Application of Low-Fermenting Yeast Lachancea thermotolerans for the Control of Toxigenic Fungi Aspergillus parasiticus, Penicillium verrucosum and Fusarium graminearum and Their Mycotoxins



Abstract
:1. Introduction
2. Results and Discussion
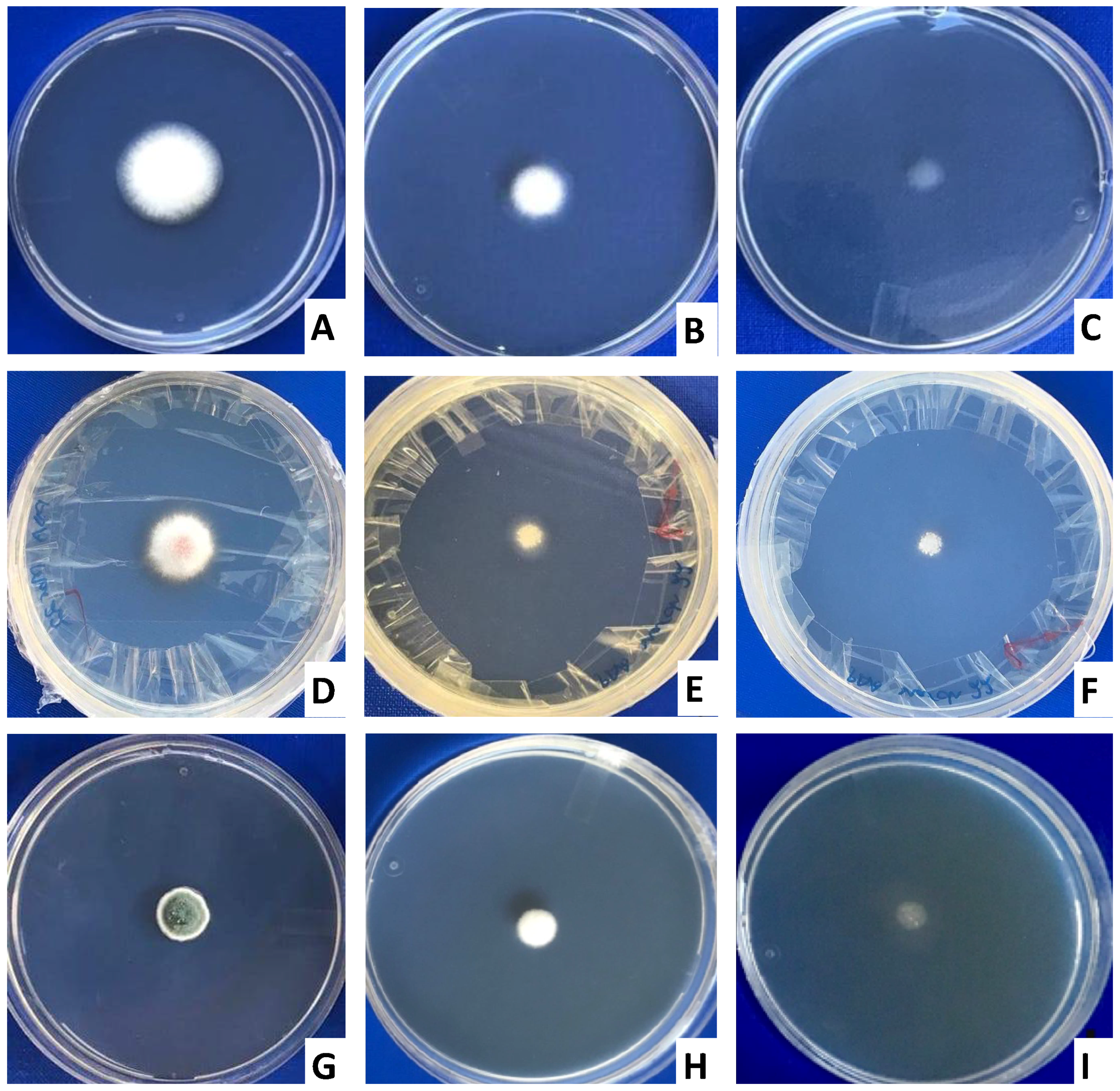
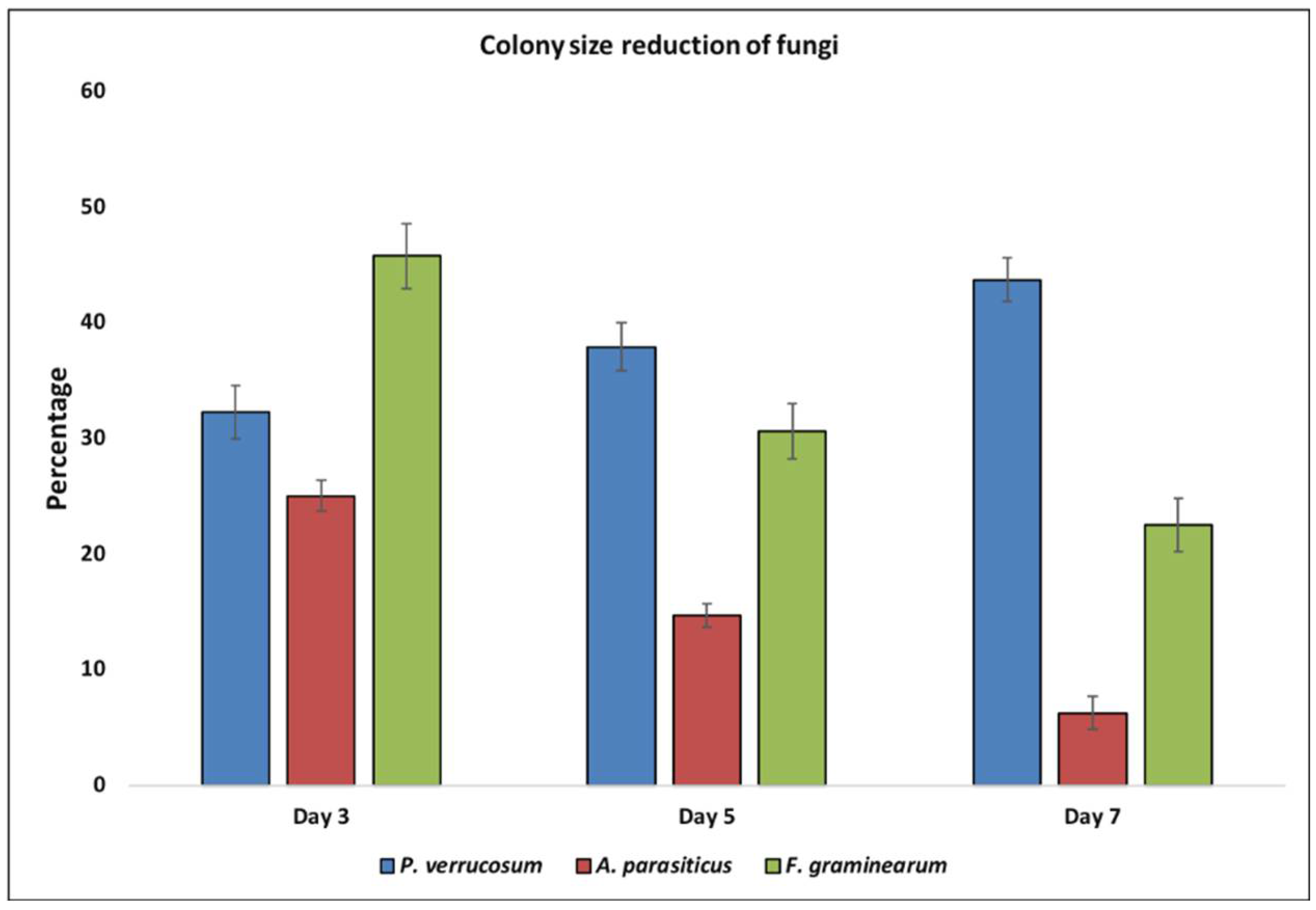
2.1. Biocontrol Activity of Yeast VOCs against Toxigenic Fungi
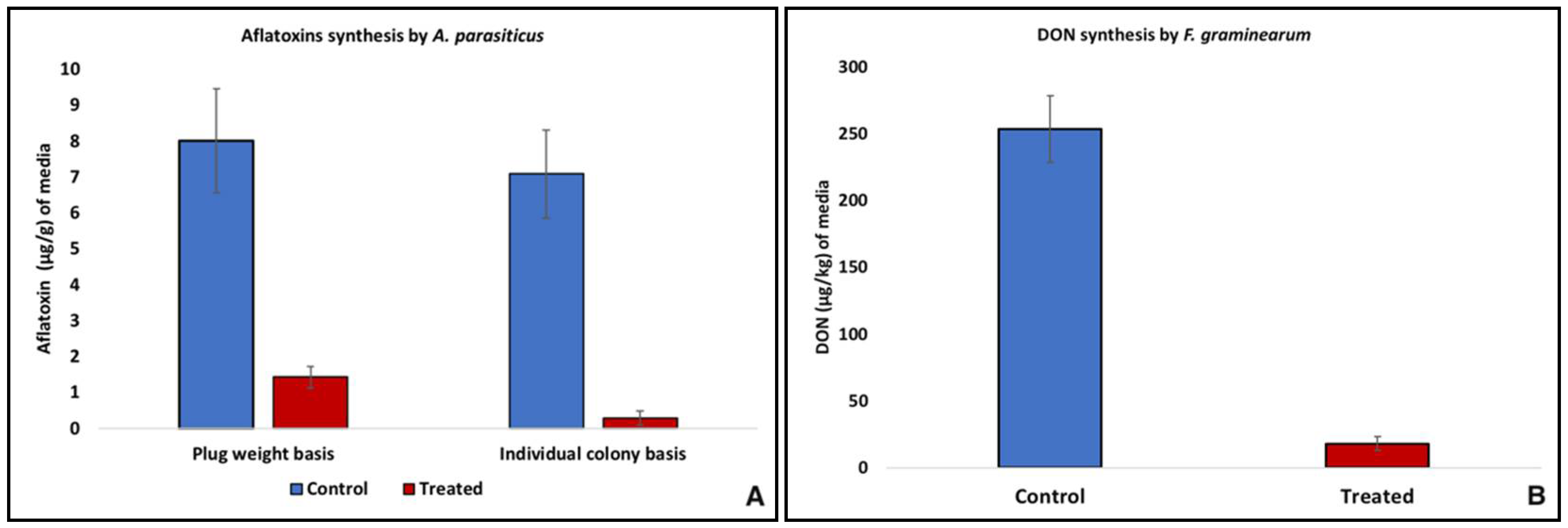
2.2. Effect of Yeast VOCs on the Mycotoxins Synthesis
2.3. Effect of No. of Yeast CFU on Fungal Growth and Mycotoxins Synthesis
2.4. Role of Nutrients Availability to Yeast on Its Antagonistic Activities against F. graminearum
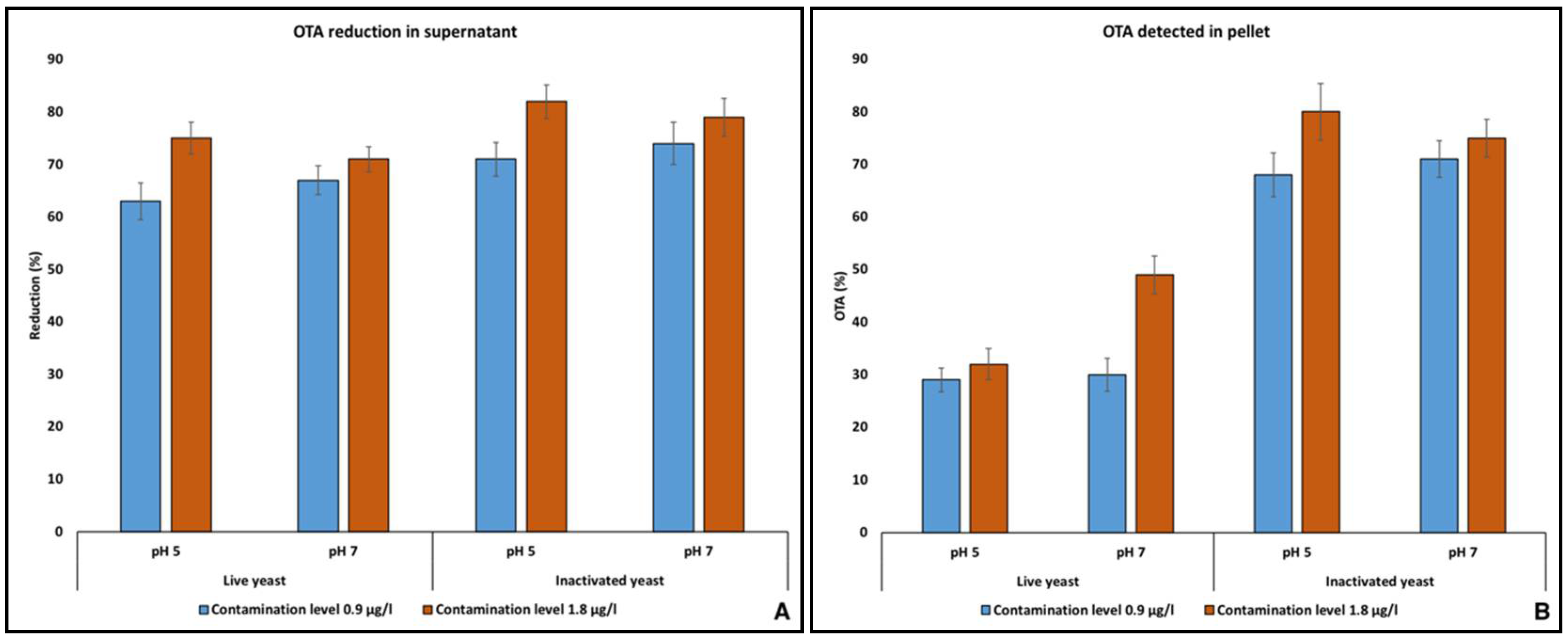
2.5. Mycotoxins Binding onto Yeast Cell Wall (YCW)
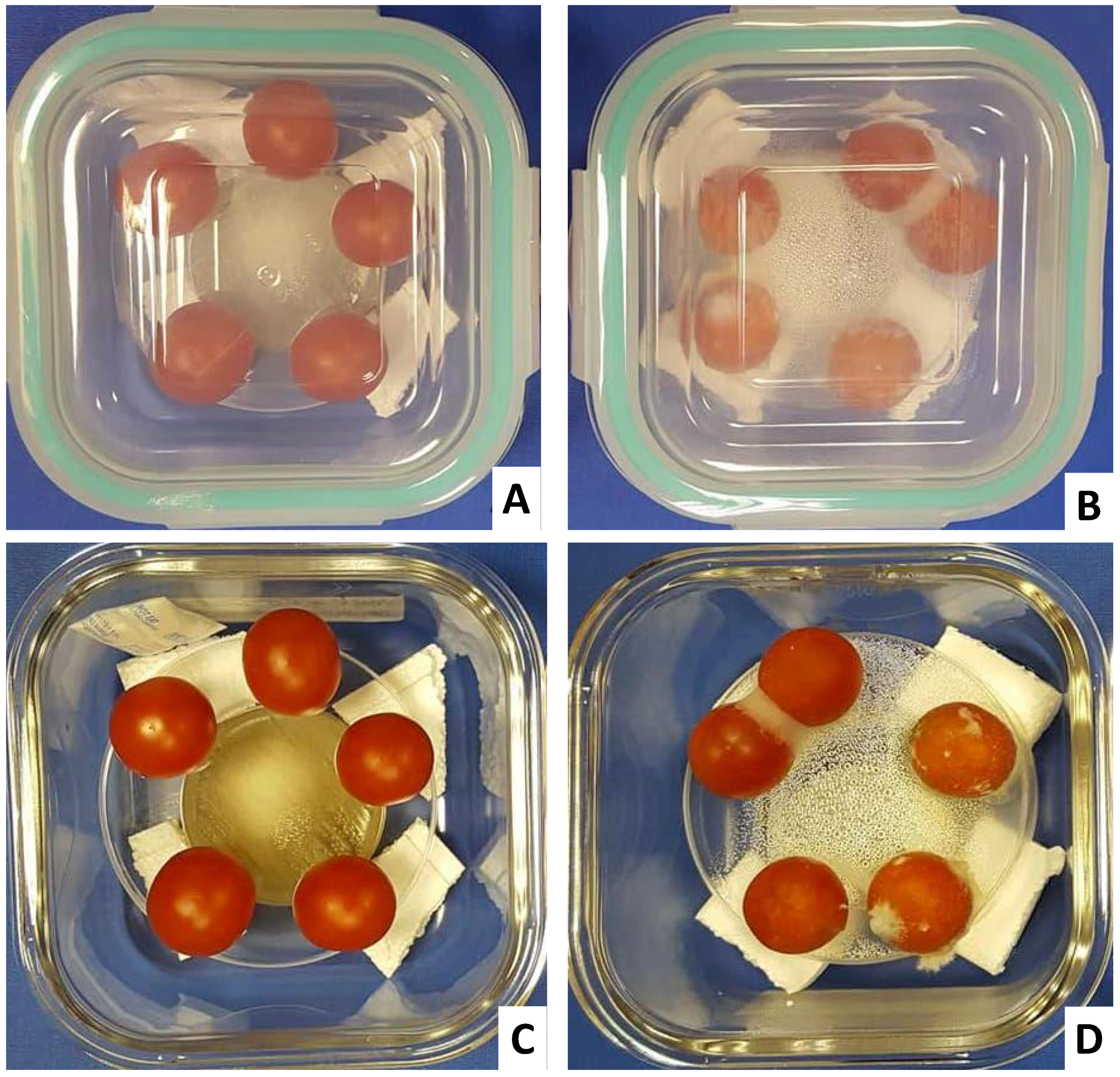
2.6. VOCs Inhibits F. oxysporum Infection in Tomato
3. Conclusions
4. Materials and Methods
4.1. Chemicals, Supplies and Biological Strains
4.2. Effect of Yeast VOCs on Growth and Sporulation of Toxigenic Fungi
4.3. Effect of Yeast VOCs on the Mycotoxins Synthesis
4.4. Effect of Number of Yeast Colony Forming Units (CFU) on F. graminearum Growth and DON Synthesis
4.5. Effect of Nutrient Availability to Yeast on Its Antagonistic Activities
4.6. In-Vitro Mycotoxins Binding Experiments
4.7. In-Vitro Testing of Yeast VOCs against F. oxysporum Infection in Tomato
