Biological and Proteolytic Variation in the Venom of Crotalus scutulatus scutulatus from Mexico


 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Mojave Toxin Detection via PCR
2.2. Mojave Toxin Detection by ELISA
2.3. Median Lethal Dose (LD)
2.4. Hemorrhagic Activity
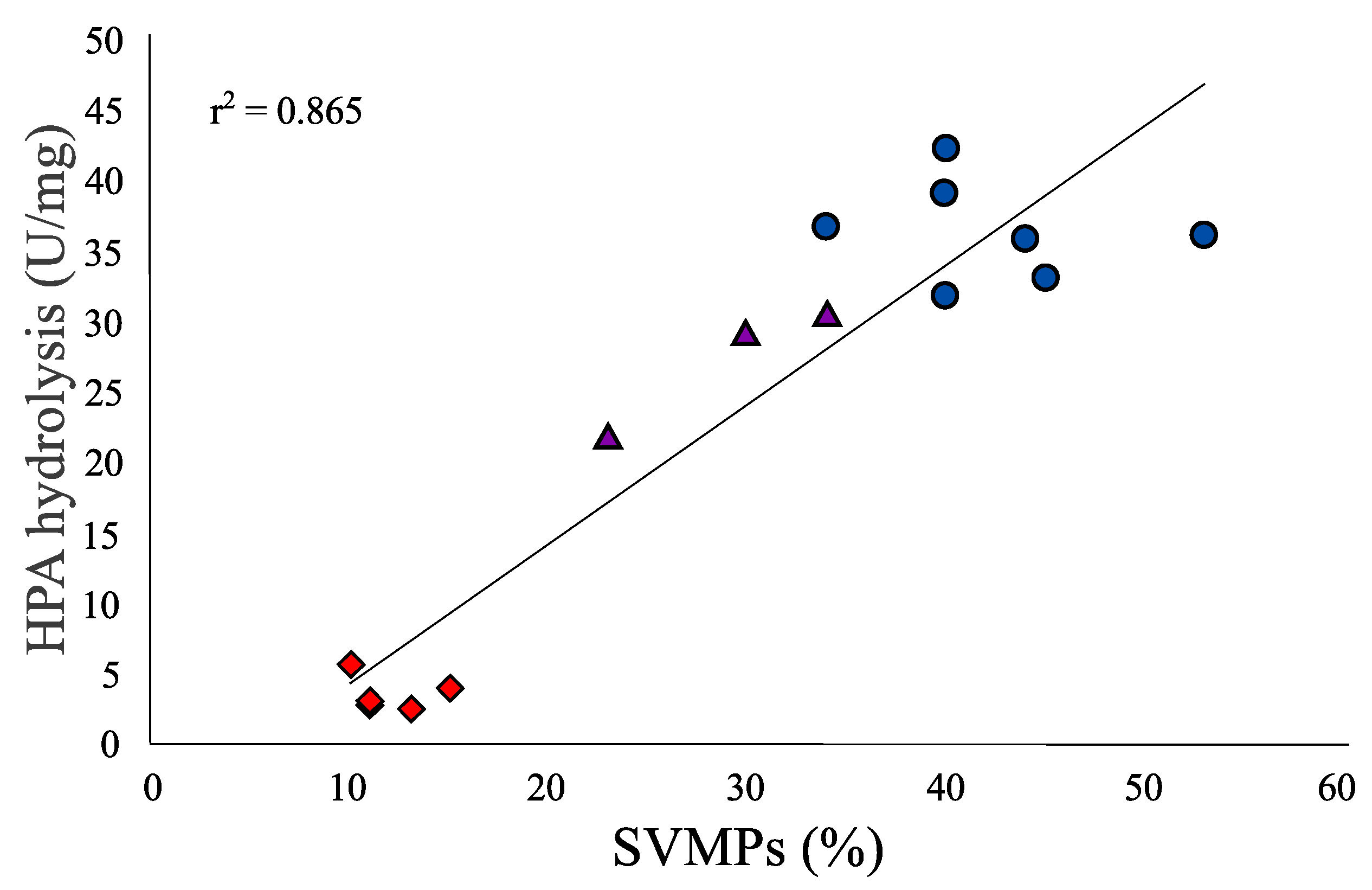
2.5. Hide Powder Azure (HPA) Hydrolysis
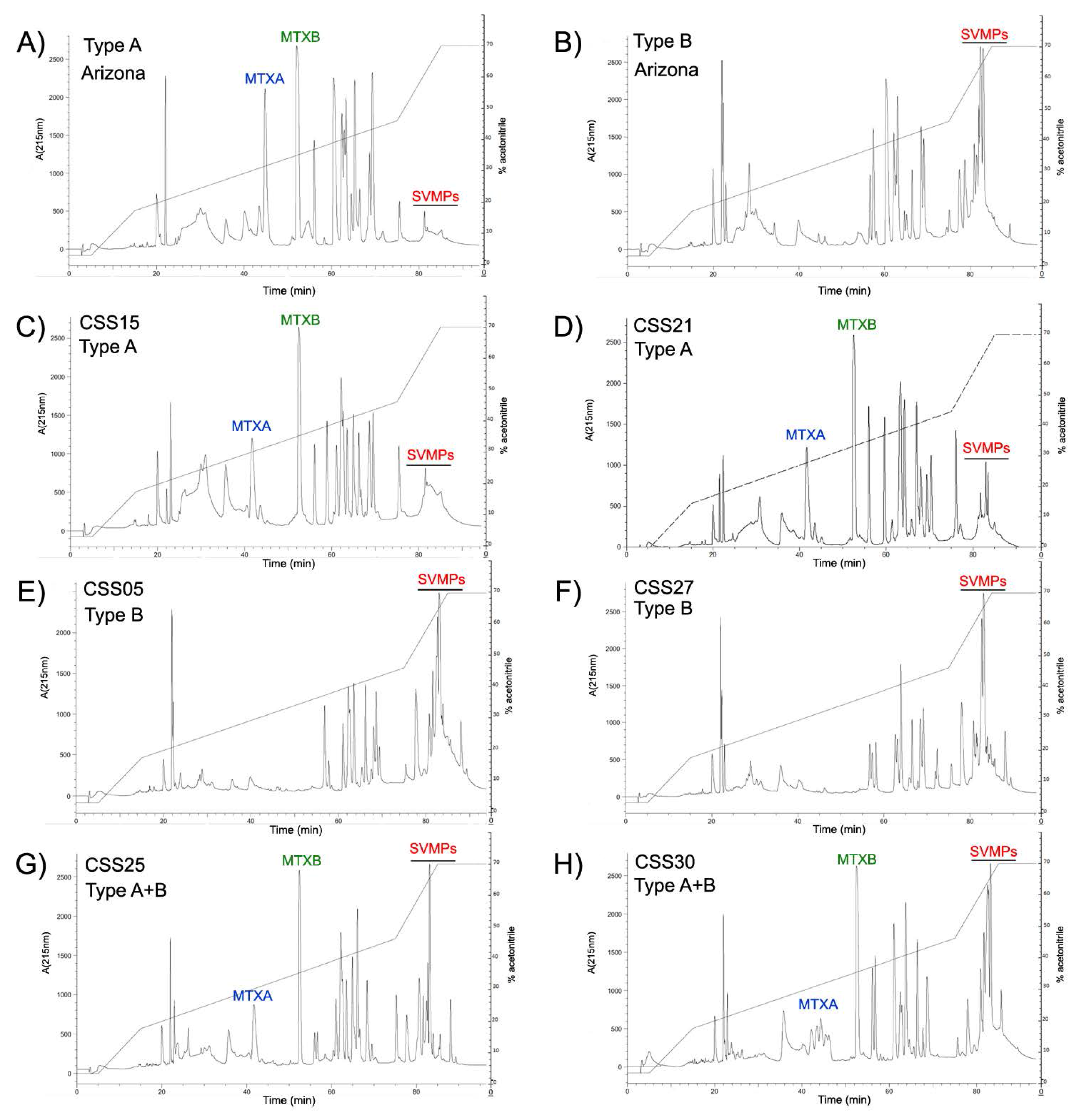
2.6. Reverse Phase HPLC for MTX and SVMPs Detection
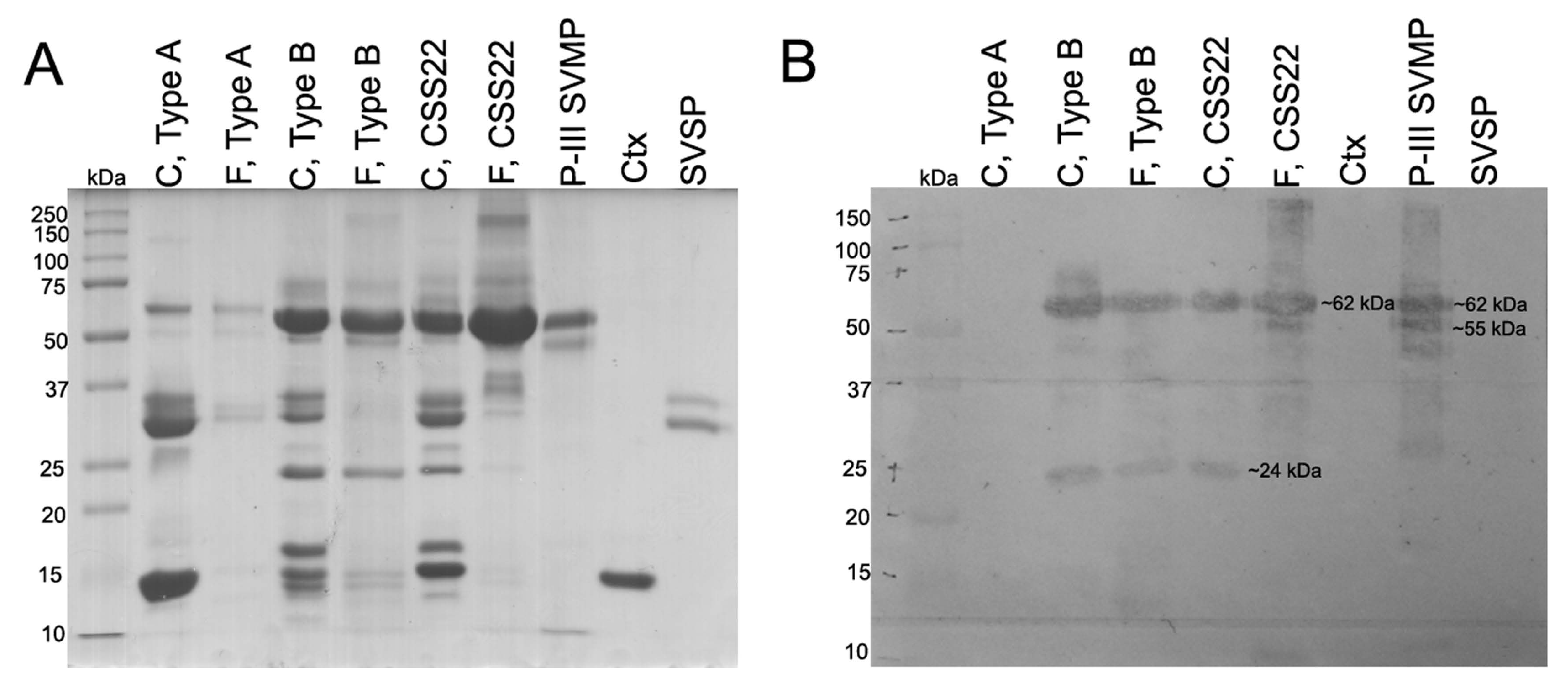
2.7. SVMPs Detection by Western Blot
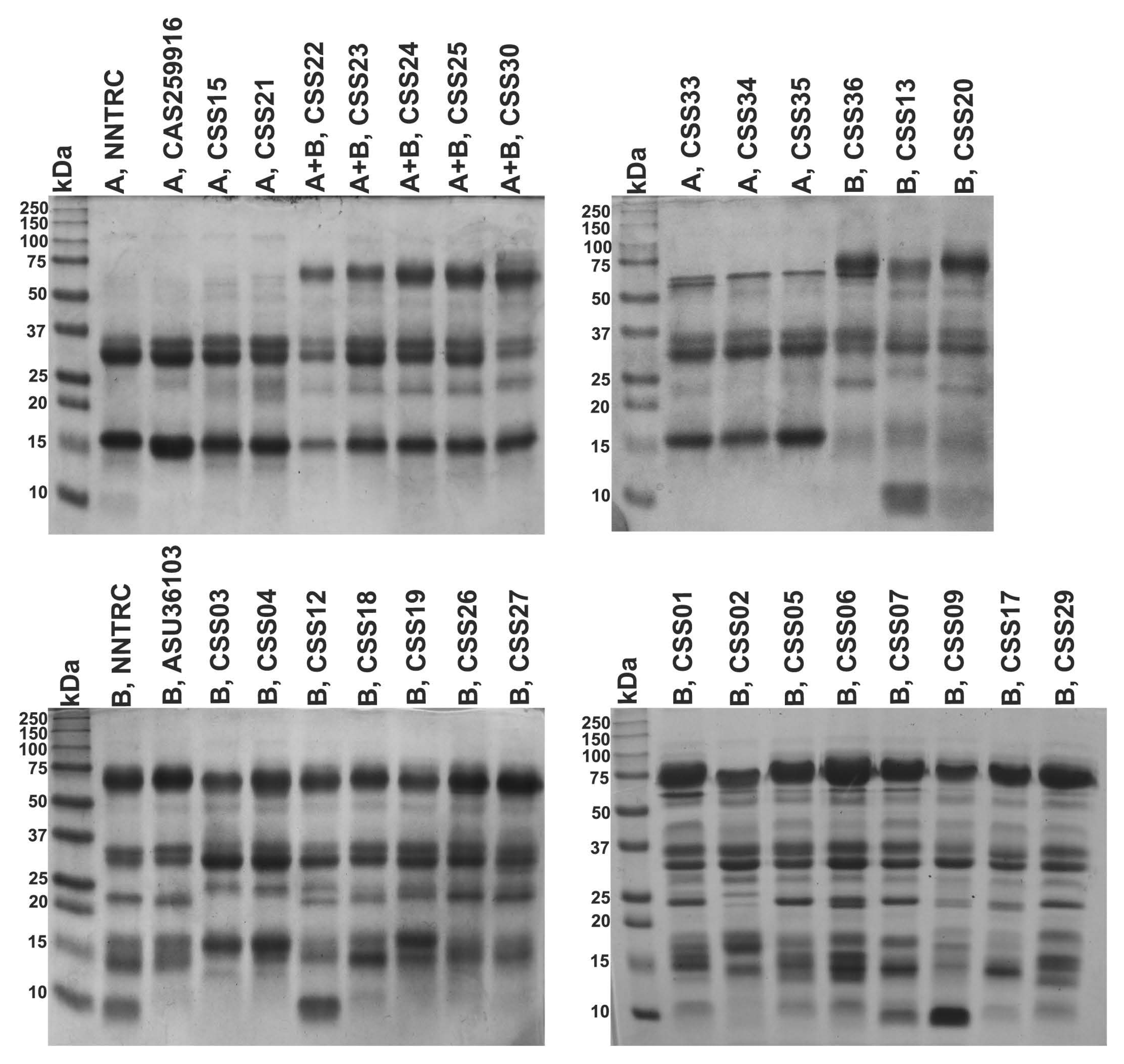
2.8. SDS-PAGE
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Ethics Statement
5.2. Crotalus scutulatus scutulatus Sampling
5.3. PCR Detection of Mojave Toxin Genes
5.4. Protein Concentration Determination
5.5. Mojave Toxin Detection by Sandwich ELISA
5.6. Median Lethal Dose (LD)
5.7. Hemorrhagic Activity
5.8. Hide Powder Azure (HPA) Hydrolysis
5.9. Reverse Phase HPLC for MTX and SVMPs Detection
5.10. SVMP Detection by Western Blot
5.11. SDS-PAGE
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| Ags. | Aguascalientes |
| Chih. | Chihuahua |
| Coah. | Coahuila |
| CSS | Crotalus scutulatus scutulatus |
| ELISA | Enzyme-linked Immunosorbent Assay |
| HPA | Hide Powder Azure |
| LD | Median Lethal Dose 50 |
| MDPI | Multidisciplinary Digital Publishing Institute |
| MTX | Mojave toxin |
| MTXA | Acidic subunit of Mojave toxin |
| MTXB | Basic subunit of Mojave toxin |
| ND | No determined due to lack of venom available |
| NNTRC | National Natural Toxins Research Center |
| PCR | Polymerase Chain Reaction |
| PLA | Phospholipase A |
| RP-HPLC | Reverse-Phased High Performance Liquid Chromatography |
| SDS-PAGE | Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis |
| SVMP | Snake Venom Metalloproteinases |
| SVSP | Snake Venom Serine Protease |
| UCF | University of Central Florida |
| UJED | Universidad Juárez del Estado de Durango |
| UNAM | Universidad Nacional Autónoma de México |
Appendix A. RP-HPLC Chromatograms

References
- Kang, T.S.; Georgieva, D.; Genov, N.; Murakami, M.T.; Sinha, M.; Kumar, R.P.; Kaur, P.; Kumar, S.; Dey, S.; Sharma, S.; et al. Enzymatic toxins from snake venom: Structural characterization and mechanism of catalysis. FEBS J. 2011, 278, 4544–4576. [Google Scholar] [CrossRef] [PubMed]
