The Impact of T-2 Toxin on Vasoactive Intestinal Polypeptide-Like Immunoreactive (VIP-LI) Nerve Structures in the Wall of the Porcine Stomach and Duodenum



Abstract
:1. Introduction
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lewis, L.; Onsongo, M.; Njapau, H.; Schurz-Rogers, H.; Luber, G.; Kieszak, S.; Nyamongo, J.; Backer, L.; Dahiye, A.M.; Misore, A.; et al. Kenya Aflatoxicosis Investigation Group. Aflatoxin contamination of commercial maize products during an outbreak of acute aflatoxicosis in eastern and central Kenya. Environ. Health Perspect. 2005, 113, 1763–1767. [Google Scholar] [CrossRef] [PubMed]
- De Ruyck, K.; De Boevre, M.; Huybrechts, I.; De Saeger, S. Dietary mycotoxins, co-exposure, and carcinogenesis in humans: Short review. Mutat. Res. Rev. Mutat. Res. 2015, 766, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Moretti, A.; Logrieco, A.F.; Susca, A. Mycotoxins: An underhand food problem. Methods Mol. Biol. 2017, 1542, 3–12. [Google Scholar] [PubMed]
- Makowska, K.; Obremski, K.; Zielonka, L.; Gonkowski, S. The Influence of Low Doses of Zearalenone and T-2 Toxin on Calcitonin Gene Related Peptide-Like Immunoreactive (CGRP-LI) Neurons in the ENS of the Porcine Descending Colon. Toxins 2017, 9, 98. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, M.J.; Macáková, P.; Juan-García, A.; Font, G. Cytotoxic effects of mycotoxin combinations in mammalian kidney cells. Food Chem. Toxicol. 2011, 49, 2718–2724. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, M.; Negi, B.; Kaushik, N.; Adhikari, A.; Al-Khedhairy, A.A.; Kaushik, N.K.; Choi, E.H. T-2 mycotoxin: Toxicological effects and decontamination strategies. Oncotarget 2017, 8, 33933–33952. [Google Scholar] [CrossRef] [PubMed]
- Makowska, K.; Gonkowski, S.; Zielonka, L.; Dabrowski, M.; Calka, J. T2 Toxin-Induced Changes in Cocaine- and Amphetamine-Regulated Transcript (CART)-Like Immunoreactivity in the Enteric Nervous System Within Selected Fragments of the Porcine Digestive Tract. Neurotox. Res. 2017, 31, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Weidner, M.; Huwel, S.; Ebert, F.; Schwerdtle, T.; Galla, H.J.; Humpf, H.U. Influence of T-2 and HT-2 toxin on the blood-brain barrier in vitro: New experimental hints for neurotoxic effects. PLoS ONE 2013, 8, e60484. [Google Scholar] [CrossRef] [PubMed]
