Leukotoxin of Bibersteinia trehalosi Contains a Unique Neutralizing Epitope, and a Non-Neutralizing Epitope Shared with Mannheimia haemolytica Leukotoxin

Abstract
:1. Introduction
2. Results and Discussion
2.1. Monoclonal Antibody AM113 Reacts Only with B. trehalosi Lkt While AM 321 Reacts with Lkts of B. trehalosi and M. haemolytica
2.2. Western Blot Analysis Confirms the Specificity of mAbs AM113 and AM321
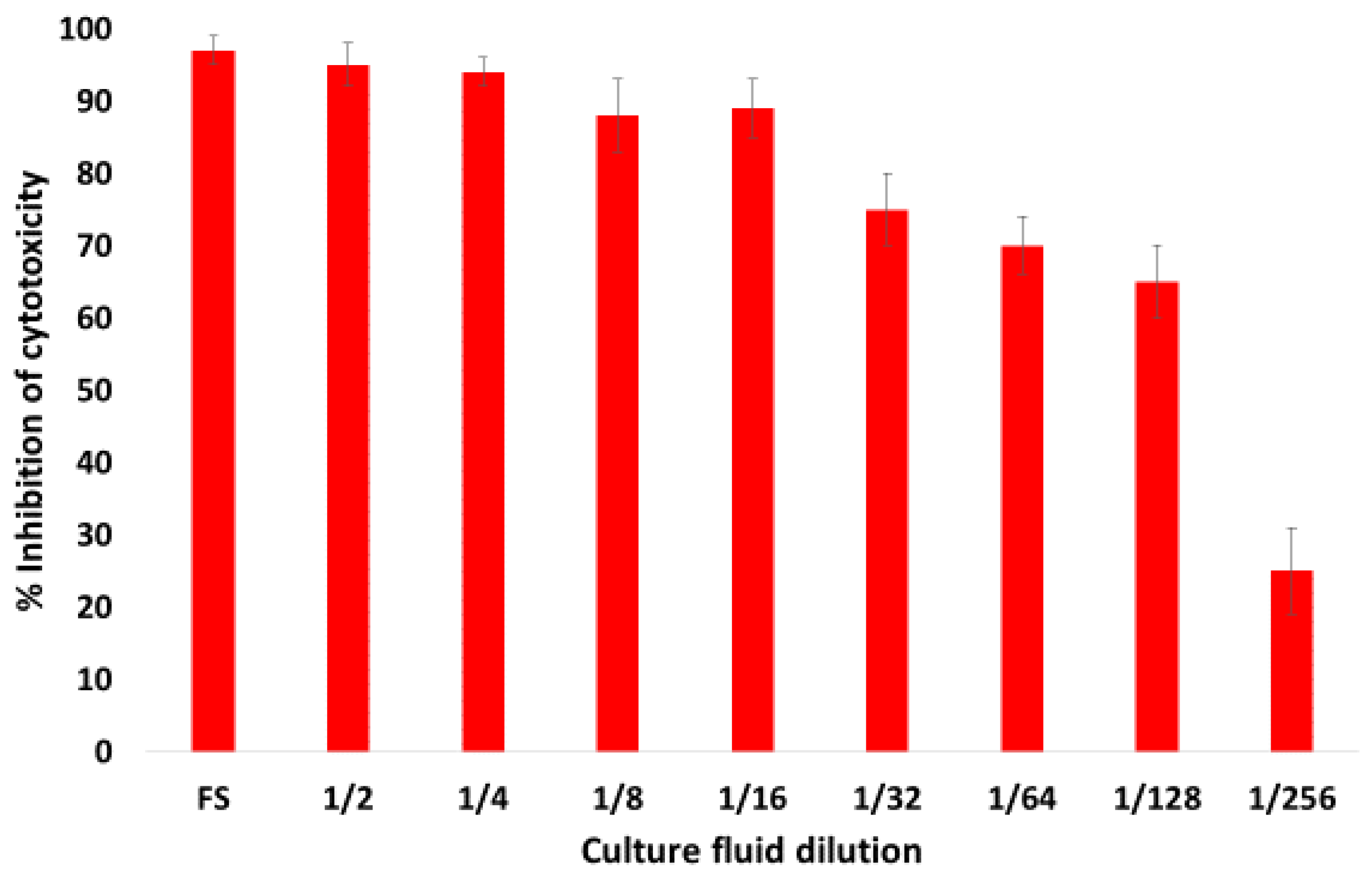
2.3. Monoclonal Antibody AM113 Neutralizes B. trehalosi Lkt While mAb AM321 Does Not
2.4. Monoclonal Antibodies Reveal the Presence of Unique Neutralizing Epitopes on the Lkts of B. trehalosi and M. haemolytica Serotype A1
3. Conclusions
- Lkts of B. trehalosi and M. haemolytica serotype A1 carry neutralizing epitopes uniquely present on them.
