Recombinant and Chimeric Disintegrins in Preclinical Research

 ,
,  ,
, 
Abstract
:1. Introduction
1.1. Historical Overview
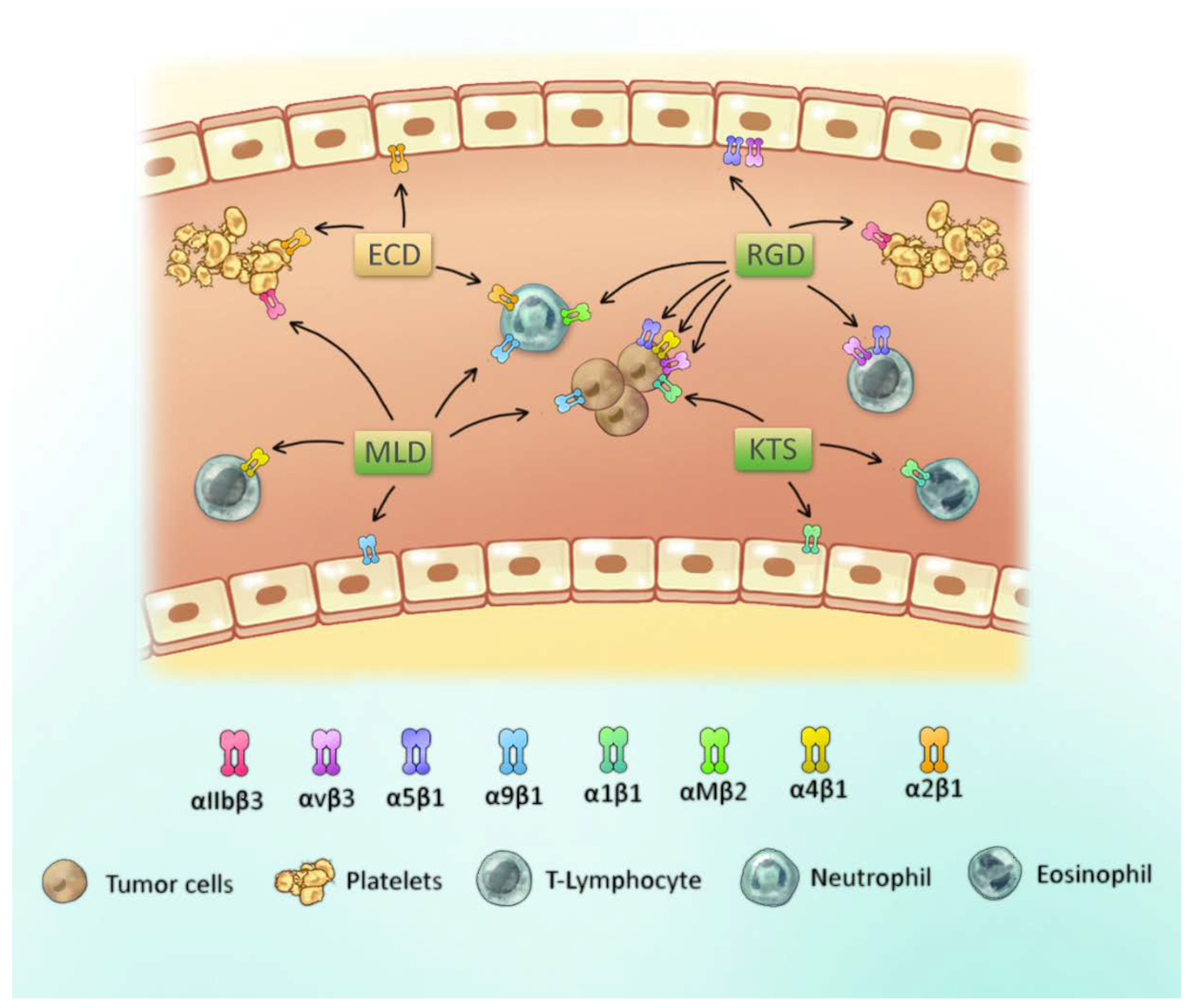
1.2. Integrins on Platelets, Tumors and Leucocytes
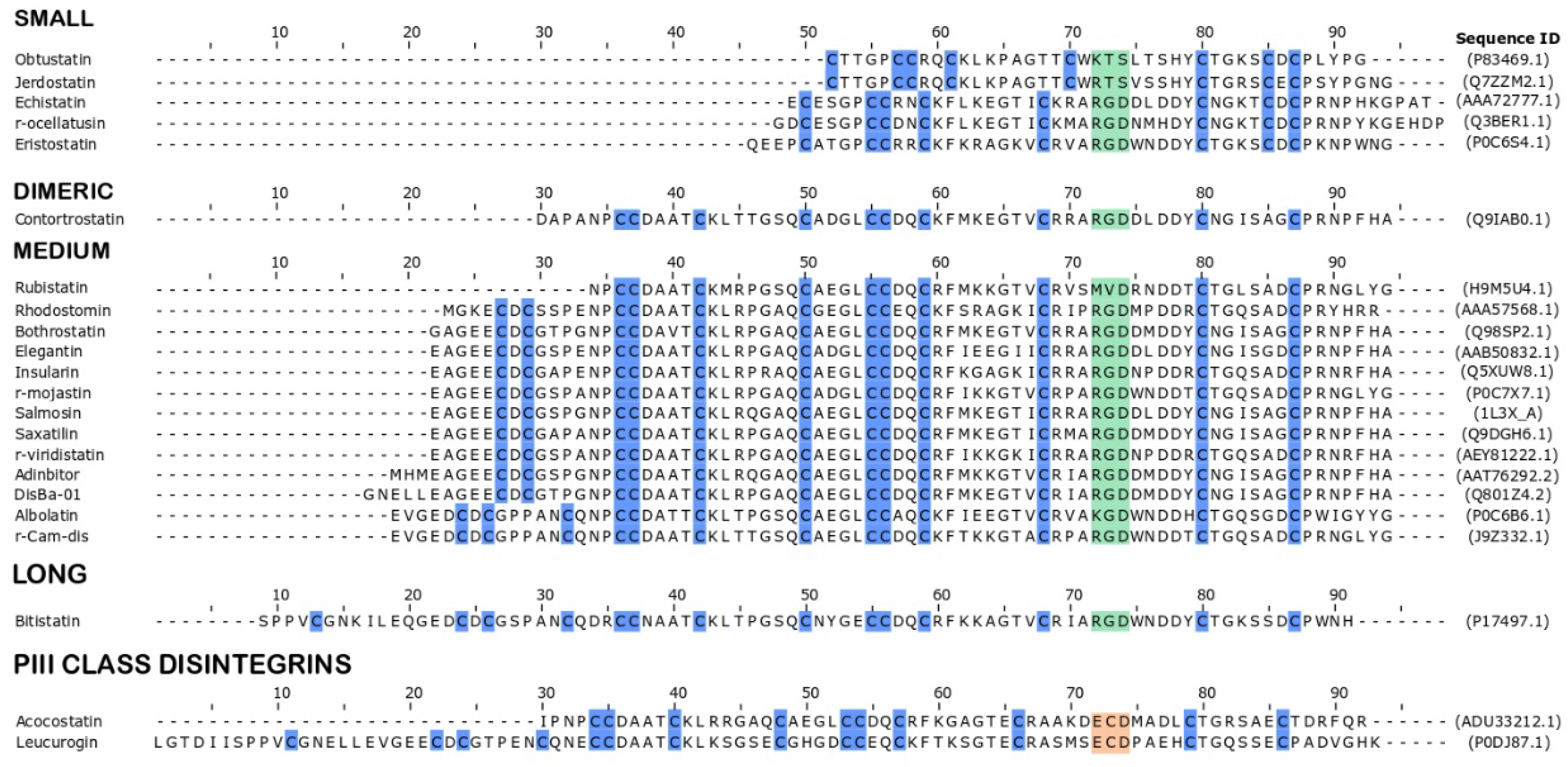
1.3. Structural Classification of Snake Venom Disintegrins
1.4. Functional Classification of Snake Venom Disintegrins
2. Recombinant Disintegrins
2.1. Disintegrins from Heterologous Expression
2.2. Mutagenesis of Disintegrins
2.3. Chimeric Disintegrins
3. Medical Relevance of Snake Venom Disintegrins
3.1. Disintegrins as Modulators of Integrin Activity
3.2. Recombinant and Chimeric Disintegrins in Preclinical Studies
4. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tcheng, J.E.; Harrington, R.A.; Kottke-Marchant, K.; Kleiman, N.S.; Ellis, S.G.; Kereiakes, D.J.; Mick, M.J.; Navetta, F.I.; Smith, J.E.; Worley, S.J.; et al. Multicenter, Randomized, Double-Blind, Placebo-Controlled Trial of the Platelet Integrin Glycoprotein IIb/IIIa Blocker Integrelin in Elective Coronary Intervention. Circulation 1995, 91, 2151–2157. [Google Scholar] [CrossRef] [PubMed]
- Gould, R.J.; Polokoff, M.A.; Friedman, P.A.; Huang, T.F.; Holt, J.C.; Cook, J.J.; Niewiarowski, S. Disintegrins: A family of integrin inhibitory proteins from viper venoms. Proc. Soc. Exp. Biol. Med. 1990, 195, 168–171. [Google Scholar] [CrossRef] [PubMed]
- McLane, M.; Sanchez, E.; Wong, A.; Paquette-Straub, C.; Perez, J. Disintegrins. Curr. Drug Target Cardiovasc. Hematol. Disord. 2004, 4, 327–355. [Google Scholar] [CrossRef]
- Phillips, D.R.; Jennings, L.K.; Edwards, H.H. Identification of membrane proteins mediating the interaction of human platelets. J. Cell Biol. 1980, 86, 77–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boudreau, N.J.; Jones, P.L. Extracellular matrix and integrin signalling: The shape of things to come. Biochem. J. 1999, 339, 481. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R. Integrins: A family of cell surface receptors. Cell 1987, 48, 549–554. [Google Scholar] [CrossRef]
