β-N-Methylamino-L-alanine (BMAA) Toxicity Is Gender and Exposure-Age Dependent in Rats

Abstract
:1. Introduction
2. Results and Discussion
2.1. General Findings
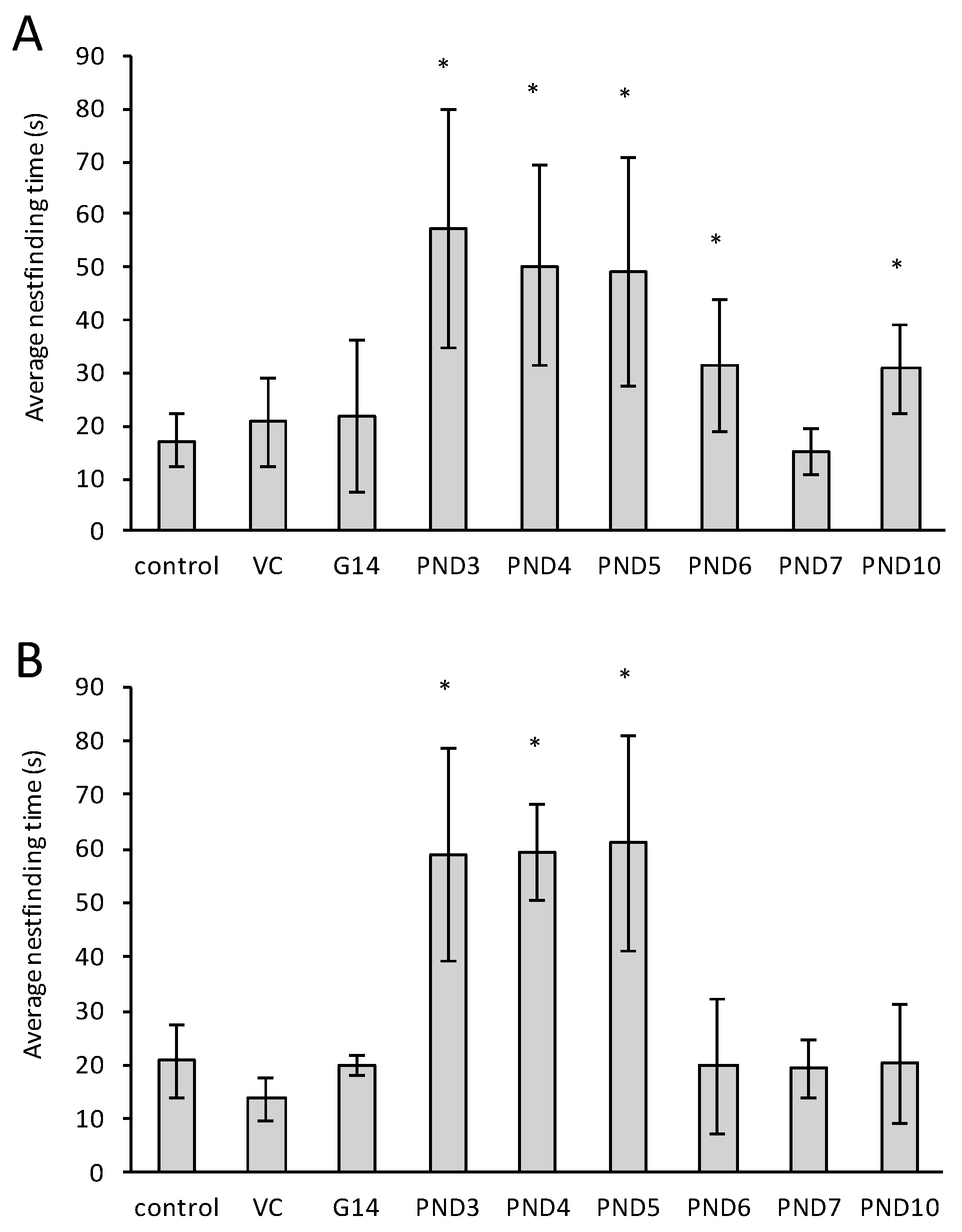
2.2. The Nest Finding Test
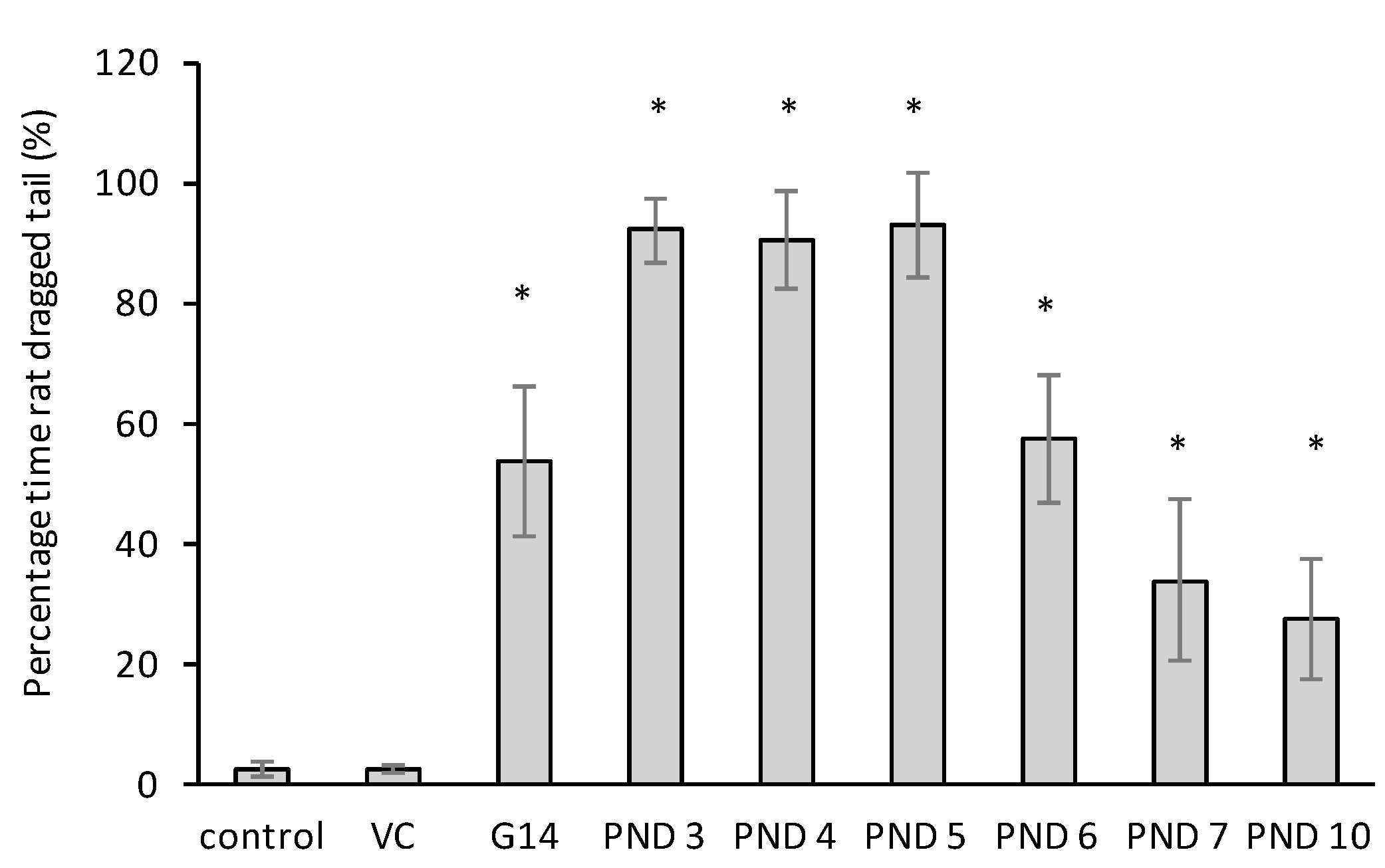
2.3. The Modified Open Field Test
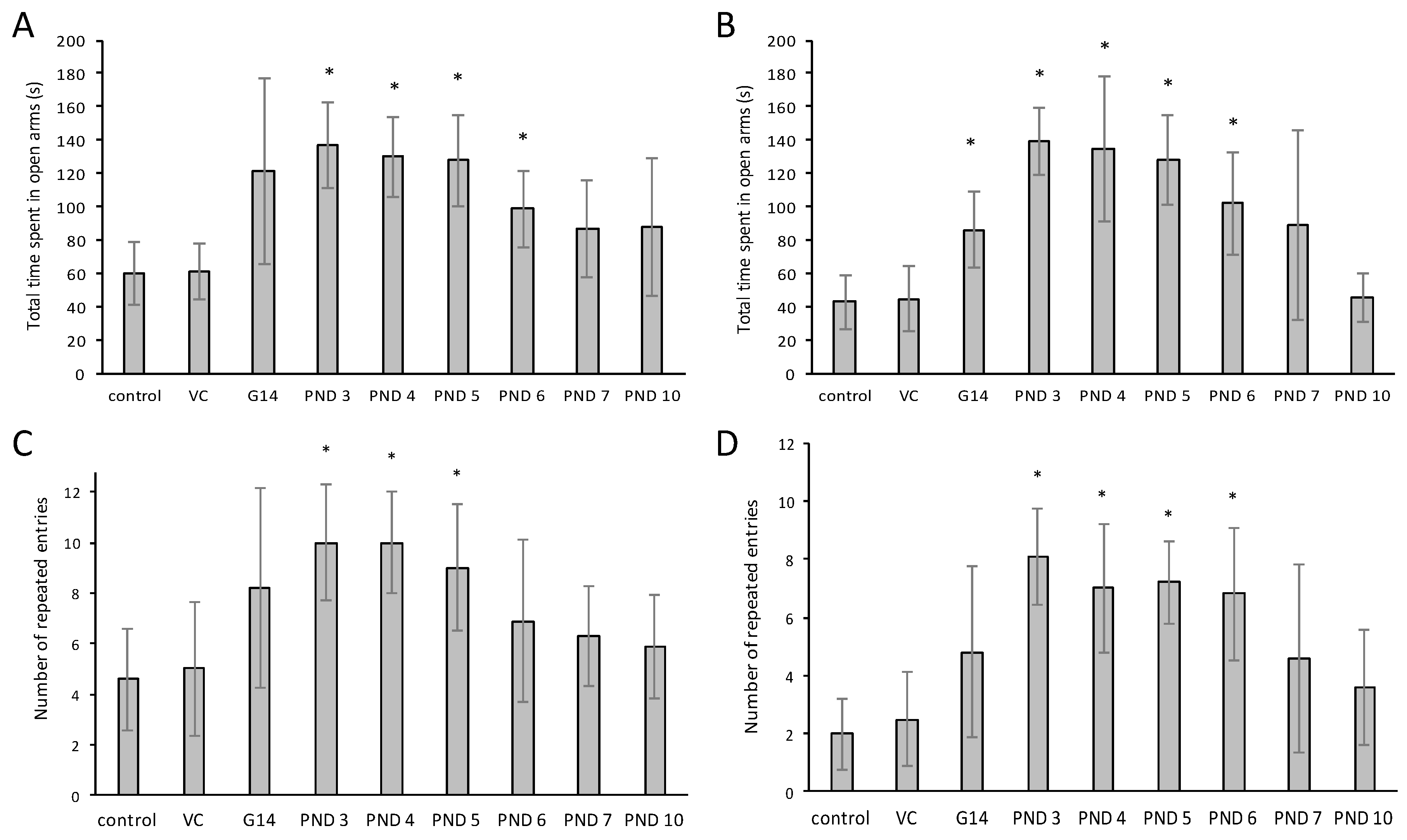
2.4. The Elevated Plus Maze
2.5. The Inclined Plane Test
2.6. Audiogenic Response
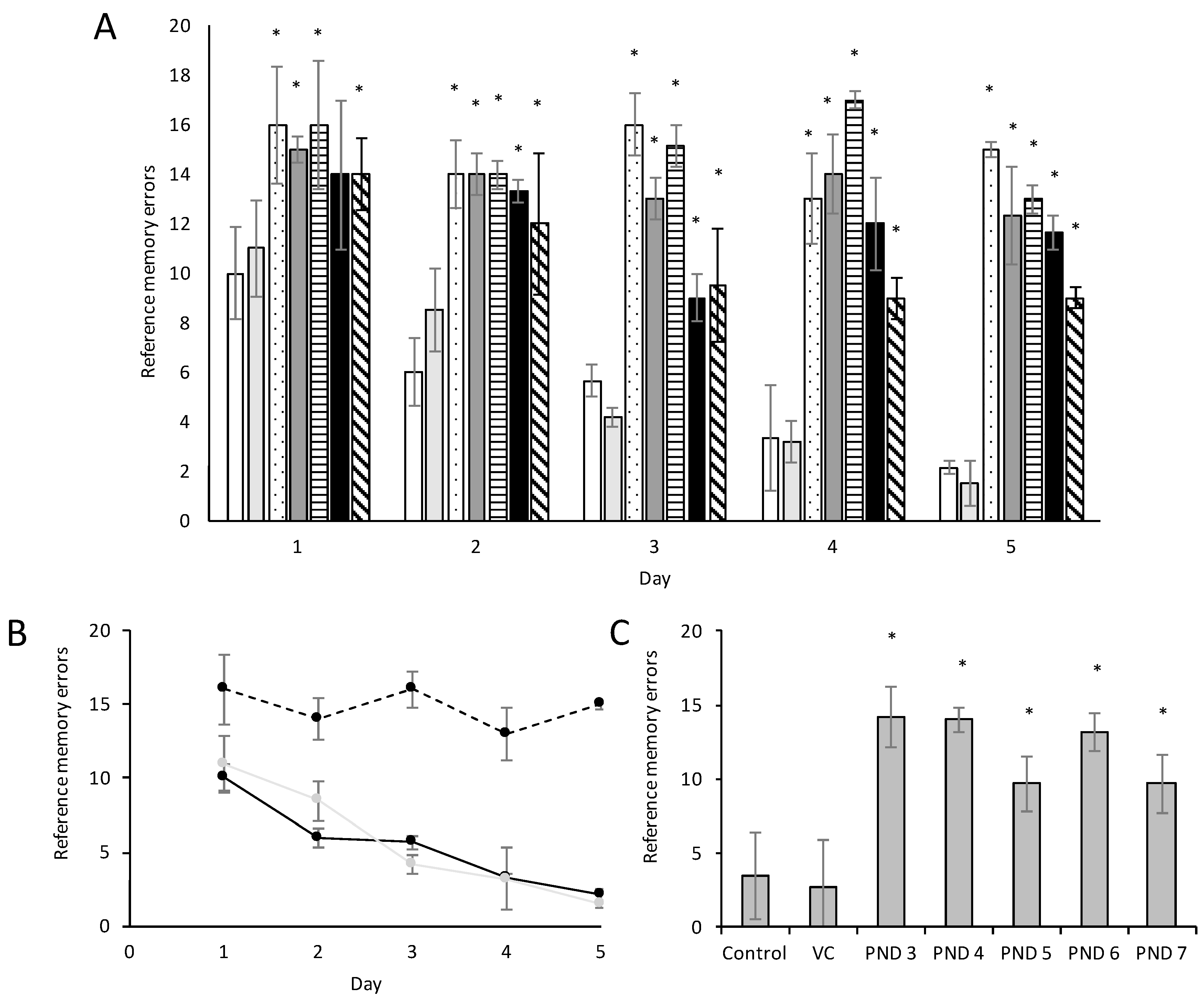
2.7. The Radial Arm
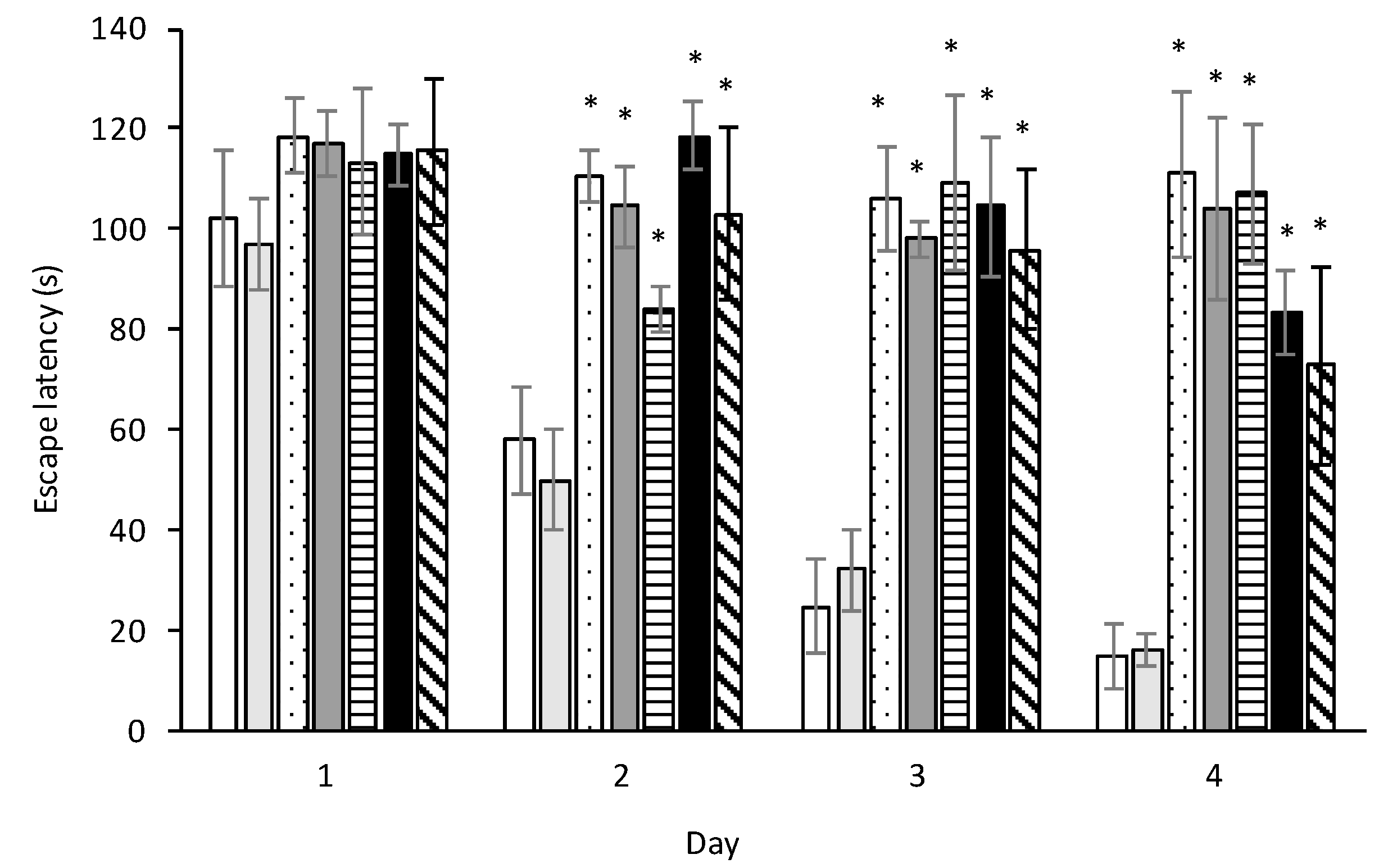
2.8. Ihe Morris Water Maze
3. Conclusions
4. Materials and Methods
4.1. Chemicals
4.2. Animal Maintenance
4.3. Exposure
4.4. Behavioural Analysis
4.4.1. The Nestfinding Test on Postnatal Day 10
4.4.2. The Modified Open Field Test on Postnatal Day 20
4.4.3. The Elevated Plus Maze Test on Postnatal Day 30
4.4.4. The Inclined Plane Test on Postnatal Day 37
4.4.5. The Audiogenic Freezing Response on Postnatal Day 55
4.4.6. The Radial Arm Maze Test from Postnatal Day 90
4.4.7. Morris Water Maze Test from Postnatal Day 108
4.5. Behavioural Recordings
4.6. Statistical Analysis
5. Patents
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dietrich, D.R.; Fischer, A.; Michel, C.; Hoeger, S.J. Toxin mixture in cyanobacterial bloom—A critical comparison of reality with current procedures employed in human health risk assessment. Cyanobact. Harmful Algal Blooms 2008, 619, 885–912. [Google Scholar]
- Brand, L.E. Human exposure to cyanobacteria and BMAA. Amyotroph. Lateral Scler. 2009, 10, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Brand, L.E.; Pablo, J.; Compton, A.; Hammerschlag, N.; Mash, D.C. Cyanobacterial blooms and the occurrence of the neurotoxin, β-N-methylamino-l-alanine (BMAA), in South Florida aquatic food webs. Harmful Algae 2010, 9, 620–635. [Google Scholar] [CrossRef] [PubMed]
- Jonasson, S.; Eriksson, J.; Berntzon, L.; Spáčil, Z.; Ilag, L.L.; Ronnevi, L.-O.; Rasmussen, U.; Bergman, B. Transfer of a cyanobacterial neurotoxin within a temperate aquatic ecosystem suggests pathways for human exposure. Proc. Natl. Acad. Sci. USA 2010, 107, 9252–9257. [Google Scholar] [CrossRef] [PubMed]
- Mondo, L.; Hammerschlag, N.; Basile, M.; Pablo, J.; Banack, S.A.; Mash, D.C. Cyanobacterial neurotoxin β-N-methylamino-l-alanine (BMAA) in shark fins. Mar. Drugs 2012, 10, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Downing, S.; Contardo-Jara, V.; Pflugmacher, S.; Downing, T.G. The fate of the cyanobacterial toxin β-N-methylamino-l-alanine in freshwater mussels. Ecotoxicol. Environ. Saf. 2014, 101, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Contardo-Jara, V.; Schwanemann, T.; Pflugmacher, S. Uptake of a cyanotoxin β-N-methylamino-l-alanine (BMAA), by wheat (Triticum aestivum). Ecotoxicol. Environ. Saf. 2014, 104, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Al-Sammak, M.A.; Hoagland, K.D.; Cassada, D.; Snow, D.D. Co-occurrence of the cyanotoxins BMAA, DABA and Anatoxin-A in Nebraska Reservoirs, Fish and Aquatic Plants. Toxins 2014, 6, 488–508. [Google Scholar] [CrossRef] [PubMed]
