Fructo-Oligosaccharide (DFA III) Feed Supplementation for Mitigation of Mycotoxin Exposure in Cattle—Clinical Evaluation by a Urinary Zearalenone Monitoring System

 ,
,
Abstract
:1. Introduction
2. Results
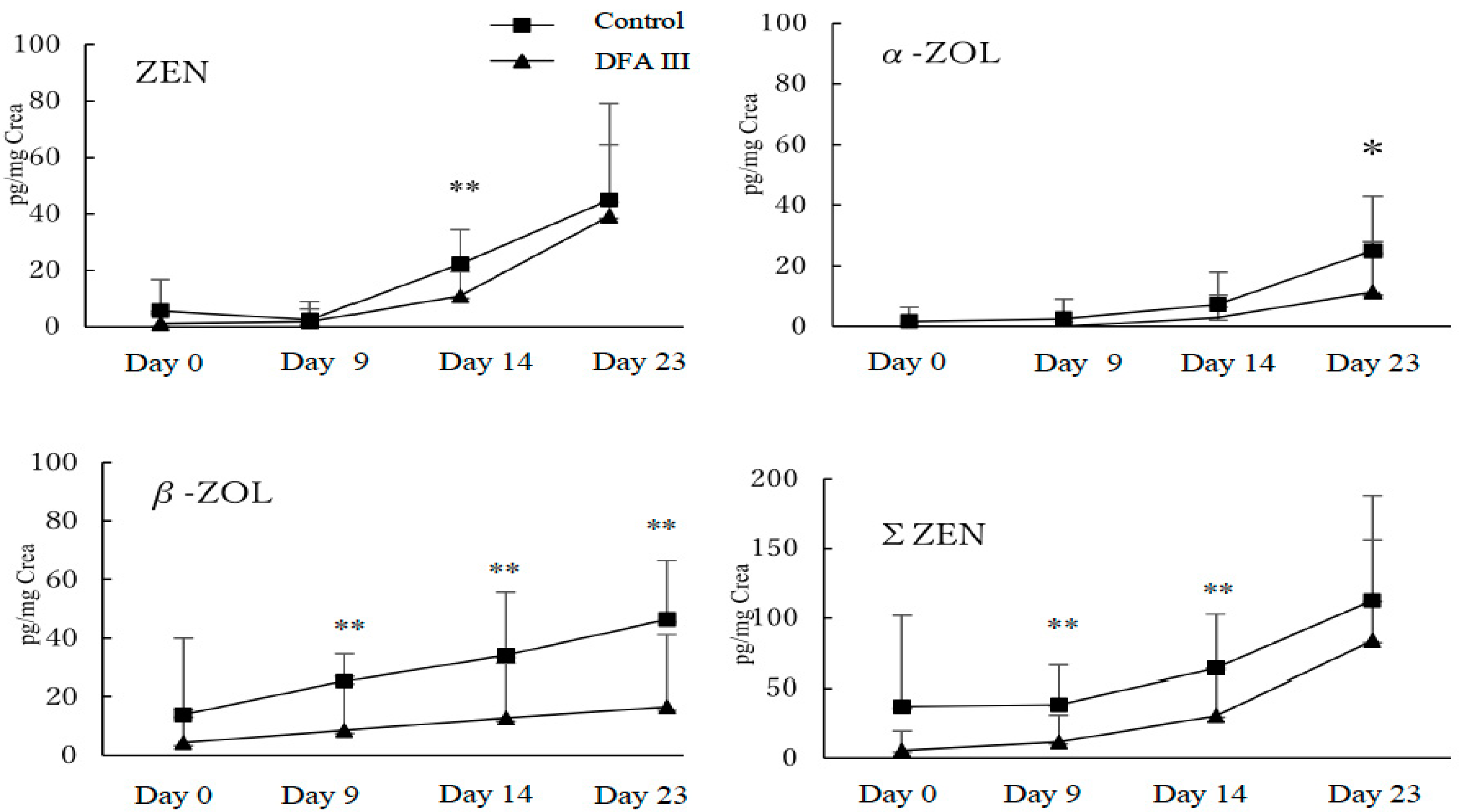
2.1. Concentration of ZEN and Its Metabolites with or without Supplementation of DFA III
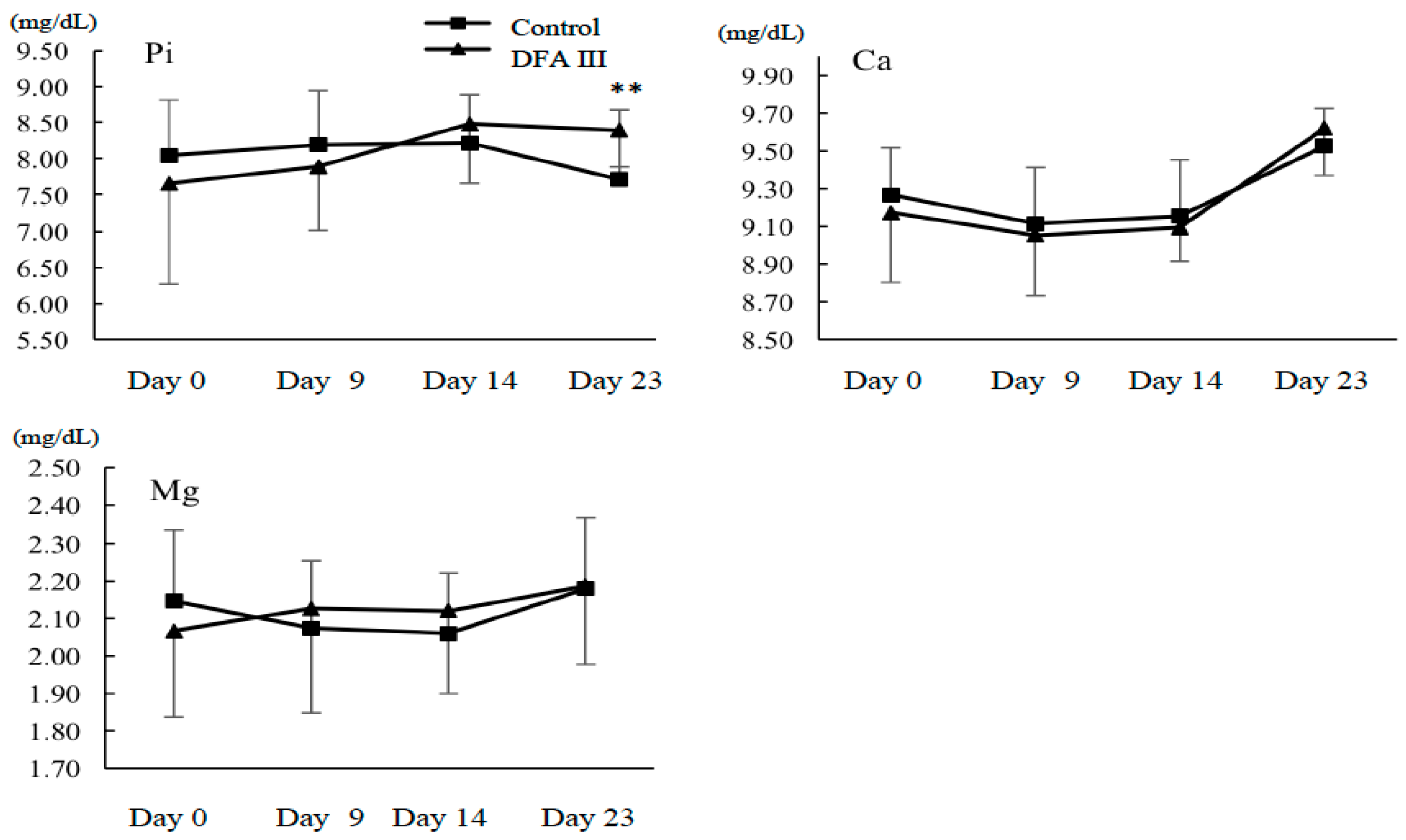
2.2. Ca, Mg, and Pi Concentrations with or without Supplementation of DFA III
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Solvents
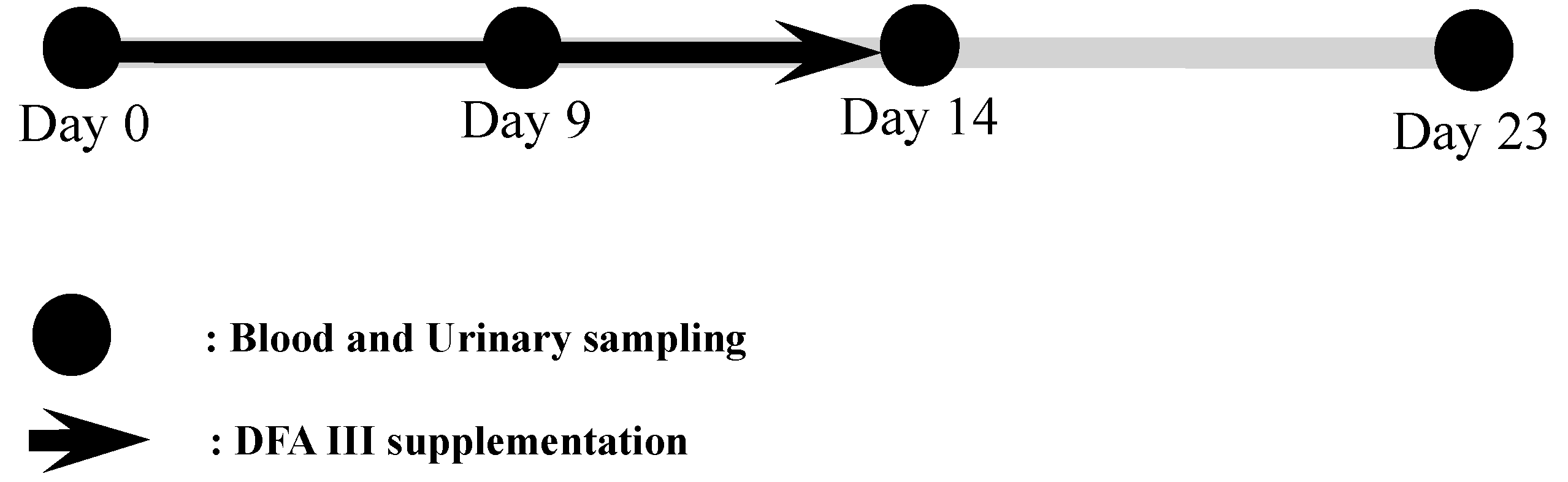
5.2. Japanese Black Cattle Herds and Sample Collection
5.3. Methods of Urine Sample Analysis
5.4. Statistical analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fink-Gremmels, J. The role of mycotoxins in the health and performance of dairy cows. Vet. J. 2008, 176, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Yan, T.; Wang, J.; Huang, Z.; Chen, X.; Jia, G.; Wu, C.; Zhao, H.; Xue, B.; Xiao, L.; et al. Biological system responses to zearalenone mycotoxin exposure by integrated metabolomic studies. J. Agric. Food Chem. 2013, 61, 11212–11221. [Google Scholar] [CrossRef] [PubMed]
- Hasunuma, H.; Takagi, M.; Kawamura, O.; Taniguchi, C.; Nakamura, M.; Chuma, T.; Uno, S.; Kokushi, E.; Matsumoto, D.; Tshering, C.; et al. Natural contamination of dietary rice straw with zearalenone and urinary zearalenone concentrations in a cattle herd. J. Anim. Sci. 2012, 90, 1610–1616. [Google Scholar] [CrossRef] [PubMed]
- Takagi, M.; Uno, S.; Kokushi, E.; Shiga, S.; Mukai, S.; Kuriyagawa, T.; Takagaki, K.; Hasunuma, H.; Matsumoto, D.; Okamoto, K.; et al. Measurement of urinary zearalenone concentrations for monitoring natural feed contamination in cattle herds-on farm trials. J. Anim. Sci. 2011, 89, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Sabater-Vilar, M.; Malekinejad, H.; Selman, M.H.J.; van der Doelen, M.A.M.; Fink-Gremmels, J. In vitro assessment of adsorbents aiming to prevent deoxynivalenol and zearalenone mycotoxicoses. Mycopathologia 2007, 163, 81–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutz, R.E.; Sampson, J.D.; Pompeu, L.B.; Ledoux, D.R.; Spain, J.N.; Vazquez-Añon, M.; Rottinghaus, G.E. Efficacy of Solis, NovasilPlus, and MTB-100 to reduce aflatoxin M1 levels in milk of early to mid-lactation dairy cows fed aflatoxin B1. J. Dairy Sci. 2009, 92, 3959–3963. [Google Scholar] [CrossRef] [PubMed]