- Mackessy, S.P. Venom composition in rattlesnakes: Trends and biological significance. In The Biology of Rattlesnakes; Hayes, W.K., Beaman, K.R., Cardwell, M.D., Bush, S.P., Eds.; Loma Linda University Press: Loma Linda, CA, USA, 2008; pp. 495–510. [Google Scholar]
- Calvete, J.J.; Fasoli, E.; Sanz, L.; Boschetti, E.; Righetti, P.G. Exploring the venom proteome of the western diamondback rattlesnake, Crotalus atrox, via snake venomics and combinatorial peptide ligand library approaches. J. Proteome Res. 2009, 8, 3055–3067. [Google Scholar] [CrossRef] [PubMed]
- Boldrini-França, J.; Corrêa-Netto, C.; Silva, M.M.; Rodrigues, R.S.; De La Torre, P.; Pérez, A.; Soares, A.M.; Zingali, R.B.; Nogueira, R.A.; Rodrigues, V.M.; et al. Snake venomics and antivenomics of Crotalus durissus subspecies from Brazil: Assessment of geographic variation and its implication on snakebite management. J. Proteom. 2010, 73, 1758–1776. [Google Scholar] [CrossRef] [PubMed]
- Massey, D.J.; Calvete, J.J.; Sánchez, E.E.; Sanz, L.; Richards, K.; Curtis, R.; Boesen, K. Venom variability and envenoming severity outcomes of the Crotalus scutulatus scutulatus (Mojave rattlesnake) from Southern Arizona. J. Proteom. 2012, 75, 2576–2587. [Google Scholar] [CrossRef] [PubMed]
- Tasoulis, T.; Isbister, G.K. A review and database of snake venom proteomes. Toxins 2017, 9, 290. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J.; Sanz, L.; Cid, P.; de la Torre, P.; Flores-Díaz, M.; Dos Santos, M.C.; Borges, A.; Bremo, A.; Angulo, Y.; Lomonte, B.; et al. Snake venomics of the Central American rattlesnake Crotalus simus and the South American Crotalus durissus complex points to neurotoxicity as an adaptive paedomorphic trend along Crotalus dispersal in South America. J. Proteome Res. 2010, 9, 528–544. [Google Scholar] [CrossRef] [PubMed]
- Doley, R.; Kini, R.M. Protein complexes in snake venom. Cell. Mol. Life Sci. 2009, 66, 2851–2871. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J.; Pérez, A.; Lomonte, B.; Sánchez, E.E.; Sanz, L. Snake venomics of Crotalus tigris: The minimalist toxin arsenal of the deadliest neartic rattlesnake venom. Evolutionary clues for generating a pan-specific antivenom against crotalid type II venoms. J. Proteome Res. 2012, 11, 1382–1390. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.; Lee, C. Presynaptic actions of Mojave toxin isolated from Mojave rattlesnake (Crotalus scutulatus) venom. Toxicon 1981, 19, 889–892. [Google Scholar] [CrossRef]