- Mondo, K.; Glover, B.W.; Murch, S.J.; Liu, G.; Cai, Y.; Davis, D.A.; Mash, D.C. Environmental neurotoxins β-N-methylamino-l-alanine (BMAA) and mercury in shark cartilage dietary supplements. Food Chem. Toxicol. 2014, 70, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Kiselova, N.; Rosen, J.; Ilag, L.L. Quantification of neurotoxin BMAA (β-N-methylamino-l-alanine) in seafood from Swedish markets. Sci. Rep. 2014, 4, 6931. [Google Scholar] [CrossRef] [PubMed]
- Lage, S.; Annadotter, H.; Rasmussen, U.; Rydberg, S. Biotransfer of β-N-methylamino-l-alanine (BMAA) in a Eutrophicated Freshwater Lake. Mar. Drugs 2015, 13, 1185–1201. [Google Scholar] [CrossRef] [PubMed]
- Reveillon, D.; Abadie, E.; Sechet, V.; Masseret, E.; Hess, P.; Amzil, Z. β-N-methylamino-l-alanine (BMAA) and isomers: Distribution on different food web compartments of Thau lagoon, French Mediterranean Sea. Mar. Environ. Res. 2015, 110, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Reveillon, D.; Sechet, V.; Hess, P.; Amzil, Z. Systemic detection of BMAA (β-N-methylamino-l-alanine) and DAB (2,4-diaminobutyric acid) in mollusks collected in shellfish production areas along the French coasts. Toxicon 2016, 110, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.A.; Sacks, O.W. Cycad neurotoxins, consumption of flying foxes, and ALS-PDC disease in Guam. Neurol 2002, 58, 956–959. [Google Scholar] [CrossRef]
- Murch, S.J.; Cox, P.A.; Banack, S.A. A mechanism for slow release of biomagnified cyanobacterial toxins and neurodegenerative disease in Guam. Proc. Natl. Acad. Sci. USA 2004, 101, 12228–12231. [Google Scholar] [CrossRef] [PubMed]
- Seawright, A.A.; Brown, A.W.; Nolan, C.C.; Cavanagh, J.B. Selective degeneration of cerebellar cortical neurons caused by cycad neurotoxin l-b-methylaminoalanine (BMAA), in rats. Neuropathol. Appl. Neurobiol. 1990, 16, 153–169. [Google Scholar] [CrossRef] [PubMed]
- Vega, A.; Bell, E.A.; Nunn, P.B. The preparation of l- and d-α-amino-bmethylaminopropionicacids and the identification of the compound isolated from Cycas circinalis as the l-isomer. Phytochemistry 1968, 7, 1885–1887. [Google Scholar] [CrossRef]
- Polsky, F.I.; Nunn, P.B.; Bell, E.A. Distribution and toxicity of amino-b-methylaminopropionic acid. Fed. Proc. 1972, 31, 1473–1475. [Google Scholar] [PubMed]