4.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Milani, J.M. Ecological conditions affecting mycotoxin production in cereals: A review. Vet. Med. 2013, 58, 405–411. [Google Scholar] [CrossRef]
- Tola, M.; Kebede, B. Occurrence, importance and control of mycotoxins: A review. Cogent Food Agric. 2016, 2. [Google Scholar] [CrossRef]
- Bhat, R.; Rai, R.V.; Karim, A.A. Mycotoxins in food and feed: Present status and future concerns. Compr. Rev. Food Sci. Food Saf. 2010, 9, 57–81. [Google Scholar] [CrossRef]
- Mazumder, P.M.; Sasmal, D. Mycotoxins—Limits and regulations. Anc. Sci. Life 2001, 20, 1–19. [Google Scholar] [PubMed]
- Abbas, H.K.; Wilkinson, J.R.; Zablotowicz, R.M.; Accinelli, C.; Abel, C.A.; Bruns, H.A.; Weaver, M.A. Ecology of Aspergillus flavus, regulation of aflatoxin production and management strategies to reduce aflatoxin contamination of corn. Toxin Rev. 2009, 2–3, 142–152. [Google Scholar] [CrossRef]
- Pankaj, S.K.; Shi, H.; Keener, K.M. A review of novel physical and chemical decontamination technologies for aflatoxin in food. Trends Food Sci. Technol. 2018, 71, 73–83. [Google Scholar] [CrossRef]
- Shanakhat, H.; Sorrentino, A.; Raiola, A.; Annalisa, R.; Paolo, M.; Silvana, C. Current methods for mycotoxins analysis and innovative strategies for their reduction in cereals: An overview. J. Sci. Food Agric. 2018. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, B.L.; Carolina, F.S.C.C.; Diane, V.N.; Carlos, H.C.; Carlos, A.F.O. Mycotoxins in fruits and fruit-based products: Occurrence and methods for decontamination. Toxin Rev. 2018. [Google Scholar] [CrossRef]