- Takeda, S.; Takeya, H.; Iwanaga, S. Snake venom metalloproteinases: Structure, function and relevance to the mammalian ADAM/ADAMTS family proteins. Biochim. Biophys. Acta 2012, 1824, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Rucavado, A.; Escalante, T. Snake venom metalloproteinases: Biological roles and participation in the pathophysiology of envenomation. In Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2010; pp. 115–138. [Google Scholar]
- Escalante, T.; Rucavado, A.; Fox, J.W.; Gutiérrez, J.M.; Rica, C. Key events in microvascular damage induced by snake venom hemorrhagic metalloproteinases. J. Proteom. 2011, 74, 1781–1794. [Google Scholar] [CrossRef] [PubMed]
- Ramos, O.; Selistre-de Araujo, H. Snake venom metalloproteases—Structure and function of catalytic and disintegrin domains. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2006, 142, 328–346. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.W.; Serrano, S.M. Snake venom metalloproteinases. In Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2010; pp. 95–113. [Google Scholar]
- Cate, R.L.; Bieber, A.L. Purification and characterization of mojave (Crotalus scutulatus scutulatus) toxin and its subunits. Arch. Biochem. Biophys. 1978, 189, 397–408. [Google Scholar] [CrossRef]
- Bieber, A.L.; Tu, T.; Tu, A.T. Studies of an acidic cardiotoxin isolated from the venom of mojave rattlesnake (Crotalus scutulatus). Biochim. Biophys. Acta 1975, 400, 178–188. [Google Scholar] [CrossRef]
- Gopalakrishnakone, P.; Hawgood, B.J.; Holbrooke, S.E.; Marsh, N.A.; Santana De Sa, S.; Tu, A.T. Sites of action of Mojave toxin isolated from the venom of the Mojave rattlesnake. Br. J. Pharmacol. 1980, 69, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Valdes, J.J.; Thompson, R.G.; Wolff, V.L.; Menking, D.E.; Rael, E.D.; Chambers, J.P. Inhibition of calcium channel dihydropyridine receptor binding by purified Mojave toxin. Neurotoxicol. Teratol. 1989, 11, 129–133. [Google Scholar] [CrossRef]
- John, T.R.; Smith, L.A.; Kaiser, I.I. Genomic sequences encoding the acidic and basic subunits of Mojave toxin: Unusually high sequence identity of non-coding regions. Gene 1994, 139, 229–234. [Google Scholar] [CrossRef]
- Zancolli, G.; Baker, T.; Barlow, A.; Bradley, R.; Calvete, J.; Carter, K.; de Jager, K.; Owens, J.; Price, J.; Sanz, L.; et al. Is hybridization a source of adaptive venom Variation in rattlesnakes? A test, using a Crotalus scutulatus x viridis hybrid zone in southwestern New Mexico. Toxins 2016, 8, 188. [Google Scholar] [CrossRef] [PubMed]