- Lutsky, I.; Mor, N. Alimentary toxic aleukia (septic angina, endemic panmyelotoxicosis, alimentary hemorrhagic aleukia): T-2 toxin-induced intoxication of cats. Am. J. Pathol. 1981, 104, 189–191. [Google Scholar] [PubMed]
- Martin, L.J.; Doebler, J.A.; Anthony, A. Scanning cytophotometric analysis of brain neuronal nuclear chromatin changes in acute T-2 toxin-treated rats. Toxicol. Appl. Pharmacol. 1986, 85, 207–214. [Google Scholar] [CrossRef]
- Gonkowski, S.; Kamińska, B.; Landowski, P.; Całka, J. Immunohistochemical distribution of cocaine- and amphetamine-regulated transcript peptide—Like immunoreactive (CART-LI) nerve fibers and various degree of co-localization with other neuronal factors in the circular muscle layer of human descending colon. Histol. Histopathol. 2013, 28, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Makowska, K.; Gonkowski, S. The Influence of Inflammation and Nerve Damage on the Neurochemical Characterization of Calcitonin Gene-Related Peptide-Like Immunoreactive (CGRP-LI) Neurons in the Enteric Nervous System of the Porcine Descending Colon. Int. J. Mol. Sci. 2018, 19, 548. [Google Scholar] [CrossRef]
- Furness, J.B.; Callaghan, B.P.; Rivera, L.R.; Cho, H.J. The enteric nervous system and gastrointestinal innervation: Integrated local and central control. Adv. Exp. Med. Biol. 2014, 817, 39–71. [Google Scholar] [CrossRef] [PubMed]
- Makowska, K.; Gonkowski, S. Cocaine-And Amphetamine-Regulated Transcript (Cart) Peptide in Mammals Gastrointestinal System—A Review. Ann. Anim. Sci. 2017, 17, 3–21. [Google Scholar] [CrossRef]
- Furness, J.B. Types of neurons in the enteric nervous system. J. Autonom. Nerv. Syst. 2000, 81, 87–96. [Google Scholar] [CrossRef]