- Varga, J.; Kocsubé, S.; Péteri, Z.; Vágvölgyi, C.; Tóth, B. Chemical, physical and biological approaches to prevent ochratoxin induced toxicoses in humans and animals. Toxins 2010, 2, 1718–1750. [Google Scholar] [CrossRef] [PubMed]
- Faucet-Marquis, V.; Joannis-Cassan, C.; Hadjeba-Medjdoub, K.; Ballet, N.; Pfohl-Leszkowicz, A. Development of an in vitro method for the prediction of mycotoxin binding on yeast-based products: Case of aflatoxin B1, zearalenone and ochratoxin A. Appl. Microbiol. Biotechnol. 2014, 98, 7583–7596. [Google Scholar] [CrossRef] [PubMed]
- Tsitsigiannis, D.I.; Dimakopoulou, M.; Antoniou, P.P.; Tjamos, E.C. Biological control strategies of mycotoxigenic fungi and associated mycotoxins in Mediterranean basin crops. Phytopathol. Mediterr. 2012, 51, 158–174. [Google Scholar]
- Kabak, B.; Dobson, A.D.W.; Var, I. Strategies to prevent mycotoxin contamination of food and animal feed: A review. Crit. Rev. Food Sci. Nutr. 2006, 46, 593–619. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, J.D.; O’Keeffe, T.L.; Abbas, H.K. Microbial interactions with mycotoxigenic fungi and mycotoxins. Toxin Rev. 2008, 27, 261–285. [Google Scholar] [CrossRef]