- Coelho, A.L.; de Freitas, M.S.; Oliveira-Carvalho, A.L.; Moura-Neto, V.; Zingali, R.B.; Barja-Fidalgo, C. Effects of jarastatin, a novel snake venom disintegrin, on neutrophil migration and actin cytoskeleton dynamics. Exp. Cell Res. 1999, 251, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Marcinkiewicz, B.C.; Vijay-Kumar, S.; Mclane, M.A.; Niewiarowski, S. Significance of RGD Loop and C-Terminal Domain of Echistatin for Recognition of αIIbβ3 and avβ3 Integrins and Expression of Ligand-Induced Binding Site. Blood 1997, 90, 1565–1576. [Google Scholar] [PubMed]
- Calvete, J.J. The continuing saga of snake venom disintegrins. Toxicon 2013, 62, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Marcinkiewicz, C. Applications of snake venom components to modulate integrin activities in cell-matrix interactions. Int. J. Biochem. Cell Biol. 2013, 45, 1974–1986. [Google Scholar] [CrossRef] [PubMed]
- Macêdo, J.K.A.; Fox, J.W.; Castro, M.D.S.; Kele, J.; Macêdo, A.; Fox, J.W.; Castro, M.D.S. Disintegrins from snake venoms and their applications in cancer research and therapy. Curr. Protein Pept. Sci. 2015, 16, 532–548. [Google Scholar] [CrossRef]
- Fox, J.W.; Serrano, S.M.T. Timeline of key events in snake venom metalloproteinase research. J. Proteom. 2009, 72, 200–209. [Google Scholar] [CrossRef] [PubMed]
- McLane, M.A.; Marcinkiewicz, C.; Vijay-Kumar, S.; Wierzbicka-Patynowski, I.; Niewiarowski, S. Viper Venom Disintegrins and Related Molecules. Exp. Biol. Med. 1998, 219, 109–119. [Google Scholar] [CrossRef]
- Calvete, J.J.; Marcinkiewicz, C.; Monleón, D.; Esteve, V.; Celda, B.; Juárez, P.; Sanz, L. Snake venom disintegrins: Evolution of structure and function. Toxicon 2005, 45, 1063–1074. [Google Scholar] [CrossRef] [PubMed]
- Mclane, M.A.; Joerger, T.; Mahmoud, A. Disintegrins in health and disease. Front. Biosci. 2008, 2003, 6617–6637. [Google Scholar] [CrossRef]
- Huang, T.-F.; Hsu, C.-C.; Kuo, Y.-J.; Teng, C.; Huang, T.; Clemetson, K.; Hynes, R.; Ginsberg, M.; Loftus, J.; Plow, E.; et al. Anti-thrombotic agents derived from snake venom proteins. Thromb. J. 2016, 14, 18. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.F.; Holt, J.C.; Lukasiewicz, H.; Niewiarowski, S. Trigramin: A low molecular weight peptide inhibiting fibrinogen interaction with platelet receptors expressed on glycoprotein IIb-IIIa complex. J. Biol. Chem. 1987, 262, 16157–16163. [Google Scholar] [PubMed]