- Kasparek, M.S.; Fatima, J.; Iqbal, C.W.; Duenes, J.A.; Sarr, M.G. Role of VIP and Substance P in NANC innervation in the longitudinal smooth muscle of the rat jejunum -influence of extrinsic denervation. J. Surg. Res. 2007, 141, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Rytel, L.; Palus, K.; Calka, J. Co-expression of PACAP with VIP, SP and CGRP in the porcine nodose ganglion sensory neurons. Anat. Histol. Embryol. 2015, 44, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, M.; Morrison, K.J.; Vanhoutte, P.M. Hyperpolarization and relaxation of canine vascular smooth muscle to vasoactive intestinal polypeptide. J. Cardiovasc. Pharmacol. 1997, 30, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Van Geldre, L.A.; Lefebvre, R.A. Interaction of NO and VIP in gastrointestinal smooth muscle relaxation. Curr. Pharm. Des. 2004, 10, 2483–2497. [Google Scholar] [CrossRef] [PubMed]
- Nassar, C.F.; Abdallah, L.E.; Barada, K.A.; Atweh, S.F.; Saadé, N.F. Effects of intravenous vasoactive intestinal peptide injection on jejunal alanine absorption and gastric acid secretion in rats. Regul. Pept. 1995, 55, 261–267. [Google Scholar] [CrossRef]
- Burleigh, D.E.; Banks, M.R. Stimulation of intestinal secretion by vasoactive intestinal peptide and cholera toin. Auton. Neurosci. Basic Clin. 2007, 133, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Kaleczyc, J.; Klimczuk, M.; Franke-Radowiecka, A.; Sienkiewicz, W.; Majewski, M.; Łakomy, M. The distribution and chemical coding of intramural neurons supplying the porcine stomach—The study on normal pigs and on animals suffering from swine dysentery. Anat. Histol. Embryol. 2007, 36, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Arciszewski, M.B.; Sand, E.; Ekblad, E. Vasoactive intestinal peptide rescues cultured rat myenteric neurons from lipopolysaccharide induced cell death. Regul. Pept. 2008, 146, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Pidsudko, Z.; Kaleczyc, J.; Wasowicz, K.; Sienkiewicz, W.; Majewski, M.; Zajac, W.; Lakomy, M. Distribution and chemical coding of intramural neurons in the porcine ileum during proliferative enteropathy. J. Comp. Pathol. 2008, 138, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Gonkowski, S.; Obremski, K.; Calka, J. The Influence of Low Doses of Zearalenone on Distribution of Selected Active Substances in Nerve Fibers within the Circular Muscle Layer of Porcine Ileum. J. Mol. Neurosci. 2015, 56, 878–886. [Google Scholar] [CrossRef] [PubMed]