- Lkts of B. trehalosi, M. haemolytica serotype A1, and serotype A2 carry a shared non-neutralizing epitope.
- The mAb developed in this study should pave the way for mapping of a neutralizing epitope on B. trehalosi Lkt, which in turn should help in the development of subunit vaccines and virus-vectored live vaccines against this economically important respiratory pathogen of domestic and wild ruminants.
4. Materials and Methods
4.1. Production of Lkt
4.2. Immunization of Mice and Development of Hybridomas
4.3. ELISA
4.4. Western Blot Assay
4.5. Cytotoxicity Assay for Detection of Cytotoxicity of Lkt Preparations and Cytotoxicity Inhibition Assay for Detection of Lkt-Neutralizing Activity of mAbs
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Mosier, D.A. Bacterial pneumonia. Vet. Clin. N. Am Food Anim. Pract. 1997, 13, 483–493. [Google Scholar] [CrossRef]
- Miller, M.W. Pasteurellosis. In Infectious Diseases of Wild Mammals; Iowa State University Press: Ames, IA, USA, 2008; pp. 330–339. [Google Scholar]
- Ackermann, M.R.; Brogden, K.A. Response of the ruminant respiratory tract to Mannheimia (Pasteurella) haemolytica. Microbes Infect. 2000, 2, 1079–1088. [Google Scholar] [CrossRef]
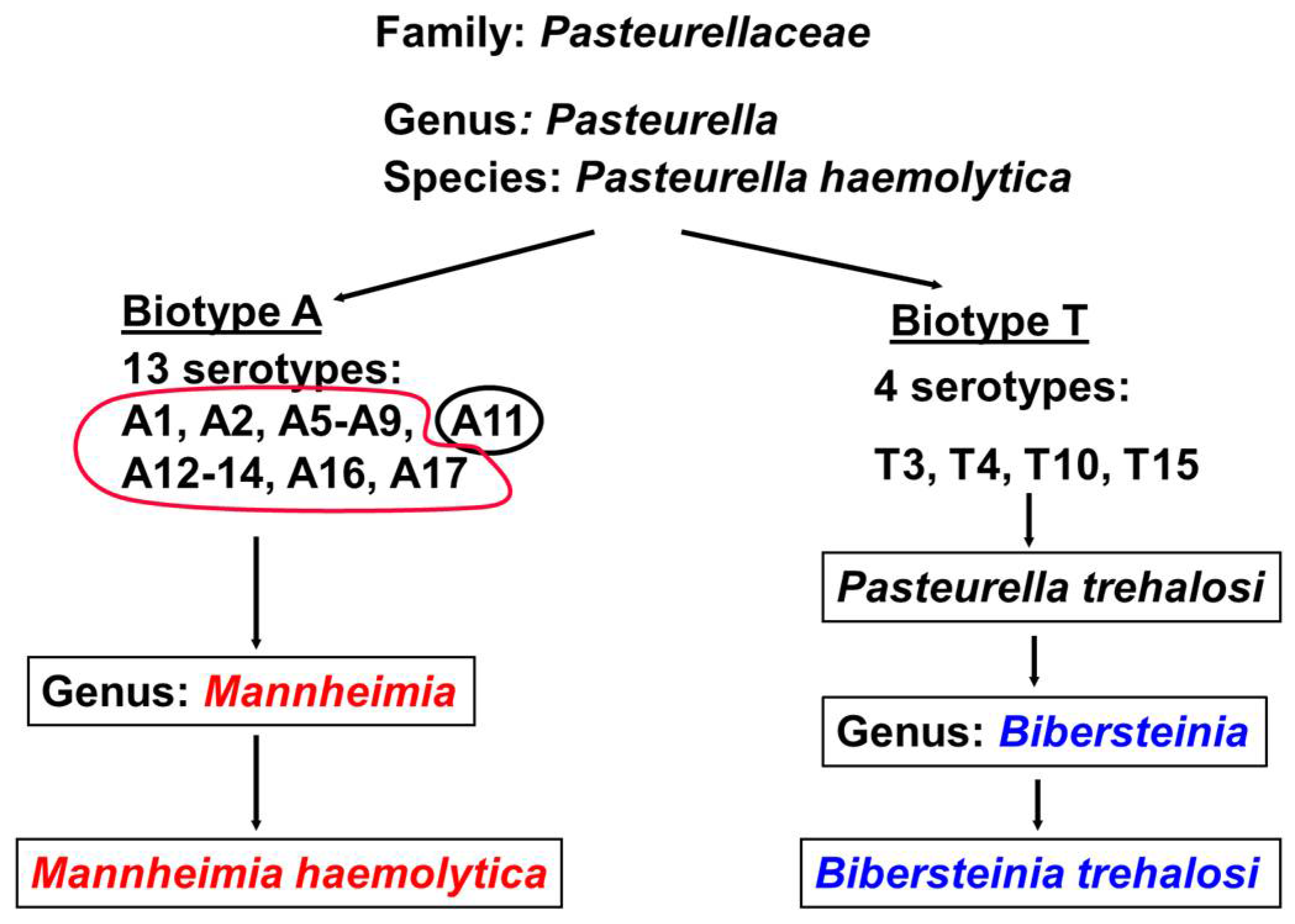
- Angen, O.; Mutters, R.; Caugant, D.A.; Olsen, J.E.; Bisgaard, M. Taxonomic relationships of the [Pasteurella] haemolytica complex as evaluated by DNA-DNA hybridizations and 16S rRNA sequencing with proposal of Mannheimia haemolytica gen. nov., comb. nov., Mannheimia granulomatis comb. nov., Mannheimia glucosida sp. nov., Mannheimia ruminalis sp. nov. and Mannheimia varigena sp. nov. Int. J. Syst. Bacteriol. 1999, 49 Pt 1, 67–86. [Google Scholar] [PubMed]
- Blackall, P.J.; Bojesen, A.M.; Christensen, H.; Bisgaard, M. Reclassification of [Pasteurella] trehalosi as Bibersteinia trehalosi gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2007, 57 Pt 4, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Dassanayake, R.P.; Shanthalingam, S.; Herndon, C.N.; Subramaniam, R.; Lawrence, P.K.; Bavananthasivam, J.; Haldorson, G.J.; Evermann, J.; Foreyt, W.J.; Knowles, D.P.; et al. Role of Bibersteinia trehalosi, respiratory syncytial virus and parainfluenza 3 virus in pneumonia in bighorn sheep. Vet. Microbiol. 2013, 162, 166–172. [Google Scholar] [CrossRef] [PubMed]
- British Veterinary Association. Northern Ireland disease surveillance report, October to December 2015. Vet. Rec. 2016, 178, 162–165. [Google Scholar]
- Bowersock, T.L.; Sobecki, B.E.; Terrill, S.J.; Martinon, N.C.; Meinert, T.R.; Leyh, R.D. Efficacy of a multivalent modified-live virus vaccine containing a Mannheimia haemolytica toxoid in calves challenge-exposed with Bibersteinia trehalosi. Am. J. Vet. Res. 2014, 75, 770–776. [Google Scholar] [CrossRef] [PubMed]