- Barczyk, M.; Carracedo, S.; Gullberg, D. Integrins. Cell Tissue Res. 2010, 339, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Goodman, S.L.; Picard, M. Integrins as therapeutic targets. Trends Pharmacol. Sci. 2012, 33, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Savage, B.; Saldívar, E.; Ruggeri, Z.M. Initiation of platelet adhesion by arrest onto fibrinogen or translocation on von Willebrand factor. Cell 1996, 84, 289–297. [Google Scholar] [CrossRef]
- Calvete, J.J. Platelet integrin GPIIb/IIIa: Structure-function correlations. An update and lessons from other integrins [Review]. Proc. Soc. Exp. Biol. Med. 1999, 222, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Nieberler, M.; Reuning, U.; Reichart, F.; Notni, J.; Wester, H.-J.; Schwaiger, M.; Weinmüller, M.; Räder, A.; Steiger, K.; Kessler, H. Exploring the Role of RGD-Recognizing Integrins in Cancer. Cancers 2017, 9, 116. [Google Scholar] [CrossRef] [PubMed]
- Horton, M.A. The αvβ3 integrin “vitronectin receptor”. Int. J. Biochem. Cell Biol. 1997, 29, 721–725. [Google Scholar] [CrossRef]
- Hynes, R.O. Integrins: Versatility, modulation, and signaling in cell adhesion. Cell 1992, 69, 11–25. [Google Scholar] [CrossRef]
- Lian, J.; Guoping, C.; Shapiro, S.S.; Tran, L.-P.; Beacham, D.A. Glycoprotein Ibα Can Mediate Endothelial Cell Migration on von Willebrand Factor-Containing Substrata. Exp. Cell Res. 1999, 252, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Wong, N.C.; Mueller, B.M.; Barbas, C.F.; Ruminski, P.; Quaranta, V.; Lin, E.C.; Smith, J.W. Alphav integrins mediate adhesion and migration of breast carcinoma cell lines. Clin. Exp. Metast. 1998, 16, 50–61. [Google Scholar] [CrossRef]
- Kourtzelis, I.; Mitroulis, I.; von Renesse, J.; Hajishengallis, G.; Chavakis, T. From leukocyte recruitment to resolution of inflammation: The cardinal role of integrins. J. Leukoc. Biol. 2017, 102, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M.; Evans, H.J. Structural domains in venom proteins: Evidence that metalloproteinases and nonenzymatic platelet aggregation inhibitors (disintegrins) from snake venoms are derived by proteolysis from a common precursor. Toxicon 1992, 30, 265–293. [Google Scholar] [CrossRef]