- Vasina, V.; Barbara, G.; Talamonti, L.; Stanghellini, V.; Corinaldesi, R.; Tonini, M.; de Ponti, F.; de Giorgio, R. Enteric neuroplasticity evoked by inflammation. Auton. Neurosci. 2006, 126–127, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Gonkowski, S.; Burlinski, P.; Calka, J. Proliferative enteropathy (PE)-induced changes in galanin-like immunoreactivity in the enteric nervous system of the porcine distal colon. Acta Vet. 2009, 59, 321–330. [Google Scholar]
- Gonkowski, S.; Burlinski, P.; Skobowiat, C.; Majewski, M.; Calka, J. Inflammation- And Axotomy-Induced Changes In Galanin-Like Immunoreactive (Gal-Li) Nerve Structures In The Porcine Descending Colon. Acta Vet. Hung. 2010, 58, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Gonkowski, S. Substance P as a neuronal factor in the enteric nervous system of the porcine descending colon in physiological conditions and during selected pathogenic processes. Biofactors 2013, 39, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Timmermans, J.P.; Adriaensen, D.; Cornelissen, W.; Scheuermann, D.W. Structural organization and neuropeptide distribution in the mammalian enteric nervous system, with special attention to those components involved in mucosal reflexes. Comp. Biochem. Physiol. A Physiol. 1997, 118, 331–340. [Google Scholar] [CrossRef]
- Gonkowski, S.; Wojtkiewicz, J.; Bossowska, A.; Kaleczyc, J.; Sienkiewicz, W.; Majewski, M. Proliferative enteropathy (PE)-induced changes in the number of vasoactive intestinal polypeptide-immunoreactive (VIP-IR) neural elements in the porcine descending colon. Pol. J. Vet. Sci. 2004, 7, 53–55. [Google Scholar]
- Verma, N.; Rettenmeier, A.W.; Schmitz-Spanke, S. Recent advances in the use of Sus scrofa (pig) as a model system for proteomic studies. Proteomics 2011, 11, 776–793. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.J.; Schemann, M.; Santer, R.M.; Cowen, T. The effects of age on the overall population and on sub-populations of myenteric neurons in the rat small intestine. J. Anat. 1998, 192, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Kleinschmidt, S.; Nolte, I.; Hewicker-Trautwein, M. Structural and functional changes of neuronal and glial components