- Liu, J.; Sui, Y.; Wisniewski, M.; Droby, S.; Liu, Y. Review: Utilization of antagonistic yeasts to manage postharvest fungal diseases of fruit. Int. J. Food Microbiol. 2013, 167, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Caridi, A. New perspectives in safety and quality enhancement of wine through selection of yeasts based on the parietal adsorption activity. Int. J. Food Microbiol. 2007, 120, 167–172. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.P. Microbial detoxification of mycotoxins. J. Chem. Ecol. 2013, 39, 907–918. [Google Scholar] [CrossRef] [PubMed]
- Halász, A.; Lásztity, R.; Abonyi, T.; Bata, Á. Decontamination of mycotoxin-containing food and feed by biodegradation. Food Rev. Int. 2009, 25, 284–298. [Google Scholar] [CrossRef]
- Schatzmayr, G.; Zehner, F.; Täubel, M.; Schatzmayr, D.; Klimitsch, A.; Loibner, A.P.; Binder, E.M. Microbiologicals for deactivating mycotoxins. Mol. Nutr. Food Res. 2006, 50, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Virgili, R.; Simoncini, N.; Toscani, T.; Camardo, L.; Formenti, S.; Battilani, P. Biocontrol of Penicillium nordicum growth and ochratoxin a production by native yeasts of dry cured ham. Toxins 2012, 4, 68–82. [Google Scholar] [CrossRef] [PubMed]