- Karlsson, O.; Roman, E.; Brittebo, E.B. Long-term cognitive impairments in adult rats treated neonatally with beta-N-Methylamino-l-Alanine. Toxicol. Sci. 2009, 112, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, O.; Lindquist, N.G.; Brittebo, E.B.; Roman, E. Selective Brain Uptake and Behavioural Effects of the Cyanobacterial Toxin BMAA (b-N-Methylamino-l-alanine) following Neonatal Administration to Rodents. Toxicol. Sci. 2009, 109, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Al-Sammak, M.A.; Rogers, D.G.; Hoagland, K.D. Acute B-N-Methylamino-l-alanine Toxicity in a mouse model. J. Toxicol. 2015, 2015, 739746. [Google Scholar] [CrossRef] [PubMed]
- Rakonczay, Z.; Matsuoka, Y.; Giacobini, E. Effects of l-β-N-methylamino-l-alanine (l-BMAA) on the cortical cholinergic and glutamatergic systems of the rat. J. Neurosci. Res. 1991, 29, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-C.; Chiu, S.-J.; Kao, K.-P. Beta-N-methylamino-l-alanine (l-BMAA) decreases brain glutamate receptor number and induces behavioral changes in rats. Chin. J. Physiol. 1993, 36, 79–84. [Google Scholar] [PubMed]
- Perry, T.L.; Bergeron, C.; Biro, A.J.; Hansen, S. Chronic oral administration of b-N-methylamino-l-alanine is not neurotoxic to mice. J. Neurol. Sci. 1989, 94, 173–180. [Google Scholar] [CrossRef]
- Duncan, M.W.; Villacreses, N.E.; Pearson, P.G.; Wyatt, L.; Rapoport, S.I.; Kopin, I.J.; Markey, S.P.; Smith, Q.R. 2-Amino-3-(methylamino)-propanoic acid (BMAA) pharmacokinetics and blood–brain barrier permeability in the rat. J. Pharmacol. Exp. Ther. 1991, 258, 27–35. [Google Scholar] [PubMed]
- Cruz-Aguado, R.; Winkler, D.; Shaw, C.A. Lack of behavioral and neuropathological effects of dietary β-methylamino-l-alanine (BMAA) in mice. Pharmacol. Biochem. Behav. 2006, 84, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.L.; Downing, S.; Downing, T.G. The evaluation of BMAA inhalation as a potential exposure route using a rat model. Neurotox. Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Smith, Q.R.; Nagura, H.; Takada, Y.; Duncan, M.W. Facilitated transport of the neurotoxin, beta-N-methylamino-l-alanine, across the blood–brain barrier. J. Neurochem. 1992, 8, 1330–1337. [Google Scholar] [CrossRef]
- Banos, G.; Daniel, P.M.; Pratt, O.E. The effect of age upon the entry of some amino acids into the brain, and their incorporation into cerebral protein. Dev. Med. Child Neurol. 1978, 20, 335–346. [Google Scholar] [CrossRef] [PubMed]
- O’Tuama, L.A.; Phillips, P.C.; Smith, Q.R.; Uno, Y.; Dannals, R.F.; Wilson, A.A.; Ravert, H.T.; Loats, S.; Loats, H.A.; Wagner, H.N., Jr. l-methionine uptake by human cerebral cortex: Maturation from infancy to old age. J. Nucl. Med. 1991, 32, 16–22. [Google Scholar] [PubMed]
- Dawson, R., Jr.; Marschall, E.G.; Chan, K.C.; Millard, W.J.; Eppler, B.; Patterson, T.A. Neurochemical and neurobehavioral effects of neonatal administration of b-methylamino-l-alanine and 3,3′iminodipropionitrile. Neurotoxicol. Teratol. 1998, 20, 181–192. [Google Scholar] [CrossRef]