- Bieber, A.L.; Becker, R.R.; McParland, R.; Hunt, D.F.; Shabanowitz, J.; Yates, J.R.; Martino, P.A.; Johnson, G.R. The complete sequence of the acidic subunit from Mojave toxin determined by Edman degradation and mass spectrometry. Biochim. Biophys. Acta 1990, 1037, 413–421. [Google Scholar] [CrossRef]
- Bouchier, C.; Ducancel, F.; Guignery-Frelat, G.; Bon, C.; Boulain, J.C.; Ménez, A. Cloning and sequencing of cDNAs encoding the two subunits of Crotoxin. Nucleic Acids Res. 1988, 16, 9050. [Google Scholar] [CrossRef] [PubMed]
- Aird, S.D.; Kruggel, W.G.; Kaiser, I.I. Amino acid sequence of the basic subunit of Mojave toxin from the venom of the Mojave rattlesnake (Crotalus s. scutulatus). Toxicon 1990, 28, 669–673. [Google Scholar] [CrossRef]
- Henderson, J.T.; Bieber, A.L. Antigenic relationships between Mojave toxin subunits, Mojave toxin and some crotalid venoms. Toxicon 1986, 24, 473–479. [Google Scholar] [CrossRef]
- Rael, E.D.; Salo, R.J.; Zepeda, H. Monoclonal antibodies to Mojave toxin and use for isolation of cross-reacting proteins in Crotalus venoms. Toxicon 1986, 24, 661–668. [Google Scholar] [CrossRef]
- Neri-Castro, E.; Lomonte, B.; del Carmen Gutiérrez, M.; Alagón, A.; Gutiérrez, J.M. Intraspecies variation in the venom of the rattlesnake Crotalus simus from Mexico: Different expression of crotoxin results in highly variable toxicity in the venoms of three subspecies. J. Proteom. 2013, 87, 103–121. [Google Scholar] [CrossRef] [PubMed]
- Borja, M.; Lazcano, D.; Martínez-Romero, G.; Morlett, J.; Sánchez, E.; Cepeda-Nieto, A.C.; Garza-García, Y.; Zugasti-Cruz, A. Intra-specific variation in the protein composition and proteolytic activity of venom of Crotalus lepidus morulus from the northeast of Mexico. Copeia 2013, 2013, 707–716. [Google Scholar] [CrossRef]
- Sunagar, K.; Undheim, E.A.B.; Scheib, H.; Gren, E.C.K.; Cochran, C.; Person, C.E.; Koludarov, I.; Kelln, W.; Hayes, W.K.; King, G.F.; et al. Intraspecific venom variation in the medically significant Southern Pacific Rattlesnake (Crotalus oreganus helleri): Biodiscovery, clinical and evolutionary implications. J. Proteom. 2014, 99, 68–83. [Google Scholar] [CrossRef] [PubMed]