of the feline enteric nervous system in cats with chronic inflammatory and non-inflammatory diseases of the gastrointestinal tract. Res. Vet. Sci. 2011, 91, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Brenneman, D.E.; Philips, T.M.; Hauser, J.; Hill, J.M.; Spong, C.Y.; Gozes, I. Complex array of cytokines released by vasoactive intestinal peptide. Neuropeptides 2003, 37, 111–119. [Google Scholar] [CrossRef]
- Vota, D.; Aguero, M.; Grasso, E.; Hauk, V.; Gallino, L.; Soczewski, E.; Pérez Leirós, C.; Ramhorst, R. Progesterone and VIP cross-talk enhances phagocytosis and anti-inflammatory profile in trophoblast-derived cells. Mol. Cell Endocrinol. 2017, 443, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Obremski, K.; Gonkowski, S.; Wojtacha, P. Zearalenone-induced changes in the lymphoid tissue and mucosal nerve fibers in the porcine ileum. Pol. J. Vet. Sci. 2015, 18, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Bednarska, O.; Walter, S.A.; Casado-Bedmar, M.; Ström, M.; Salvo-Romero, E.; Vicario, M.; Mayer, E.A.; Keita, Å.V. Vasoactive Intestinal Polypeptide and Mast Cells Regulate Increased Passage of Colonic Bacteria in Patients With Irritable Bowel Syndrome. Gastroenterology 2017, 153, 948–960. [Google Scholar] [CrossRef] [PubMed]
- Kirchgessner, A.L.; Dodd, J.; Gershon, M.D. Markers shared between dorsal root and enteric ganglia. J. Comp. Neurol. 1988, 276, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Z.; Beier, R.C.; Shen, J.; De Smet, D.; De Saeger, S.; Zhang, S. T-2 toxin, a trichothecene mycotoxin: Review of toxicity, metabolism, and analytical methods. J. Agric. Food Chem. 2011, 59, 3441–3453. [Google Scholar] [CrossRef] [PubMed]
- Chang, I.M.; Mar, W.C. Effect of T-2 toxin on lipid peroxidation in rats: Elevation of conjugated diene formation. Toxicol. Lett. 1988, 40, 275–280. [Google Scholar] [PubMed]
- Shifrin, V.I.; Anderson, P. Trichothecene mycotoxins trigger a ribotoxic stress response that activates c-jun N-terminal kinase and p38 mitogen-activated protein kinase and induces apoptosis. J. Biol. Chem. 1999, 274, 13985–13992. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fitzpatrick, D.W.; Wilson, J.R. Effects of the trichothecene mycotoxin T-2 toxin on neurotransmitters and metabolites in discrete areas of the rat brain. Food Chem. Toxicol. 1998, 36, 947–953. [Google Scholar] [CrossRef]