- Karlsson, O.; Roman, E.; Berg, A.L.; Brittebo, E.B. Early hippocampal cell death, and late learning and memory deficits in rats exposed to the environmental toxin BMAA (β-N-methylamino-l-alanine) during the neonatal period. Behav. Brain Res. 2011, 219, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, O.; Berg, A.-L.; Lindstrom, A.-K.; Arnerup, G.; Roman, E.; Bergquist, J.; Hanrieder, J.; Lindquist, N.G.; Brittebo, E.B.; Andersson, M. Neonatal exposure to the cyanobacterial toxin BMAA induces changes in protein expression, and neurodegeneration in adult hippocampus. Toxicol. Sci. 2012, 130, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Dobbing, J.; Sands, J. Comparative aspects of the brain growth spurt. Early Hum. Dev. 1979, 3, 79–83. [Google Scholar] [CrossRef]
- Gottlieb, A.; Keydar, I.; Epstein, H.T. Rodent brain growth spurts: An analytical review. Biol. Neonate 1977, 32, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Baloch, S.; Verma, R.; Huang, H.; Khurd, P.; Clark, S. Quantification of brain maturation and growth patterns in C57Bl/6 mice via computational neuroanatomy of diffusion tensor images. Cereb. Cortex 2009, 19, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Bockhorst, K.H.; Narayana, P.A.; Liu, R.; Ahobila-Vijjula, P.; Ramu, J. Early postnatal development of rat brain: In vivo diffusion tensor imaging. J. Neurosci. Res. 2008, 86, 1520–1528. [Google Scholar] [CrossRef] [PubMed]
- Semple, B.D.; Blomgren, K.; Gimlin, K.; Ferriero, D.M.; Noble-Haeusslein, L. Brain development in rodents and humans: Identifying benchmarks of maturation and vulnerability to injury across species. Prog. Neurobiol. 2013, 106–107, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Garruto, R.M.; Gajdusek, D.C.; Chen, K.M. Amyotrophic lateral sclerosis and Parkinsonism dementia among Filipino migrants to Guam. Ann. Neurol. 1981, 10, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Sabel, C.E.; Boyle, P.J.; Löytönen, M.; Gatrell, A.C.; Jokelainen, M.; Flowerdew, R.; Maasilta, P. Spatial clustering of amyotrophic lateral sclerosis in Finland at place of birth and place of death. Am. J. Epidemiol. 2003, 157, 898–905. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.M.; Campos, F.L.; Coimbra, B.; Pêgo, J.M.; Rodrigues, C.; Lima, R.; Rodrigues, A.J.; Sousa, N.; Salgado, A.J. Behavioural characterization of the 6-hydroxidopamine model of Parkinson’s disease and pharmacological rescuing of non-motor deficits. Mol. Neurodegener. 2013, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meredith, G.E.; Kang, U.J. Behavioral models of Parkinson’s disease in rodents: A new look at an old problem. Mov. Disord. 2006, 21, 1595–1606. [Google Scholar] [CrossRef] [PubMed]