- Rokyta, D.R.; Wray, K.P.; McGivern, J.J.; Margres, M.J. The transcriptomic and proteomic basis for the evolution of a novel venom phenotype within the Timber Rattlesnake (Crotalus horridus). Toxicon 2015, 98, 34–48. [Google Scholar] [CrossRef] [PubMed]
- Glenn, J.J.L.; Straight, R.C.; Wolfe, M.M.C.; Hardy, D.D.L. Geographical variation in Crotalus scutulatus scutulatus (Mojave rattlesnake) venom properties. Toxicon 1983, 21, 119–130. [Google Scholar] [CrossRef]
- Glenn, J.L.; Straight, R.C. Intergradation of two different venom populations of the Mojave rattlesnake (Crotalus scutulatus scutulatus) in Arizona. Toxicon 1989, 27, 411–418. [Google Scholar] [CrossRef]
- Sánchez, E.E.; Galán, J.A.; Powell, R.L.; Reyes, S.R.; Soto, J.G.; Russell, W.K.; Russell, D.H.; Pérez, J.C. Disintegrin, hemorrhagic, and proteolytic activities of Mohave rattlesnake, Crotalus scutulatus scutulatus venoms lacking Mojave toxin. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2005, 141, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.A.; Lamar, W.W. The venomous reptiles of the Western Hemisphere; Comstock Publishing Associates: Ithaca, NY, USA, 2004; p. 870. [Google Scholar]
- Borja, M.; Castañeda, G.; Espinosa, J.; Neri, E.; Carbajal, A.; Clement, H.; García, O.; Alagon, A. Mojave Rattlesnake (Crotalus scutulatus scutulatus) with type B venom from Mexico. Copeia 2014, 2014, 7–13. [Google Scholar] [CrossRef]
- Dowell, N.L.; Giorgianni, M.W.; Kassner, V.A.; Selegue, J.E.; Sanchez, E.E.; Carroll, S.B. The deep origin and recent loss of venom toxin genes in rattlesnakes. Curr. Biol. 2016, 26, 2434–2445. [Google Scholar] [CrossRef] [PubMed]
- Wooldridge, B.; Pineda, G.; Banuelas-Ornelas, J.; Dagda, R.; Gasanov, S.; Rael, E.; Lieb, C. Mojave rattlesnakes (Crotalus scutulatus scutulatus) lacking the acidic subunit DNA sequence lack Mojave toxin in their venom. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2001, 130, 169–179. [Google Scholar] [CrossRef]
- Powell, R.L.; Lieb, C.S.; Rael, E.D. Geographic distribution of Mojave toxin and Mojave toxin subunits among selected Crotalus species. In The Biology of Rattlesnakes; Hayes, W.K., Beaman, K.R., Cardwell, M.D., Bush, S.P., Eds.; Loma Linda University Press: Loma Linda, CA, USA, 2008; pp. 537–550. [Google Scholar]
- Wilkinson, J.A.; Glenn, J.L.; Straight, R.C.; Sites, J.W. Distribution and genetic variation in venom A and B populations of the Mojave rattlesnake (Crotalus scutulatus scutulatus) in Arizona. Herpetologica 1991, 47, 54–68. [Google Scholar]