- Awad, W.A.; Ghareeb, K.; Bohm, J.; Zentek, J. Decontamination and detoxification strategies for the Fusarium mycotoxin deoxynivalenol in animal feed and the effectiveness of microbial biodegradation. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2010, 27, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Wambacq, E.; Vanhoutte, I.; Audenaert, K.; De Gelder, L.; Haesaert, G. Occurrence, prevention and remediation of toxigenic fungi and mycotoxins in silage: A review. J. Sci. Food Agric. 2016, 96, 2284–2302. [Google Scholar] [CrossRef] [PubMed]
- Fushimi, Y.; Takagi, M.; Hasunuma, H.; Uno, S.; Kokushi, E.; Watanabe, U.; Liu, J.; Marey, M.; Miyamoto, A.; Otoi, T.; et al. Application of mycotoxin adsorbent to cattle feed naturally contaminated with zearalenone: Urinary zearalenone excretion and association with anti-Müllerian hormone. World Mycotoxin J. 2014, 7, 367–378. [Google Scholar] [CrossRef]
- Heinrichs, A.J.; Jones, C.M.; Heinrichs, B.S. Effects of mannan oligosaccharide or antibiotics in neonatal diets on health and growth of dairy calves. J. Dairy Sci. 2003, 86, 4064–4069. [Google Scholar] [CrossRef]
- Franklin, S.T.; Newman, M.C.; Newman, K.E.; Meek, K.I. Immune parameters of dry cows fed mannanoligosaccharide and subsequent transfer of immunity to calves. J. Dairy Sci. 2005, 88, 766–775. [Google Scholar] [CrossRef]
- Donovan, D.C.; Franklin, S.T.; Chase, C.C.; Hippen, A.R. Growth and health of Holstein calves fed milk replacers supplemented with antibiotics or enteroguard. J. Dairy Sci. 2002, 85, 947–950. [Google Scholar] [CrossRef]
- Fleige, S.; Preißinger, W.; Meyer, HH.; Pfaffl, M.W. Effect of lactulose on growth performance and intestinal morphology of pre-ruminant calves using a milk replacer containing Enterococcus faecium. Animal 2007, 1, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Fleige, S.; Preissinger, W.; Meyer, H.H.; Pfaffl, M.W. The immune-modulatory effect of lactulose on Enterococcus faecium fed pre-ruminant calves. J. Anim. Sci. 2009, 87, 1731–1738. [Google Scholar] [CrossRef] [PubMed]
- Yokota, A.; Hirayama, S.; Enomoto, K.; Miura, Y.; Takao, S.; Tomita, F. Production of inulin fructotransferase (de-polymerizing) by Arthrobacter sp. H65-7 and preparation of DFA III from inulin by the enzyme. J. Ferm. Bioeng. 1991, 72, 258–261. [Google Scholar] [CrossRef]