- Betarbet, R.; Sherer, T.B.; MacKenzie, G.; Garcia-Osuna, M.; Panov, A.V.; Greenamyre, J.T. Chronic systemic pesticide exposure reproduces features of Parkinson’s disease. Nat. Neurosci. 2000, 3, 1301–1306. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, W.J.; Alam, M. Controversies on new animal models of Parkinson’s disease pro and con: The rotenone model of Parkinson’s disease (PD). J. Neural Transm. Suppl. 2006, 70, 273–276. [Google Scholar]
- Tole, S.; Christian, C.; Grove, E.A. Early specification and autonomous development of cortical fields in the mouse hippocampus. Development 1997, 124, 4959–4970. [Google Scholar] [PubMed]
- Lee, S.M.; Tole, S.; Grove, E.; McMahon, A.P. A local Wnt-3a signal is required for development of the mammalian hippocampus. Development 2000, 127, 457–467. [Google Scholar] [PubMed]
- Tole, S.; Grove, E.A. Detailed field pattern is intrinsic to the embryonic mouse hippocampus early in neurogenesis. J. Neurosci. 2001, 21, 1580–1589. [Google Scholar] [PubMed]
- Altman, J.; Bayer, S.A. Atlas of Prenatal Rat Brain Development; CRC: Boca Raton, FL, USA, 1995. [Google Scholar]
- Huang, H.; Liu, C.M.; Sun, J.; Hao, T.; Xu, C.M.; Wang, D.; Wu, Y.Q. Ketamine Affects the Neurogenesis of the Hippocampal Dentate Gyrus in 7-Day-Old Rats. Neurotox. Res. 2016, 30, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Voorn, P.; Kalsbeck, A.; Jorritsma-Byham, B.; Groenewegen, H.J. The pre- and postnatal development of the dopaminergic cell groups in the ventral mesencephalon and the dopaminergic innervation of the stria- tum of the rat. Neuroscience 1988, 25, 857–887. [Google Scholar] [CrossRef]
- Tepper, J.M.; Damlama, M.; Trent, F. Postnatal changes in the dis- tribution and morphology of rat substantia Nigra dopaminergic neurons. Neuroscience 1994, 60, 469–477. [Google Scholar] [CrossRef]
- Schmidt, U.; Beyer, C.; Oestreicher, A.B.; Reisert, I.; Schilling, K.; Pilgrim, C. Activation of dopaminergic D1 receptors promotes morphogenesis of developing striatal neurons. Neuroscience 1996, 74, 453–460. [Google Scholar] [CrossRef]
- Spencer, G.E.; Klumperman, J.; Syed, N.I. Neurotransmitters and neurodevelopment. Role of dopamine in neurite outgrowth, target selection and specific synapse formation. Perspect. Dev. Neurobiol. 1998, 5, 451–467. [Google Scholar] [PubMed]
- Stanwood, G.; Levitt, P. The effects of cocaine on the developing nervous system. In Handbook of Developmental Cognitive Neuroscience; Nelson, C.A., Luciana, M., Eds.; MIT Press: Cambridge, MA, USA, 2001; pp. 519–536. [Google Scholar]