- Rehmtulla, A.J.; Thomson, R.G. A review of the lesions in shipping fever of cattle. Can. J. Vet. 1981, 22, 1–8. [Google Scholar]
- Confer, A.W.; Panciera, R.J.; Clinkenbeard, K.D.; Mosier, D.A. Molecular aspects of virulence of Pasteurella haemolytica. Can. J. Vet. Res. 1990, 54, S48–S52. [Google Scholar] [PubMed]
- Dassanayake, R.P.; Shanthalingam, S.; Herndon, C.N.; Lawrence, P.K.; Frances Cassirer, F.E.; Potter, K.A.; Foreyt, W.J.; Clinkenbeard, K.D.; Srikumaran, S. Mannheimia haemolytica serotype A1 exhibits differential pathogenicity in two related species, Ovis canadensis and Ovis aries. Vet. Microbiol. 2009, 133, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Petras, S.F.; Chidambaram, M.; Illyes, E.F.; Froshauer, S.; Weinstock, G.M.; Reese, C.P. Antigenic and virulence properties of Pasteurella haemolytica leukotoxin mutants. Infect. Immun. 1995, 63, 1033–1039. [Google Scholar] [PubMed]
- Highlander, S.K.; Fedorova, N.D.; Dusek, D.M.; Panciera, R.; Alvarez, L.E.; Rinehart, C. Inactivation of Pasteurella (Mannheimia) haemolytica leukotoxin causes partial attenuation of virulence in a calf challenge model. Infect. Immun. 2000, 68, 3916–3922. [Google Scholar] [CrossRef] [PubMed]
- Strathdee, C.A.; Lo, R.Y. Cloning, nucleotide sequence, and characterization of genes encoding the secretion function of the Pasteurella haemolytica leukotoxin determinant. J. Bacteriol. 1989, 171, 916–928. [Google Scholar] [CrossRef] [PubMed]
- Devenish, J.; Rosendal, S.; Johnson, R.; Hubler, S. Immunoserological comparison of 104-kilodalton proteins associated with hemolysis and cytolysis in Actinobacillus pleuropneumoniae, Actinobacillus suis, Pasteurella haemolytica, and Escherichia coli. Infect. Immun. 1989, 57, 3210–3213. [Google Scholar] [PubMed]
- Kolodrubetz, D.; Dailey, T.; Ebersole, J.; Kraig, E. Cloning and expression of the leukotoxin gene from Actinobacillus actinomycetemcomitans. Infect. Immun. 1989, 57, 1465–1469. [Google Scholar] [PubMed]
- Kaehler, K.L.; Markham, R.J.; Muscoplat, C.C.; Johnson, D.W. Evidence of species specificity in the cytocidal effects of Pasteurella haemolytica. Infect. Immun. 1980, 30, 615–616. [Google Scholar] [PubMed]
- Chang, Y.F.; Renshaw, H.W.; Martens, R.J.; Livingston, C.W., Jr. Pasteurella haemolytica leukotoxin: Chemiluminescent responses of peripheral blood leukocytes from several different mammalian species to leukotoxin- and opsonin-treated living and killed Pasteurella haemolytica and Staphylococcus aureus. Am. J. Vet. Res. 1986, 47, 67–74. [Google Scholar] [PubMed]