- Hua, S.S.T.; Beck, J.J.; Sarreal, S.B.L.; Gee, W. The major volatile compound 2-phenylethanol from the biocontrol yeast, Pichia anomala, inhibits growth and expression of aflatoxin biosynthetic genes of Aspergillus flavus. Mycotoxin Res. 2014, 30, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Schisler, D.; Boehm, M.; Lipps, P.; Slininger, P. Field testing of antagonists of Fusarium head blight incited by Gibberella zeae. Biol. Control 2004, 29, 245–255. [Google Scholar] [CrossRef]
- Fiori, S.; Urgeghe, P.P.; Hammami, W.; Razzu, S.; Jaoua, S.; Migheli, Q. Biocontrol activity of four non- and low-fermenting yeast strains against Aspergillus carbonarius and their ability to remove ochratoxin A from grape juice. Int. J. Food Microbiol. 2014, 17, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.K.; Hua, S.S.T.; Sarreal, S.B.L.; Li, R.W. Suppression of aflatoxin biosynthesis in Aspergillus flavus by 2-Phenylethanol is associated with stimulated growth and decreased degradation of branched-chain amino acids. Toxins 2015, 7, 3887–3902. [Google Scholar] [CrossRef] [PubMed]
- Nazareth, T.M.; Corrêa, J.A.; Pinto, A.C.; Palma, J.B.; Meca, G.; Bordin, K.; Luciano, F.B. Evaluation of gaseous allyl isothiocyanate against the growth of mycotoxigenic fungi and mycotoxin production in corn stored for 6 months. J. Sci. Food Agric. 2018. [CrossRef] [PubMed]
- Šari, L.; Škrinjar, M.M.; Saka, M.B.; Plavši, D.V.; Abarkapa, I.V. Effect of the baker′s yeast on the production of aflatoxin B1 by Aspergillus flavus. Food Prog. Qual. Saf. 2008, 4, 165–168. [Google Scholar]