- Bellone, C.; Mameli, M.; Luscher, C. In Utero exposure to cocaine delays postnatal synaptic maturation of glutamatergic transmission in the VTA. Nat. Neurosci. 2011, 14, 1439–1446. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, D.M.; Zhang, X.; Darnell, S.B.; Sangrey, G.R.; Yanagawa, Y.; Sadri-Vakili, G. Cocaine alters BDNF expression and neuronal migration in the embryonic mouse forebrain. J. Neurosci. 2011, 31, 13400–13411. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.M.; Barnett, A.; Malick, J.B. The evaluation of anti-Parkinson drugs on reserpine-induced rigidity in rats. Eur. J. Pharmacol. 1975, 33, 183–188. [Google Scholar] [CrossRef]
- Green, A.R.; Backus, L.I. Animal models of serotonin behavior. Ann. N. Y. Acad. Sci. 1990, 600, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, J.L.; Reid, J.J. Effects of methylenedioxymethamphetamine on the release of monoamines from rat brain slices. Eur. J. Pharmacol. 1990, 191, 217–220. [Google Scholar] [CrossRef]
- Eiden, L.E.; Weihe, E. VMAT2: A dynamic regulator of brain monoaminergic neuronal function interacting with drugs of abuse. Ann. N. Y. Acad. Sci. 2011, 1216, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Hamers, F.P.T.; Lankhorst, A.J.; Van Laar, T.J.; Veldhuis, W.B.; Gispen, W.H. Automatedquantitative gait analysis during overground locomotion inthe rat: Its application to spinal cord contusion and transec-tion injuries. J. Neurotrauma 2001, 18, 187–201. [Google Scholar] [CrossRef] [PubMed]
- Bakke, J.L.; Lawrence, N.L.; Robinson, S.A.; Bennett, J.; Bowers, C. Late endocrine effects of l-dopa, 5-HTP, and 6-OH-dopa administered to neonatal rats. Neuroendocrinology 1978, 25, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Glazova, N.Y.; Merchieva, S.A.; Volodina, M.A.; Sebentsova, E.A.; Manchenko, D.M.; Kudrun, V.S.; Levitskaya, N.G. Effects of Neonatal Fluvoxamine Administration on the Physical Development and Activity of the Serotoninergic System in White Rats. Acta Nat. 2014, 6, 98–105. [Google Scholar]
- Meyerson, B.J. Influence of early beta-endorphin treatment on the behavior and reaction to beta-endorphin in the adult male rat. Psychoneuroendocrinology 1985, 10, 135–147. [Google Scholar] [CrossRef]
- Kranick, S.M.; Duda, J.E. Olfactory dysfunction in Parkinson’s disease. Neurosignals 2008, 16, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Driver-Duckley, E.; Adler, C.H.; Hentz, J.G.; Dugger, B.N.; Shill, H.A.; Caviness, J.N.; Sabbagh, M.N.; Beach, T.G.; Arizona Parkinson Disease Consortium. Olfactory dysfunction in incidental Lewy body disease and Parkinson’s disease. Parkinsonism Relat. Disord. 2014, 20, 1260–1262. [Google Scholar]
- Alves, J.; Petrosyan, A.; Magalhaes, R. Olfactory disfunction in dementia. World J. Clin. Cases 2014, 2, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Bohnen, N.I.; Gedela, S.; Herath, P. Selective hyposmia in Parkinson disease: Association with hippocampal dopamine activity. Neurosci. Lett. 2008, 447, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Mega, M.S.; Cummings, J.L.; Fiorello, T.; Gornbein, J. The spectrum of behavioral changes in Alzheimer’s Disease. Neurol 1996, 46, 130–135. [Google Scholar] [CrossRef]