- Shewen, P.E.; Wilkie, B.N. Cytotoxin of Pasteurella haemolytica acting on bovine leukocytes. Infect. Immun. 1982, 35, 91–94. [Google Scholar] [PubMed]
- Slocombe, R.J.; Marark, J.; Ingersoll, R.; Derksen, F.J.; Robinson, N.E. Importance of neutrophils in the pathogenesis of acute pneumonic pasteurellosis in calves. Am. J. Vet. Res. 1985, 46, 2253–2258. [Google Scholar] [PubMed]
- Highlander, S.K. Molecular genetic analysis of virulence in Mannheimia (Pasteurella) haemolytica. Front. Biosci. 2001, 1, D1128–D1150. [Google Scholar]
- Jayaseelan, S.; Sreevatsan, S.; Maheswaran, S.K. Role of Mannheimia haemolytica leukotoxin in the pathogenesis of bovine pneumonic pasteurellosis. Anim. Health Res. Rev. 2002, 3, 69–82. [Google Scholar] [CrossRef]
- Hodgins, D.C.; Shewen, P.E. Vaccination of neonatal colostrum-deprived calves against Pasteurella haemolytica A1. Can. J. Vet. Res. 2000, 64, 3–8. [Google Scholar] [PubMed]
- Orouji, S.; Hodgins, D.C.; Lo, R.Y.C.; Shewen, P.E. Serum IgG response in calves to the putative pneumonic virulence factor Gs60 of Mannheimia haemolytica A1. Can. J. Vet. Res. 2012, 76, 292–300. [Google Scholar] [PubMed]
- Sreevatsan, S.; Ames, T.R.; Werdin, R.E.; Yoo, H.S.; Maheswaran, S.K. Evaluation of three experimental subunit vaccines against pneumonic pasteurellosis in cattle. Vaccine 1996, 14, 147–154. [Google Scholar] [CrossRef]
- Subramaniam, R.; Shanthalingam, S.; Bavananthasivam, J.; Kugadas, A.; Potter, K.A.; Foreyt, W.J.; Hodgins, D.C.; Shewen, P.; Barrington, G.M.; Knowles, D.P.; et al. A multivalent Mannheimia/Bibersteinia vaccine protects bighorn sheep against Mannheimia haemolytica challenge. Clin. Vaccine Immunol. 2011, 18, 1689–1694. [Google Scholar] [CrossRef] [PubMed]
- Gentry, M.J.; Srikumaran, S. Neutralizing monoclonal antibodies to Pasteurella haemolytica leukotoxin affinity-purify the toxin from crude culture supernatants. Microb. Pathog. 1991, 10, 411–417. [Google Scholar] [CrossRef]
- Gerbig, D.G.; Cameron, M.R.; Struck, D.K.; Moore, R.N. Characterization of a Neutralizing Monoclonal Antibody to Pasteurella haemolytica Leukotoxin. Infect. Immun. 1992, 60, 1734–1739. [Google Scholar] [PubMed]
- Shanthalingam, S.; Srikumaran, S. Intact signal peptide of CD18, the beta- subunit of beta2-integrins, renders ruminants susceptible to Mannheimia haemolytica leukotoxin. Proc. Natl. Acad. Sci. USA 2009, 106, 15448–15453. [Google Scholar] [CrossRef] [PubMed]
- Shewen, P.E.; Wilkie, B.N. Pasteurella haemolytica cytotoxin: Production by recognized serotypes and neutralization by type-specific rabbit antisera. Am. J. Vet. Res. 1983, 44, 715–719. [Google Scholar] [PubMed]