- Armando, M.R.; Pizzolitto, R.P.; Dogi, C.A.; Cristofolini, A.; Merkis, C.; Poloni, V.; Dalcero, A.M.; Cavaglieri, L.R. Adsorption of ochratoxin A and zearalenone by potential probiotic Saccharomyces cerevisiae strains and its relation with cell wall thickness. J. Appl. Microbiol. 2012, 113, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Dawson, K.A. Application of yeast and yeast derivatives in poultry industry. Proc. Aust. Poult. Sci. Symp. 2011, 13, 100–105. [Google Scholar]
- Piotrowska, M.; Masek, A. Saccharomyces cerevisiae cell wall components as tools for ochratoxin A decontamination. Toxins 2015, 7, 1151–1162. [Google Scholar] [CrossRef] [PubMed]
- Mil, T.D.; Devreese, M.; Baere, S.D.; Ranst, E.V.; Eeckhout, M.; Backer, P.D.; Croubels, S. Characterization of 27 mycotoxin binders and the relation with in vitro zearalenone adsorption at a single concentration. Toxins 2015, 7, 21–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devegowda, G.; Rayu, M.V.L.N.; Swamy, H.V.L.N. Mycotoxins: Novel solutions for their counteraction. Feedstuffs 1998, 7, 12–15. [Google Scholar]
- Jouany, J.P.; Yiannikouris, A.; Bertin, G. The chemical bonds between mycotoxins and cell wall components of Saccharomyces cerevisiae have been identified. Arch. Zootech. 2005, 8, 26–50. [Google Scholar]