- Tamura, A.; Shiomi, T.; Tamaki, N.; Shigematsu, N.; Tomita, F.; Hara, H. Comparative effect of repeated ingestion of difructose anhydride III and palatinose on the induction of gastrointestinal symptom in human. Biosci. Biotechnol. Biochem. 2004, 68, 1882–1887. [Google Scholar] [CrossRef] [PubMed]
- Mineo, H.; Hara, H.; Shigematsu, N.; Okuhara, Y.; Tomita, F. Melibiose, difructose anhydride III and difructose anhydride IV enhance net calcium absorption in rat small and large intestinal epithelium by increasing the passage of tight junctions in vitro. J. Nutr. 2002, 132, 3394–3399. [Google Scholar] [CrossRef] [PubMed]
- Shiga, K.; Hara, H.; Okano, G.; Ito, M.; Minami, A.; Tomita, F. Ingestion of difructose anghdride III and voluntary running exercise independently increase femoral and tibial bone mineral density and bone strength with increasing calcium absorption in rats. J. Nutr. 2003, 123, 4207–4211. [Google Scholar] [CrossRef] [PubMed]
- Shigematsu, N.; Okuhara, Y.; Shiomi, T.; Tomita, F.; Hara, H. Effect of difructose anhydride III on calcium absorption in humans. Biosci. Biotechnol. Biochem. 2004, 68, 1011–1016. [Google Scholar] [CrossRef] [PubMed]
- Tomita, K.; Shiomi, T.; Okuhara, Y.; Tamura, A.; Shigematsu, N.; Hara, H. Ingestion of difructose anhydride III enhances absorption and retention of calcium in healthy men. Biosci. Biotechnol. Biochem. 2007, 71, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Nakai, T.; Sadoya, H.; Ohtani, M.; Hanada, M.; Okamoto, M. Effect of difructose anhydride III on hypocalcemia in dairy cows after calving. Anim. Sci. J. 2007, 78, 37–43, (In Japanese with English Abstract). [Google Scholar] [CrossRef]
- Teramura, M.; Wynn, S.; Reshalaitihan, M.; Kyuno, W.; Sato, T.; Ohtani, M.; Kawashima, C.; Hanada, M. Supplementation with difructose anhydride III promotes passive calcium absorption in the small intestine immediately after calving in dairy cows. J. Dairy Sci. 2015, 98, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Minamida, K.; Shiga, K.; Sujaya, I.N.; Sone, T.; Yokota, A.; Hara, H.; Asano, K.; Tomita, F. Effects of difructose anhydride III (DFA III) administration on rat intestinal microbiota. J. Biosci. Bioeng. 