- Menza, M.A.; Robertson-Hoffman, D.E.; Bonapace, A.S. Parkinson’s disease and anxiety: Comorbidity with depression. Biol. Psychiatry 1993, 34, 465–470. [Google Scholar] [CrossRef]
- Lee, I.; Hunsaker, M.R.; Kesner, R.P. The Role of Hippocampal Subregions in Detecting Spatial Novelty. Behav. Neurosci. 2005, 119, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Saab, B.J.; Georgiou, J.; Nath, A.; Lee, F.J.S.; Wang, M.; Michalon, A.; Liu, F.; Mansuy, I.M.; Roder, J.C. NCS-1 in the Dentate Gyrus Promotes Exploration, Synaptic Plasticity, and Rapid Acquisition of Spatial Memory. Neuron 2009, 63, 643–656. [Google Scholar] [CrossRef] [PubMed]
- Lever, C.; Burton, S.; O’Keefe, J. Rearing on hind legs, environmental novelty, and the hippocampal formation. Rev. Neurosci. 2006, 17, 111–133. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.C. Neurocognitive effects of methamphetamine: A critical review and meta-analysis. Neuropsychol. Rev. 2007, 17, 275–297. [Google Scholar] [CrossRef] [PubMed]
- Tirelli, E.; Jodogne, C. Dopamine-GABAergic mechanisms of rearing and locomotion in infant and weanling mice. Psychobiology 1990, 18, 443–450. [Google Scholar]
- Weinshenker, D.; Warren, S.T. Fragile dopamine. Nature 2008. [Google Scholar] [CrossRef] [PubMed]
- Russell, K.H.; Giordano, M.; Sanberg, P.R. Amphetamine-induced on- and off-wall rearing in adult laboratory rats. Pharmacol. Biochem. Behav. 1987, 26, 7–10. [Google Scholar] [CrossRef]
- Menon, M.K.; Clark, W.G. GABA-ergic drugs block the locomotor stimulant effects of 1,3-dimethyl-5-aminoadamantane (D-145). Neuropharmacology 1979, 18, 223–225. [Google Scholar] [CrossRef]
- Scheel-Kruger, J.; Christensen, A.V.; Arnt, J. Muscimol differentially facilitates stereotypy but antagonizes motility induced by dopaminergic drugs: A complex GABA-dopamine interaction. Life Sci. 1978, 22, 75–84. [Google Scholar] [CrossRef]
- Callaghan, R.C.; Cunningham, J.K.; Sykes, J.; Kish, S.J. Increased risk of Parkinson’s disease in individuals hospitalized with conditions related to the use of methamphetamine or other amphetamine-type drugs. Drug Alcohol Depend. 2012, 120, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Rudnicki, S.A.; Archer, R.L.; Labib, B.T. Motor neuron disease in methamphetamine abusers. Amyotroph. Lateral Scler. 2007, 8, 126–127. [Google Scholar] [CrossRef] [PubMed]
- NIH-National Institute on Drug Abuse. Methamphetamine Abuse Linked to Long-Term Damage to Brain Cells; NIH-National Institute on Drug Abuse: Rockville, MD, USA, 2000.