- Ringot, D.; Lerzy, B.; Bonhoure, J.P.; Auclair, E.; Oriol, E.; Larondelle, Y. Effect of temperature on in vitro ochratoxin biosorption onto yeast cell wall derivatives. Process Biochem. 2005, 40, 3008–3016. [Google Scholar] [CrossRef]
- Joannis-Cassan, C.; Tozlovanu, M.; Hadjeba-Medjdoub, K.; Ballet, N.; Pfohl-Leszkowicz, A. Binding of zearalenone, aflatoxin B1, and ochratoxin A by yeast-based products: A method for quantification of adsorption performance. J. Food Prot. 2011, 74, 1175–1185. [Google Scholar] [CrossRef] [PubMed]
- Hassan, Z.U.; Al-Thani, R.F.; Migheli, Q.; Jaoua, S. Detection of toxigenic mycobiota and mycotoxins in cereal feed market. Food Control 2018, 84, 389–394. [Google Scholar] [CrossRef]
- Bzducha-Wróbel, A.; Błażejak, S.; Kawarska, A.; Stasiak-Różańska, L.; Gientka, I.; Majewska, E. Evaluation of the efficiency of different disruption methods on yeast cell wall preparation for β-Glucan isolation. Molecules 2014, 19, 20941–20961. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Colony Diameter (mm) | DON Synthesis (µg/kg) | Colony Diameter and DON Synthesis by Fungi Not Exposed to Yeast VOCs | |
|---|---|---|---|---|
| Control (No yeast) | 30 ± 1.0 a | 311.48 ± 33.78 a | Colony diameter (mm) | DON synthesis (µg/kg) |
| 1X YPDA | 8 ± 0.6 c | 218.09 ± 29.52 b | 30 ± 0.7 | 331.00 ± 41.09 |
| 0.5X YPDA | 20 ± 0.8 b | 203.04 ± 8.69 b | 32 ± 0.6 | 340.44 ± 35.00 |