- Lainson, F.; Murray, J.; Davies, R.; Donachie, W. Characterization of epitopes involved in the neutralization of Pasteurella haemolytica serotype A1 leukotoxin. Microbiology 1996, 142, 2499–2507. [Google Scholar] [CrossRef] [PubMed]
- Confer, A.; Ayalew, S.; Montelongo, M.; Step, D.; Wray, J.; Hansen, R.; Panciera, R. Immunity of cattle following vaccination with a Mannheimia haemolytica chimeric PlpE–LKT (SAC89) protein. Vaccine 2009, 27, 1771–1776. [Google Scholar] [CrossRef] [PubMed]
- Batra, S.A.; Shanthalingam, S.; Donofrio, G.; Srikumaran, S. A chimeric protein comprising the immunogenic domains of Mannheimia haemolytica leukotoxin and outer membrane protein PlpE induces antibodies against leukotoxin and PlpE. Vet. Immunol. Immunopathol. 2016, 175, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Draper, S.J.; Heeney, J.L. Viruses as vaccine vectors for infectious diseases and cancer. Nat. Rev. Microbiol. 2010, 8, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Batra, S.A.; Shanthalingam, S.; Donofrio, G.; Chowdhury, S.; Haldorson, G.J.; White, S.N.; Srikumaran, S. Immunization of bighorn sheep against Mannheimia haemolytica with a bovine herpesvirus 1-vectored vaccine. Vaccine 2016, 35, 1630–1636. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Monoclonal Antibody | Leukotoxin Neutralization | ||
|---|---|---|---|
| B. trehalosi | M. haemolytica A1 | M. haemolytica A2 | |
| AM113 | + | - | - |
| MM601 | - | + | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murugananthan, A.; Shanthalingam, S.; Batra, S.A.; Alahan, S.; Srikumaran, S. Leukotoxin of Bibersteinia trehalosi Contains a Unique Neutralizing Epitope, and a Non-Neutralizing Epitope Shared with Mannheimia haemolytica Leukotoxin. Toxins 2018, 10, 220. https://doi.org/10.3390/toxins10060220
Murugananthan A, Shanthalingam S, Batra SA, Alahan S, Srikumaran S. Leukotoxin of Bibersteinia trehalosi Contains a Unique Neutralizing Epitope, and a Non-Neutralizing Epitope Shared with Mannheimia haemolytica Leukotoxin. Toxins. 2018; 10(6):220. https://doi.org/10.3390/toxins10060220
Chicago/Turabian StyleMurugananthan, Arumugam, Sudarvili Shanthalingam, Sai Arun Batra, Sitara Alahan, and Subramaniam Srikumaran. 2018. "Leukotoxin of Bibersteinia trehalosi Contains a Unique Neutralizing Epitope, and a Non-Neutralizing Epitope Shared with Mannheimia haemolytica Leukotoxin" Toxins 10, no. 6: 220. https://doi.org/10.3390/toxins10060220
APA StyleMurugananthan, A., Shanthalingam, S., Batra, S. A., Alahan, S., & Srikumaran, S. (2018). Leukotoxin of Bibersteinia trehalosi Contains a Unique Neutralizing Epitope, and a Non-Neutralizing Epitope Shared with Mannheimia haemolytica Leukotoxin. Toxins, 10(6), 220. https://doi.org/10.3390/toxins10060220