- Durban, J.; Pérez, A.; Sanz, L.; Gómez, A.; Bonilla, F.; Rodríguez, S.; Chacón, D.; Sasa, M.; Angulo, Y.; Gutiérrez, J.M.; et al. Integrated “omics” profiling indicates that miRNAs are modulators of the ontogenetic venom composition shift in the Central American rattlesnake, Crotalus simus simus. BMC Genom. 2013, 14, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dagda, R.K.; Gasanov, S.E.; De La OIII, Y.; Rael, E.D.; Lieb, C.S. Genetic basis for variation in metalloproteinase-associated biochemical activity in venom of the Mojave Rattlesnake (Crotalus scutulatus scutulatus). Biochem. Res. Int. 2013. [Google Scholar] [CrossRef] [PubMed]
- Margres, M.J.; Walls, R.; Suntravat, M.; Lucena, S.; Sánchez, E.E.; Rokyta, D.R.; Sanchez, E.E. Functional characterizations of venom phenotypes in the eastern diamondback rattlesnake (Crotalus adamanteus) and evidence for expression-driven divergence in toxic activities among populations. Toxicon 2016, 119, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; León, G.; Burnouf, T. Antivenoms for the treatment of snakebite envenomings: The road ahead. Biologicals 2011, 39, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Dobson, J.; Yang, D.; op den Brouw, B.; Cochran, C.; Huynh, T.; Kurrupu, S.; Sánchez, E.E.; Massey, D.J.; Baumann, K.; Jackson, T.N.; et al. Rattling the border wall: Pathophysiological implications of functional and proteomic venom variation between Mexican and US subspecies of the desert rattlesnake Crotalus scutulatus. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Powell, R.L.; Lieb, C.S. Perspectives on venom evolution in Crotalus. In The Biology of Rattlesnakes; Hayes, W.K., Beaman, K.R., Cardwell, M.D., Bush, S.P., Eds.; Loma Linda University Press: Loma Linda, CA, USA, 2008; pp. 551–556. [Google Scholar]
- Krasilnikov, P.; Gutiérrez-Castorena, M.d.C.; Ahrens, R.J.; Cruz-Gaistardo, C.O.; Sedov, S.; Solleiro-Rebolledo, E. The Soils of Mexico; World Soils Book Series; Springer: Dordrecht, The Netherlands, 2013. [Google Scholar]
- Marshall, C.J.; Liebherr, J.K. Cladistic biogeography of the Mexican transition zone. J. Biogeogr. 2000, 27, 203–216. [Google Scholar] [CrossRef]
- Bryson, R.W.; García-Vázquez, U.O.; Riddle, B.R. Phylogeography of Middle American gophersnakes: Mixed responses to biogeographical barriers across the Mexican Transition Zone. J. Biogeogr. 2011, 38, 1570–1584. [Google Scholar] [CrossRef]