2005, 99, 230–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minamida, K.; Ohashi, M.; Hara, H.; Asano, K.; Tomita, F. Effects of ingestion of difructose anhydride III (DFA III) and the DFA III-assimilating bacterium Ruminococcus products on rat intestine. Biosci. Biotechnol. Biochem. 2006, 70, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, D.; Takagi, M.; Hasunuma, H.; Fushimi, Y.; Ohtani, M.; Sato, T.; Okamoto, K.; Shahada, F.; Tanaka, T.; Deguchi, E. Effects of oral administration of difructose anhydride III on selected health and blood parameters of group-housed Japanese Black calves during the preweaning period. Asian Aust. J. Anim. Sci. 2009, 22, 1640–1647. [Google Scholar] [CrossRef]
- Takagi, M.; Hasunuma, H.; Matsumoto, D.; Obi, T.; Takase, K.; Ohtani, M.; Sato, T.; Watanabe, U.; Okamoto, K.; Tanaka, T.; et al. Effects of daily oral administration of difructose anhydride III on health status, blood parameters and faecal shedding of coliform bacteria of Japanese Black calves during the pre-weaning period. Anim. Nutr. Feed. Technol. 2011, 11, 147–158. [Google Scholar]
- Akbari, P.; Braber, S.; Alizadeh, A.; Verheijden, K.A.; Schoterman, M.H.; Kraneveld, A.D.; Garssen, J.; Fink-Gremmels, J. Galacto-oligosaccharides protect the intestinal barrier by maintaining the tight junction network and modulating the inflammatory responses after a challenge with the mycotoxin deoxynivalenol in human Caco-2 cell monolayers and B6C2F1 mice. J. Nutr. 2015, 29, 1604–1613. [Google Scholar] [CrossRef] [PubMed]
- Akbari, P.; Braber, S.; Varasteh, S.; Alizadeh, A.; Garssen, J.; Fink-Gremmels, J. The intestinal barrier as an emerging target in the toxicological assessment of mycotoxins. Arch. Toxicol. 2017, 91, 1007–1029. [Google Scholar] [CrossRef] [PubMed]
- Akbari, P.; Fink-Gremmels, J.; Willems, R.H.A.M.; Difilippo, E.; Schols, H.A.; Schoterman, M.H.C.; Garssen, J.; Braber, S. Characterizing microbiota-independent effects of oligosaccharides on intestinal epithelial cells: Insight into the role of structure and size. Eur. J. Nutr. 2017, 56, 1919–1930. [Google Scholar] [CrossRef] [PubMed]
- Malekinejad, H.; Maas-Bakker, R.; Fink-Gremmels, J. Species differences in the hepatic biotransformation of zearalenone. Vet. J. 2006, 172, 96–102. [Google Scholar] [CrossRef] [PubMed]