- Walf, A.A.; Frye, C.A. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Marcondes, F.K.; Miguel, K.J.; Melo, L.L.; Spadari-Bratfisch, R.C. Estrous cycle influences the response of female rats in the elevated plus-maze test. Physiol. Behav. 2001, 74, 435–440. [Google Scholar] [CrossRef]
- Hard, E.; Engel, J.; Larsson, K.; Musi, B. Effect of Diazepam, Apomorphine and Haloperidol on the audiogenic immobility reaction and on the open field behavior. Psychopharmacology 1985, 85, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Hard, E.; Ahlenius, S.; Engel, J. Effects of neonatal treatment with 5,7-dihydroxytryptamine or 6-hydroxydopamine on the ontogenetic development of the audiogenic immobility reaction in the rat. Psychopharmacology 1983, 80, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Stopford, C.L.; Thompson, J.C.; Neary, D.; Richardson, A.M.; Snowden, J.S. Working memory, attention, and executive function in Alzheimer’s disease and frontotemporal dementia. Cortex 2012, 48, 429–446. [Google Scholar] [CrossRef] [PubMed]
- Hodges, H. Maze procedures: The radial-arm and water maze compared. Brain Res. Cogn. Brain Res. 1996, 3, 167–181. [Google Scholar] [CrossRef]
- Clark, R.E.; Broadbent, N.J.; Squire, L.R. The hippocampus and spatial memory: Findings with a novel modification of the water maze. J. Neurosci. 2007, 27, 6647–6654. [Google Scholar] [CrossRef] [PubMed]
- Buenz, E.J.; Howe, C.L. Beta-methylamino-alanine (BMAA) injures hippocampal neurons in vivo. Neurotoxicology 2007, 28, 702–704. [Google Scholar] [CrossRef] [PubMed]
- Terry, A.V., Jr. Chapter 13: Spatial Navigation (Water Maze) Tasks in Methods of Behaviour Analysis in Neuroscience, 2nd ed.; Buccafusco, J.J., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2009. [Google Scholar]
- Bromley-Brits, K.; Deng, Y.; Song, W. Morris Water Maze Test for Learning and Memory Deficits in Alzheimer’s Disease Model Mice. J. Vis. Exp. 2011, e2920. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, R.J.; Hoesing, J.M. Posterior cingulate cortex and spa- tial memory: A microlimnology analysis. In Neurobiology of Cingulate Cortex and Limbic Thalamus: A Comprehensive Handbook; Vogt, B.A., Gabriel, M., Eds.; Birkhauser: Boston, MA, USA, 1993; pp. 461–477. [Google Scholar]
- Voorhees, C.V.; Williams, M.T. Assessing spatial learning and memory in rodents. ILAR 2014, 55, 310–332. [Google Scholar] [CrossRef] [PubMed]
- Santiago, M.; Matarredona, E.R.; Machado, A.; Cano, J. Acute perfusion of BMAA in the rat’s striatum by in vivo microdialysis. Toxicol. Lett. 2006, 167, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Lindstrom, H.; Luthman, J.; Mouton, P.; Spencer, P.; Olson, L. Plant-derived neurotoxic amino acid (beta-N-oxalylamino-l-alanine and beta-N-methylamino-l-alanine): Effects on central monoamine neurons. J. Neurochem. 1990, 55, 941–999. [Google Scholar] [CrossRef] [PubMed]
- Herlenius, E.; Langercrantz, H. Development of neurotransmitter systems during critical periods. Exp. Neurol. 2004, 190, S8–S21. [Google Scholar] [CrossRef] [PubMed]
- Clancy, B.; Kersh, B.; Hyde, J.; Darlington, R.B.; Anand, K.J.S.; Finlay, B.L. Web-based method for translating neurodevelopment from laboratory species to humans. Neuroinformatics 2007, 5, 79–94. [Google Scholar] [CrossRef]
- Banack, S.A.; Metcalf, J.S.; Spáčil, Z.; Downing, T.G.; Downing, S.; Long, A.; Nunn, P.B.; Cox, P.A. Distinguishing the cyanobacterial neurotoxin β-N-methylamino-l-alanine (BMAA) from other diamino acids. Toxicon 2011, 57, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Lamprea, M.R.; Cardenas, F.P.; Setem, J.; Morato, S. Thigmotactic responses in an open field. Braz. J. Med. Biol. Res. 2008, 41, 135–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, K.R.; Crawley, J.N. Anxiety-related disorders in Mice; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2009. [Google Scholar]
- Conn, P.M. Paradigms for the Study of Behaviour. Methods in Neuroscience; Academic Press, Inc.: San Diego, CA, USA, 1993. [Google Scholar]
- Roos, M.W.; Ericsson, A.; Berg, M.; Sperber, G.O.; Sjoquist, M.; Meyerson, B.J. Functional evaluation of cerebral microembolization in the rat. Brain Res. 2003, 961, 15–21. [Google Scholar] [CrossRef]
- Olton, D.S.; Samuelson, R.J. Remembrance of places passed: Spatial memory in rats. J. Exp. Psychol. 1976, 2, 97–116. [Google Scholar] [CrossRef]
- Wenk, G.L. Assessment of spatial memory using the radial arm maze and Morris Water Maze. In Current Protocols in Neuroscience; Gerfen, C., Holmes, A., Sibley, D., Skolnick, P., Wray, S., Eds.; Wiley & Sons, Inc.: New York, NY, USA, 2004. [Google Scholar]
- Penley, S.C.; Gaudet, C.M.; Threlkeld, S.W. Use of an Eight-arm Radial Water Maze to Assess Working and Reference Memory Following Neonatal Brain Injury. J. Vis. Exp. 2013, e50940. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Group | Trial 1 | Trial 2 | Trial 3 |
|---|---|---|---|
| Control | 100% | 100% | 100% |
| Vehicle control | 100% | 100% | 100% |
| G14 | 100% | 80% | 90% |
| PND 3 | 60% | 60% | 60% |
| PND 4 | 60% | 70% | 70% |
| PND 5 | 60% | 60% | 80% |
| PND 6 | 80% | 80% | 80% |
| PND 7 | 100% | 100% | 80% |
| PND 10 | 90% | 90% | 100% |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scott, L.L.; Downing, T.G. β-N-Methylamino-L-alanine (BMAA) Toxicity Is Gender and Exposure-Age Dependent in Rats. Toxins 2018, 10, 16. https://doi.org/10.3390/toxins10010016
Scott LL, Downing TG. β-N-Methylamino-L-alanine (BMAA) Toxicity Is Gender and Exposure-Age Dependent in Rats. Toxins. 2018; 10(1):16. https://doi.org/10.3390/toxins10010016
Chicago/Turabian StyleScott, Laura Louise, and Timothy Grant Downing. 2018. "β-N-Methylamino-L-alanine (BMAA) Toxicity Is Gender and Exposure-Age Dependent in Rats" Toxins 10, no. 1: 16. https://doi.org/10.3390/toxins10010016
APA StyleScott, L. L., & Downing, T. G. (2018). β-N-Methylamino-L-alanine (BMAA) Toxicity Is Gender and Exposure-Age Dependent in Rats. Toxins, 10(1), 16. https://doi.org/10.3390/toxins10010016