- Sehata, S.; Kiyosawa, N.; Makino, T.; Atsumi, F.; Ito, K.; Yamoto, T.; Teranishi, M.; Baba, Y.; Uetauka, K.; Nakayama, H.; et al. Morphological and microarray analysis of T-2 toxin-induced rat fetal brain lesion. Food Chem. Toxicol. 2004, 42, 1727–1736. [Google Scholar] [CrossRef] [PubMed]
- Doi, K.; Uetsuka, K. Mechanisms of mycotoxin-induced neurotoxicity through oxidative stress-associated pathways. Int. J. Mol. Sci. 2011, 12, 5213–5237. [Google Scholar] [CrossRef] [PubMed]
- Brenneman, D.E.; Nicol, T.; Warren, D.; Bowers, L.M. Vasoactive intestinal peptide: A neurotrophic releasing agent and an astroglial mitogen. J. Neurosci. Res. 1990, 25, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Cazillis, M.; Gonzalez, B.J.; Billardon, C.; Lombet, A.; Fraichard, A.; Samarut, J.; Gressens, P.; Vaudry, H.; Rostène, W. VIP and PACAP induce selective neuronal differentiation of mouse embryonic stem cells. Eur. J. Neurosci. 2004, 19, 798–808. [Google Scholar] [CrossRef] [PubMed]
- Passemard, S.; Sokolowska, P.; Schwendimann, L.; Gressens, P. VIP-induced neuroprotection of the developing brain. Curr. Pharm. Des. 2011, 17, 1036–1039. [Google Scholar] [CrossRef] [PubMed]
- Rangon, C.M.; Dicou, E.; Goursaud, S.; Mounien, L.; Jégou, S.; Janet, T.; Muller, J.M.; Lelièvre, V.; Gressens, P. Mechanisms of VIP-induced neuroprotection against neonatal excitotoxicity. Ann. N. Y. Acad. Sci. 2006, 1070, 512–517. [Google Scholar] [CrossRef] [PubMed]
- Gressens, P.; Marret, S.; Hill, J.M.; Brenneman, D.E.; Gozes, I.; Fridkin, M.; Evrard, P. Vasoactive intestinal peptide prevents excitotoxic cell death in the murine developing brain. J. Clin. Investig. 1997, 100, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Carniglia, L.; Ramírez, D.; Durand, D.; Saba, J.; Turati, J.; Caruso, C.; Scimonelli, T.N.; Lasaga, M. Neuropeptides and Microglial Activation in Inflammation, Pain, and Neurodegenerative Diseases. Mediat. Inflamm. 2017, 2017, 5048616. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekharan, B.; Nezami, B.G.; Srinivasan, S. Emerging neuropeptide targets in inflammation: NPY and VIP. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G949–G957. [Google Scholar] [CrossRef] [PubMed]