| 0.1X YPDA | 28 ± 0.7 a | 164.83 ± 19.22 c | 29 ± 0.6 | 312.32 ± 30.33 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeidan, R.; Ul-Hassan, Z.; Al-Thani, R.; Balmas, V.; Jaoua, S. Application of Low-Fermenting Yeast Lachancea thermotolerans for the Control of Toxigenic Fungi Aspergillus parasiticus, Penicillium verrucosum and Fusarium graminearum and Their Mycotoxins. Toxins 2018, 10, 242. https://doi.org/10.3390/toxins10060242
Zeidan R, Ul-Hassan Z, Al-Thani R, Balmas V, Jaoua S. Application of Low-Fermenting Yeast Lachancea thermotolerans for the Control of Toxigenic Fungi Aspergillus parasiticus, Penicillium verrucosum and Fusarium graminearum and Their Mycotoxins. Toxins. 2018; 10(6):242. https://doi.org/10.3390/toxins10060242
Chicago/Turabian StyleZeidan, Randa, Zahoor Ul-Hassan, Roda Al-Thani, Virgilio Balmas, and Samir Jaoua. 2018. "Application of Low-Fermenting Yeast Lachancea thermotolerans for the Control of Toxigenic Fungi Aspergillus parasiticus, Penicillium verrucosum and Fusarium graminearum and Their Mycotoxins" Toxins 10, no. 6: 242. https://doi.org/10.3390/toxins10060242
APA StyleZeidan, R., Ul-Hassan, Z., Al-Thani, R., Balmas, V., & Jaoua, S. (2018). Application of Low-Fermenting Yeast Lachancea thermotolerans for the Control of Toxigenic Fungi Aspergillus parasiticus, Penicillium verrucosum and Fusarium graminearum and Their Mycotoxins. Toxins, 10(6), 242. https://doi.org/10.3390/toxins10060242