- El-Nekeety, A.A.; El-Kady, A.A.; Abdel-Wahhab, K.G.; Hassan, N.S.; Abdel-Wahhab, M.A. Reduction of individual or combined toxicity of fumonisin B1 and zearalenone via dietary inclusion of organo-modified nano-montmorillonite in rats. Environ. Sci. Pollut. Res. 2017, 24, 20770–20783. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Nougayrede, J.P.; Del Rio, J.C.; Moreno, C.; Marin, D.E.; Ferrier, L.; Bracarense, A.P.; Kolf-Clauw, M.; Oswald, I.P. The food contaminant deoxynivalenol, decreases intestinal barrier permeability and reduces claudin expression. Toxicol. Appl. Pharmacol. 2009, 237, 41–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajecka, M.; Zielonka, L.; Gajecki, M. Activity of zearalenone in the porcine intestinal tract. Molecules 2017, 22, 18. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Hara, H. Role of flavonoids in intestinal tight junction regulation. J. Nutri. Biochem. 2011, 22, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Goossens, J.; Pasmans, F.; Verbrugghe, E.; Vandenbroucke, V.; De Baere, S.; Meyer, E.; Haesebrouck, F.; De Backer, P.; Croubels, S. Porcine intestinal epithelial barrier disruption by the Fusarium mycotoxins deoxynivalenol and T-2 toxin promotes transepithelial passage of doxycycline and paromomycin. BMC Vet. Res. 2012, 8, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrer, M.; Manyes, L.; Mañes, J.; Meca, G. Influence of prebiotics, probiotics and protein ingredients on mycotoxin bioaccessibility. Food Funct. 2015, 6, 987–994. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, H.; Nagura, T.; Inoue, M.; Kishida, T.; Sakurai, H.; Yokota, A.; Asano, K.; Tomita, F.; Sayama, K.; Senba, Y. Physical, chemical and physiological properties of difructose anhydride III produced from inukin by enzymatic reaction. J. Appl. Glycosci. 2004, 51, 291–296. [Google Scholar] [CrossRef]
- Suzuki, T.; Hara, H.; Kasai, T.; Tomita, F. Effects of difructose anhydride III on calcium absorption in small and large intestines of rats. Biosci. Biotechnol. Biochem. 1998, 62, 837–841. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Hara, H. Various nondigestible saccharides open a paracellular valcium transport pathway with the induction of intracellular calcium signaling in human intestinal Caco-2 cells. J. Nutr. 2004, 134, 1935–1941. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Hara, H. Difructose anhydride III and sodium caprate activate paracellular transport via different intracellular events in Caco-2 cells. Life Sci. 2006, 79, 401–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mineo, H.; Amano, M.; Minaminida, K.; Chiji, H.; Shigematsu, N.; Tomita, F.; Hara, H. Two-week feeding of difructose anhydride III enhances calcium absorptive activity with epithelial cell proliferation in isolated rat cecal mucosa. Nutrition 2006, 22, 312–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Herd | Forage Feed, kg | Formula Feed | ||||||
|---|---|---|---|---|---|---|---|---|
| Total, kg | Bran, % | Cereal, % | Oil Seed Meal, % | Other, % | TDN, % | CP, % | ||
| Herd 1 | Straw 2.0, Timothy grass 2.0 | 3 | 24 | 46 | 16 | 14 | >70.0 | >16 |
| Herd 2 | Straw 2.0, Oats 2.0 | 2 | 27 | 56 | 7 | 10 | >71.5 | >14 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toda, K.; Uno, S.; Kokushi, E.; Shiiba, A.; Hasunuma, H.; Matsumoto, D.; Ohtani, M.; Yamato, O.; Shinya, U.; Wijayagunawardane, M.; et al. Fructo-Oligosaccharide (DFA III) Feed Supplementation for Mitigation of Mycotoxin Exposure in Cattle—Clinical Evaluation by a Urinary Zearalenone Monitoring System. Toxins 2018, 10, 223. https://doi.org/10.3390/toxins10060223
Toda K, Uno S, Kokushi E, Shiiba A, Hasunuma H, Matsumoto D, Ohtani M, Yamato O, Shinya U, Wijayagunawardane M, et al. Fructo-Oligosaccharide (DFA III) Feed Supplementation for Mitigation of Mycotoxin Exposure in Cattle—Clinical Evaluation by a Urinary Zearalenone Monitoring System. Toxins. 2018; 10(6):223. https://doi.org/10.3390/toxins10060223
Chicago/Turabian StyleToda, Katsuki, Seiichi Uno, Emiko Kokushi, Ayaka Shiiba, Hiroshi Hasunuma, Daisaku Matsumoto, Masayuki Ohtani, Osamu Yamato, Urara Shinya, Missaka Wijayagunawardane, and et al. 2018. "Fructo-Oligosaccharide (DFA III) Feed Supplementation for Mitigation of Mycotoxin Exposure in Cattle—Clinical Evaluation by a Urinary Zearalenone Monitoring System" Toxins 10, no. 6: 223. https://doi.org/10.3390/toxins10060223
APA StyleToda, K., Uno, S., Kokushi, E., Shiiba, A., Hasunuma, H., Matsumoto, D., Ohtani, M., Yamato, O., Shinya, U., Wijayagunawardane, M., Fink-Gremmels, J., Taniguchi, M., & Takagi, M. (2018). Fructo-Oligosaccharide (DFA III) Feed Supplementation for Mitigation of Mycotoxin Exposure in Cattle—Clinical Evaluation by a Urinary Zearalenone Monitoring System. Toxins, 10(6), 223. https://doi.org/10.3390/toxins10060223