, C.S.; Hung, Y.O.; Blackwell, R.Q. Amino acid sequence of pelamitoxin a, the main neurotoxin of the sea snake, Pelamis platurus. Toxicon 1976, 14, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Maeda, N.; Tamiya, N. The primary structure of the toxin Laticauda semifasciata III, a weak and reversibly acting neurotoxin from the venom of a sea snake, Laticauda semifasciata. Biochem. J. 1974, 141, 389–400. [Google Scholar] [PubMed]
- Tsernoglou, P.; Petsko, G.A.; Tu, A.T. Protein sequencing by computer graphics. Biochim. Biophys. Acta 1977, 491, 605–608. [Google Scholar] [PubMed]
- Yang, C.C.; Yang, H.J.; Huang, J.S. The amino acid sequence of cobrotoxin. Biochim. Biophys. Acta 1969, 188, 65–77. [Google Scholar] [PubMed]
- Armberg, H.; Eaker, D.; Frykuand, L.; Karlsson, E. Amino acid sequence of oxiana a, The main neurotoxin of the venom of Naja naja oxiana. Biochim. Biophys. Acta 1974, 359, 222–232. [Google Scholar] [PubMed]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nagamizu, M.; Komori, Y.; Uchiya, K.-i.; Nikai, T.; Tu, A.T. Isolation and Chemical Characterization of a Toxin Isolated from the Venom of the Sea Snake, Hydrophis torquatus aagardi. Toxins 2009, 1, 162-172. https://doi.org/10.3390/toxins1020162
Nagamizu M, Komori Y, Uchiya K-i, Nikai T, Tu AT. Isolation and Chemical Characterization of a Toxin Isolated from the Venom of the Sea Snake, Hydrophis torquatus aagardi. Toxins. 2009; 1(2):162-172. https://doi.org/10.3390/toxins1020162
Chicago/Turabian StyleNagamizu, Masaya, Yumiko Komori, Kei-ichi Uchiya, Toshiaki Nikai, and Anthony T. Tu. 2009. "Isolation and Chemical Characterization of a Toxin Isolated from the Venom of the Sea Snake, Hydrophis torquatus aagardi" Toxins 1, no. 2: 162-172. https://doi.org/10.3390/toxins1020162
APA StyleNagamizu, M., Komori, Y., Uchiya, K.-i., Nikai, T., & Tu, A. T. (2009). Isolation and Chemical Characterization of a Toxin Isolated from the Venom of the Sea Snake, Hydrophis torquatus aagardi. Toxins, 1(2), 162-172. https://doi.org/10.3390/toxins1020162