- Rinderknecht, H.; Geokas, M.; Silverman, P.; Haverback, B. A new ultrasensitive method for the determination of proteolytic activity. Clin. Chim. Acta 1968, 21, 197–203. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
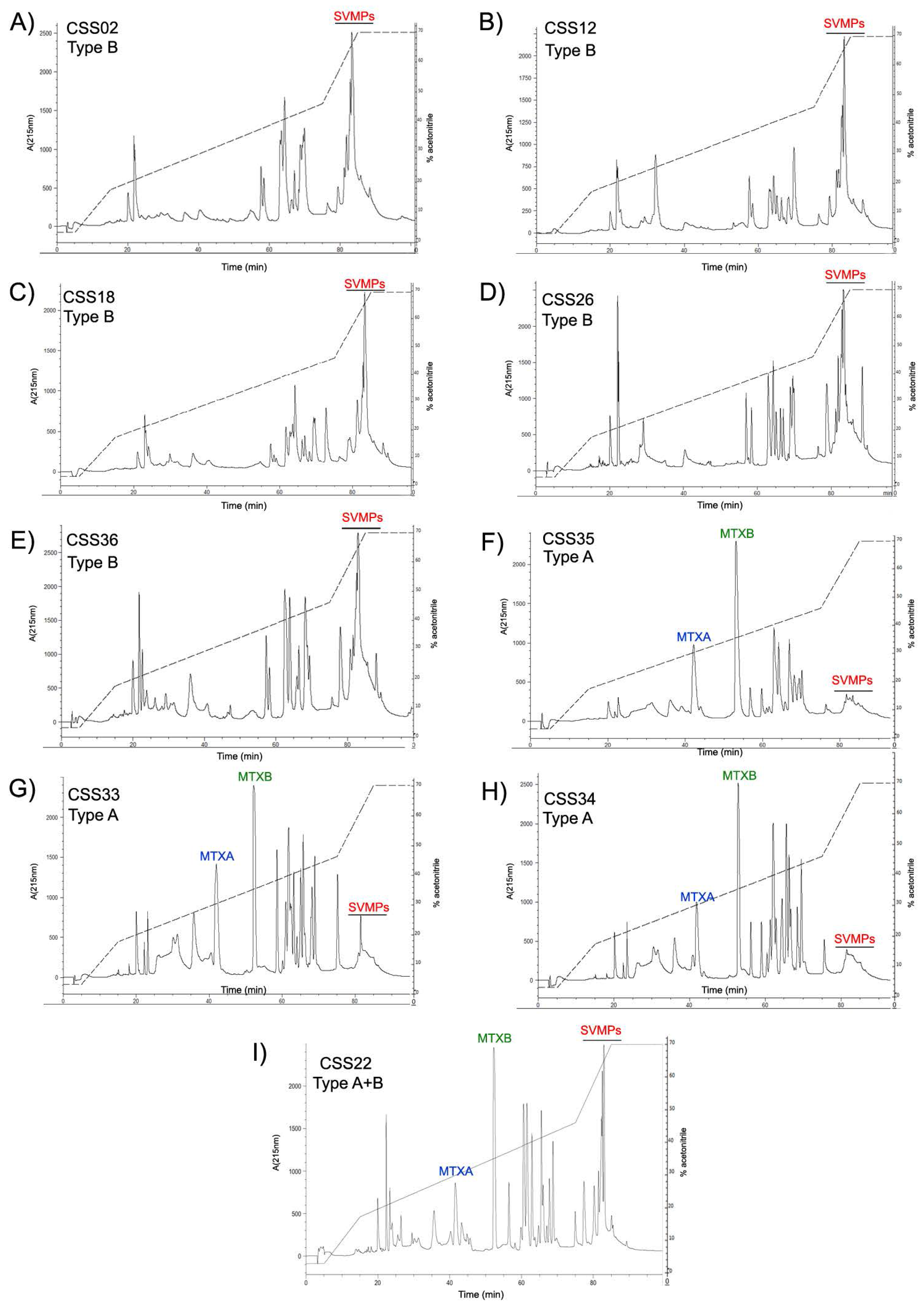{kind=link}
| ID | Geographic Location | Genomic Mojave Toxin Presence (MTXA/MTXB) | % of MTX in Venom | % of SVMPs in Venom (RP-HPLC) | LD (mg/kg) | Hemorrhagic Activity (Intensity) | HPA Hydrolysis (U/mg) |
|---|---|---|---|---|---|---|---|
| CSS01 | Aldama-Coyame, Chih. | No/No | 0 | ND | ND | 3 | 42.4 ± 1.8 |
| CSS02 | Aldama-Coyame, Chih. | No/No | 0 | 40 | ND | 4 | 39.3 ± 1.8 |
| CSS03 | Aldama-Coyame, Chih. | No/No | 0 | ND | ND | 4 | 46.5 ± 10.7 |
| CSS04 | Aldama-Coyame, Chih. | No/No | 0 | ND | 0.622 | 4 | 46.5 ± 3.1 |
| CSS26 | Aldama-Coyame, Chih. | No/No | 0 | 40 | 0.842 | 3 | 31.9 ± 0.6 |
| CSS27 | Aldama-Coyame, Chih. | No/No | 0 | 45 | 0.863 | 3 | 33.3 ± 2.4 |
| CSS09 | Matamoros, Coah. | No/No | 0 | ND | ND | 4 | 42.4 ± 1.8 |
| CSS12 | Matamoros, Coah. | No/No | 0 | 40 | 0.590 | 4 | 42.4 ± 1.8 |
| CSS13 | Matamoros, Coah. | No/No | 0 | ND | ND | ND | ND |
| CSS20 | Matamoros, Coah. | No/No | 0 | ND | 0.566 | 3 | 35.6 ± 1.9 |
| CSS06 | Tanque Aguilereño, Coah. | No/No | 0 | ND | ND | ND | 35.5 ± 1.1 |
| CSS17 | Tanque Aguilereño, Coah. | No/No | 0 | ND | ND | ND | ND |
| CSS18 | Tanque Aguilereño, Coah. | No/No | 0 | 44 | 0.300 | 4 | 36.0 ± 0.3 |
| CSS19 | Tanque Aguilereño, Coah. | No/No | 0 | ND | 0.584 | 3 | 32.1 ± 1.8 |
| CSS29 | Tanque Aguilereño, Coah. | No/No | 0 | ND | ND | 4 | 37.2 ± 3.1 |
| CSS05 | Peñón Blanco, Durango | No/No | 0 | 53 | 0.684 | 4 | 36.3 ± 0.6 |
| CSS07 | Mapimí, Durango | No/No | 0 | ND | ND | 4 | 42.1 ± 4.2 |
| CSS36 | Plateros, Zacatecas | No/No | 0 | 34 | 0.890 | 4 | 36.9 ± 0.9 |
| ASU36103 | Arizona, USA | No/No | 0 | 34 | 1.090 | 3 | 34.9 ± 0.2 |
| ID | Geographic Location | Genomic Mojave Toxin Presence (MTXA/MTXB) | % of MTX in Venom | % of SVMPs in Venom (RP-HPLC) | LD (mg/kg) | Hemorrhagic Activity (Intensity) | HPA Hydrolysis (U/mg) |
|---|---|---|---|---|---|---|---|
| CSS15 | Tepezala, Ags. | Yes/Yes | 18 | 15 | 0.152 | 0 | 4.0 ± 0.2 |
| CSS16 | Tepezala, Ags. | Yes/Yes | 12 | ND | 0.102 | 0 | 5.0 ± 0.3 |
| CSS21 | Tepezala, Ags. | Yes/Yes | 21.5 | 13 | 0.150 | 0 | 2.6 ± 0.3 |