- Sandgren, K.; Lin, Z.; Ekblad, E. Differential effects of VIP and PACAP on survival of cultured adult rat myenteric neurons. Regul. Pept. 2003, 111, 211–217. [Google Scholar] [CrossRef]
- Goossens, J.; Pasmans, F.; Verbrugghe, E.; Vandenbroucke, V.; De Baere, S.; Meyer, E.; Haesebrouck, F.; De Backer, P.; Croubels, S. Porcine intestinal epithelial barrier disruption by the Fusarium mycotoxins deoxynivalenol and T-2 toxin promotes transepithelial passage of doxycycline and paromomycin. BMC Vet. Res. 2012, 8, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obremski, K.; Zielonka, L.; Gajecka, M.; Jakimiuk, E.; Bakuła, T.; Baranowski, M.; Gajecki, M. Histological estimation of the small intestine wall after administration of feed containing deoxynivalenol, T-2 toxin and zearalenone in the pig. Pol. J. Vet. Sci. 2008, 11, 339–345. [Google Scholar] [PubMed]
- Moriez, R.; Abdo, H.; Chaumette, T.; Faure, M.; Lardeux, B.; Neunlist, M. Neuroplasticity and neuroprotection in enteric neurons: Role of epithelial cells. Biochem. Biophys. Res. Commun. 2009, 382, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Ganea, D.; Hooper, K.M.; Kong, W. The neuropeptide vasoactive intestinal peptide: Direct effects on immune cells and involvement in inflammatory and autoimmune diseases. Acta Physiol 2015, 213, 442–452. [Google Scholar] [CrossRef] [PubMed]
- Delgado, M.; Munoz-Elias, E.J.; Gomariz, R.P.; Ganea, D. Vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide enhance IL-10 production by murine macrophages: In vitro and in vivo studies. J. Immunol. 1999, 162, 1707–1716. [Google Scholar] [PubMed]
- Delgado, M.; Pozo, D.; Ganea, D. The significance of vasoactive intestinal peptide in immunomodulation. Pharmacol. Rev. 2004, 56, 249–290. [Google Scholar] [CrossRef] [PubMed]