| CSS33 | El Llano, Ags. | Yes/Yes | 27.7 | 11 | 0.092 | 0 | 2.7 ± 0.3 |
| CSS34 | El Llano, Ags. | Yes/Yes | 25.6 | 10 | 0.177 | 0 | 5.8 ± 0.2 |
| CSS35 | Rio San Pedro, Ags. | Yes/Yes | 22.5 | 11 | 0.098 | 0 | 3.0 ± 0.4 |
| CAS259916 | Arizona, USA | Yes/Yes | 37.9 | 1 | 0.130 | 0 | 5.7 ± 0.7 |
| ID | Geographic Location | Genomic Mojave Toxin Presence (MTXA/MTXB) | % of MTX in Venom | % of SVMPs in Venom (RP-HPLC) | LD (mg/kg) | Hemorrhagic Activity (Intensity) | HPA Hydrolysis (U/mg) |
|---|---|---|---|---|---|---|---|
| CSS22 | Genaro García, Zacatecas | Yes/Yes | 17 | 30 | 0.136 | 4 | 29.2 ± 2.5 |
| CSS23 | Ojuelos, Jalisco | Yes/Yes | 29.5 | ND | ND | 2 | 25.5 ± 1.3 |
| CSS24 | Ojuelos, Jalisco | Yes/Yes | 29 | ND | ND | 3 | 39.8 ± 1.3 |
| CSS25 | Ojuelos, Jalisco | Yes/Yes | 21 | 23 | 0.203 | 2 | 21.8 ± 0.2 |
| CSS30 | La Asención, Nuevo León | Yes/Yes | 21.7 | 34 | 0.179 | 4 | 30.5 ± 2.3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borja, M.; Neri-Castro, E.; Castañeda-Gaytán, G.; Strickland, J.L.; Parkinson, C.L.; Castañeda-Gaytán, J.; Ponce-López, R.; Lomonte, B.; Olvera-Rodríguez, A.; Alagón, A.; et al. Biological and Proteolytic Variation in the Venom of Crotalus scutulatus scutulatus from Mexico. Toxins 2018, 10, 35. https://doi.org/10.3390/toxins10010035
Borja M, Neri-Castro E, Castañeda-Gaytán G, Strickland JL, Parkinson CL, Castañeda-Gaytán J, Ponce-López R, Lomonte B, Olvera-Rodríguez A, Alagón A, et al. Biological and Proteolytic Variation in the Venom of Crotalus scutulatus scutulatus from Mexico. Toxins. 2018; 10(1):35. https://doi.org/10.3390/toxins10010035
Chicago/Turabian StyleBorja, Miguel, Edgar Neri-Castro, Gamaliel Castañeda-Gaytán, Jason L. Strickland, Christopher L. Parkinson, Juan Castañeda-Gaytán, Roberto Ponce-López, Bruno Lomonte, Alejandro Olvera-Rodríguez, Alejandro Alagón, and et al. 2018. "Biological and Proteolytic Variation in the Venom of Crotalus scutulatus scutulatus from Mexico" Toxins 10, no. 1: 35. https://doi.org/10.3390/toxins10010035
APA StyleBorja, M., Neri-Castro, E., Castañeda-Gaytán, G., Strickland, J. L., Parkinson, C. L., Castañeda-Gaytán, J., Ponce-López, R., Lomonte, B., Olvera-Rodríguez, A., Alagón, A., & Pérez-Morales, R. (2018). Biological and Proteolytic Variation in the Venom of Crotalus scutulatus scutulatus from Mexico. Toxins, 10(1), 35. https://doi.org/10.3390/toxins10010035