- Obremski, K.; Podlasz, P.; Zmigrodzka, M.; Winnicka, A.; Woźny, M.; Brzuzan, P.; Jakimiuk, E.; Wojtacha, P.; Gajecka, M.; Zielonka, Ł.; et al. The effect of T-2 toxin on percentages of CD4+, CD8+, CD4+ CD8+ and CD21+ lymphocytes, and mRNA expression levels of selected cytokines in porcine ileal Peyer’s patches. Pol. J. Vet. Sci. 2013, 16, 341–349. [Google Scholar] [CrossRef] [PubMed]





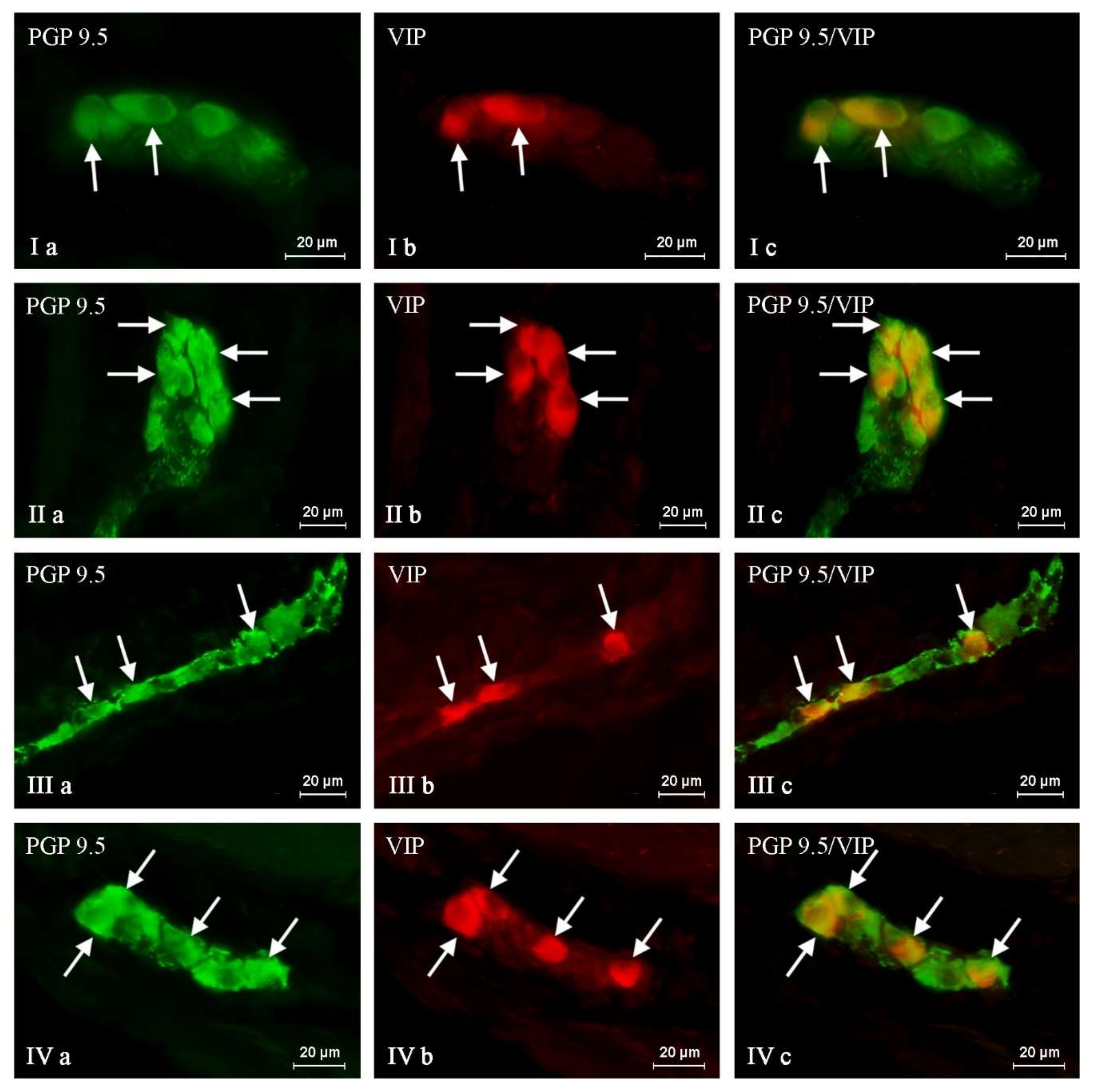{kind=link}
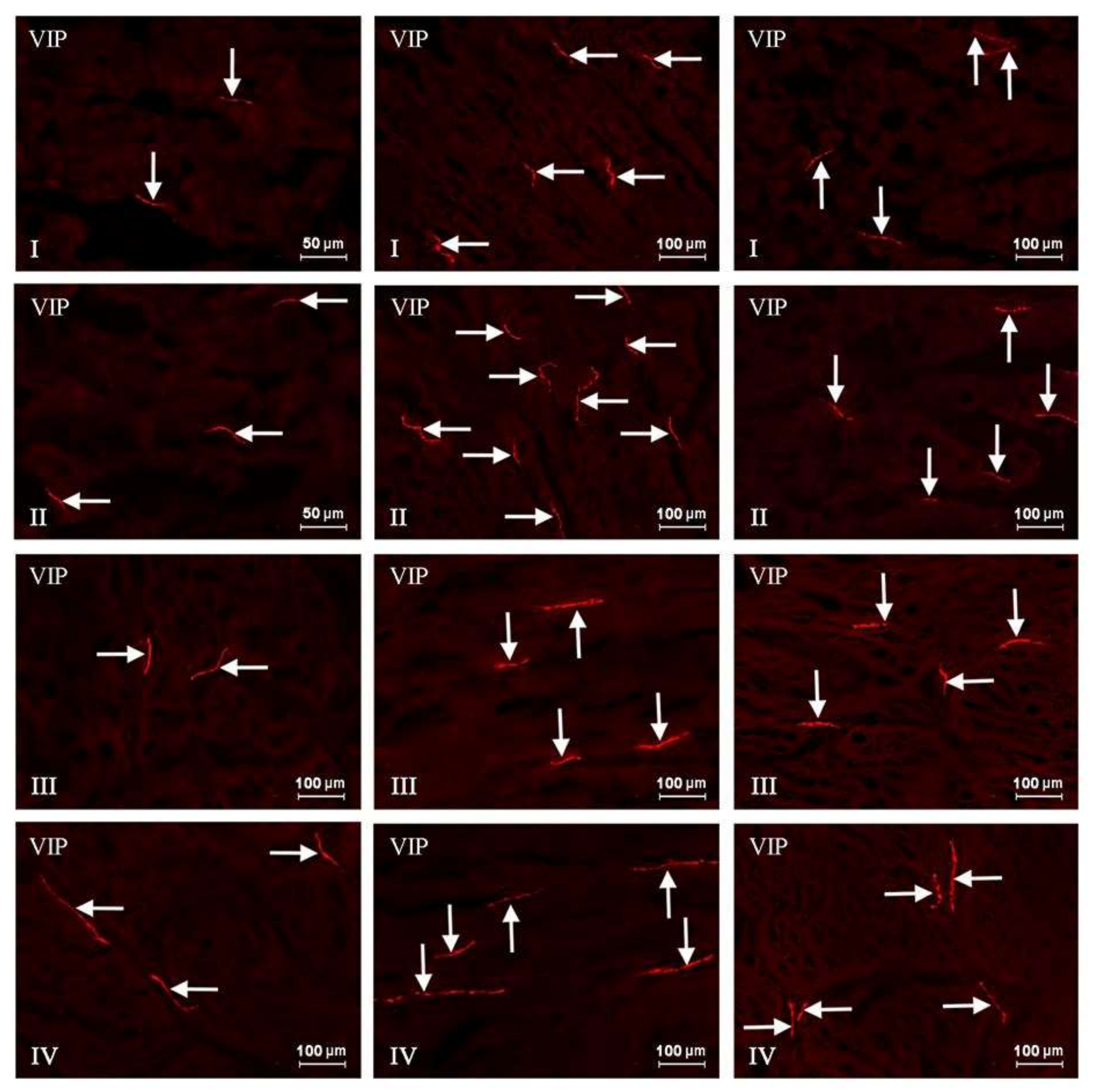{kind=link}
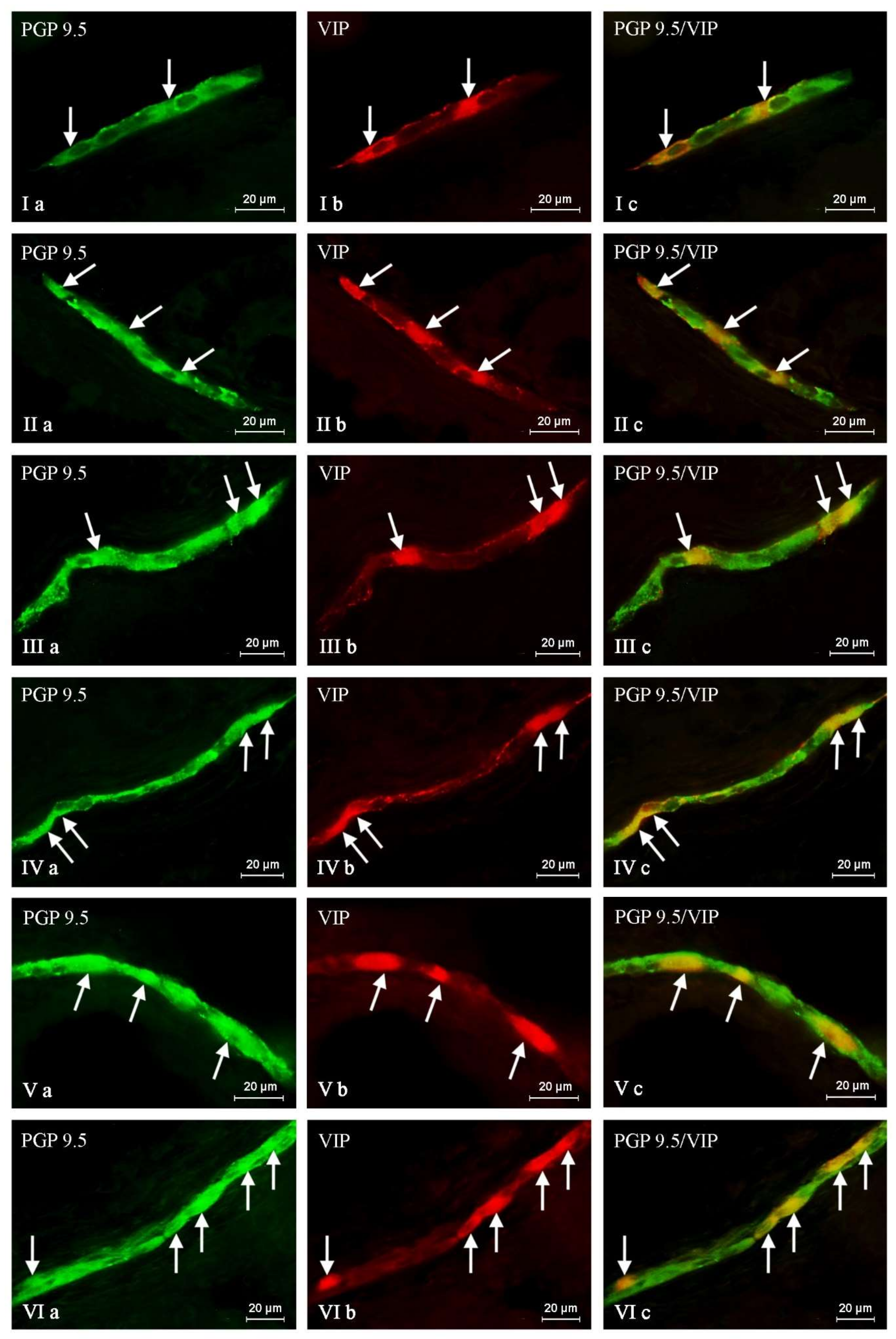{kind=link}
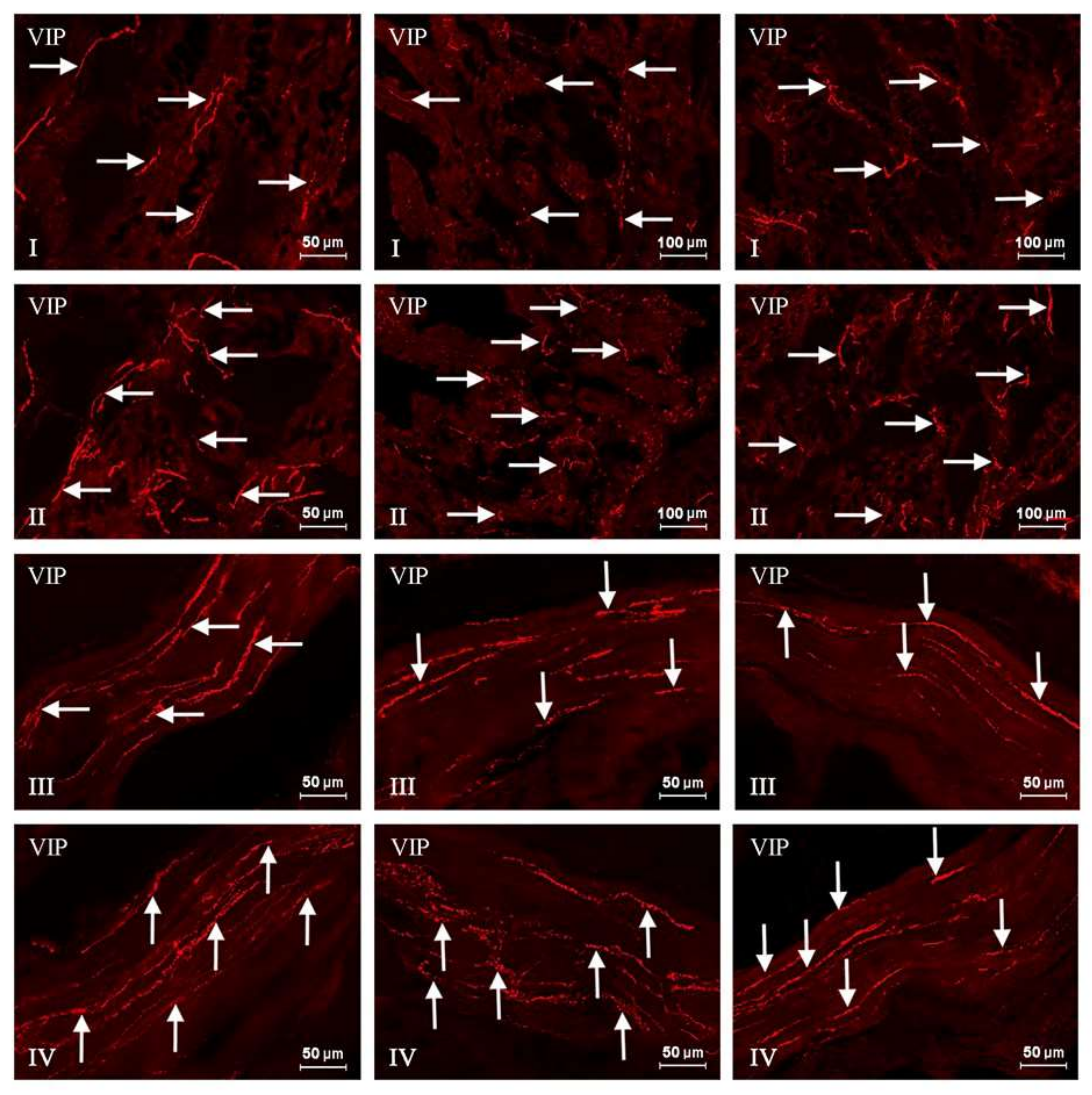{kind=link}
| STOMACH | Group C | Group T2 | |
|---|---|---|---|
| CML 1 | 4.26 ± 0.21 | 5.17 ± 0.11 * | |
| MP | CB 2 | 37.56± 0.84 | 42.85 ± 0.74 * |
| NF 3 | + | + | |
| SP | CB 2 | 36.78 ± 0.4 | 43.83 ± 1.18 * |
| NF 3 | − | + | |
| S/ML 1 | 2.84 ± 0.07 | 4.15 ± 0.15 * | |
| DUDODENUM | Group C | Group T2 | |
|---|---|---|---|
| CML 1 | 17.08 ± 0.08 | 21.22 ± 0.24 * | |
| MP | CB 2 | 31.45 ± 0.77 | 39.24 ± 1.02 * |
| NF 3 | + | ++ | |
| OSP | CB 2 | 32.43 ± 1.83 | 40.59 ± 0.67 * |
| NF 3 | ++ | ++ | |
| ISP | CB 2 | 28.50 ± 1.17 | 35.42 ± 1.52 * |
| NF 3 | + | ++ | |
| S/ML 1 | 32.35 ± 0.32 | 39.97 ± 1.23 * | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Makowska, K.; Obremski, K.; Gonkowski, S. The Impact of T-2 Toxin on Vasoactive Intestinal Polypeptide-Like Immunoreactive (VIP-LI) Nerve Structures in the Wall of the Porcine Stomach and Duodenum. Toxins 2018, 10, 138. https://doi.org/10.3390/toxins10040138
Makowska K, Obremski K, Gonkowski S. The Impact of T-2 Toxin on Vasoactive Intestinal Polypeptide-Like Immunoreactive (VIP-LI) Nerve Structures in the Wall of the Porcine Stomach and Duodenum. Toxins. 2018; 10(4):138. https://doi.org/10.3390/toxins10040138
Chicago/Turabian StyleMakowska, Krystyna, Kazimierz Obremski, and Slawomir Gonkowski. 2018. "The Impact of T-2 Toxin on Vasoactive Intestinal Polypeptide-Like Immunoreactive (VIP-LI) Nerve Structures in the Wall of the Porcine Stomach and Duodenum" Toxins 10, no. 4: 138. https://doi.org/10.3390/toxins10040138
APA StyleMakowska, K., Obremski, K., & Gonkowski, S. (2018). The Impact of T-2 Toxin on Vasoactive Intestinal Polypeptide-Like Immunoreactive (VIP-LI) Nerve Structures in the Wall of the Porcine Stomach and Duodenum. Toxins, 10(4), 138. https://doi.org/10.3390/toxins10040138