- Calvete, J.J.; Moreno-Murciano, M.P.; Theakston, R.D.G.; Kisiel, D.G.; Marcinkiewicz, C. Snake venom disintegrins: Novel dimeric disintegrins and structural diversification by disulphide bond engineering. Biochem. J. 2003, 372, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Niewiarowski, S.; McLane, M.A.; Kloczewiak, M.; Stewart, G.J. Disintegrins and other naturally occurring antagonists of platelet fibrinogen receptors. Semin. Hematol. 1994, 31, 289–300. [Google Scholar] [PubMed]
- Lu, X.; Lu, D.; Scully, M.F.; Kakkar, V. V Integrins in drug targeting-RGD templates in toxins. Curr. Pharm. Des. 2006, 12, 2749–2769. [Google Scholar] [CrossRef] [PubMed]
- Müller, G.; Gurrath, M.; Kessler, H. Pharmacophore refinement of gpIIb/IIIa antagonists based on comparative studies of antiadhesive cyclic and acyclic RGD peptides. J. Comput. Aided Mol. Des. 1994, 8, 709–730. [Google Scholar] [CrossRef] [PubMed]
- Dechantsreiter, M.A.; Planker, E.; Mathä, B.; Lohof, E.; Hölzemann, G.; Jonczyk, A.; Goodman, S.L.; Kessler, H. N-Methylated Cyclic RGD Peptides as Highly Active and Selective α V β 3 Integrin Antagonists. J. Med. Chem. 1999, 42, 3033–3040. [Google Scholar] [CrossRef] [PubMed]
- Mas-Moruno, C.; Rechenmacher, F.; Kessler, H. Cilengitide: The First Anti-Angiogenic Small Molecule Drug Candidate. Design, Synthesis and Clinical Evaluation. Anticancer Agents Med. Chem. 2010, 10, 753–768. [Google Scholar] [CrossRef] [PubMed]
- Kapp, T.G.; Rechenmacher, F.; Neubauer, S.; Maltsev, O.V.; Cavalcanti-Adam, E.A.; Zarka, R.; Reuning, U.; Notni, J.; Wester, H.-J.; Mas-Moruno, C.; et al. A Comprehensive Evaluation of the Activity and Selectivity Profile of Ligands for RGD-binding Integrins. Sci. Rep. 2017, 7, 39805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarborough, R.M.; Rose, J.W.; Naughton, M.A.; Phillips, D.R.; Nannizzi, L.; Arfsten, A.; Campbell, A.M.; Charo, I.F. Characterization of the integrin specificities of disintegrins isolated from American pit viper venoms. J. Biol. Chem. 1993, 268, 1058–1065. [Google Scholar] [PubMed]
- Lucena, S.E.; Romo, K.; Suntravat, M.; Sánchez, E.E. Anti-angiogenic activities of two recombinant disintegrins derived from the Mohave and Prairie rattlesnakes. Toxicon 2014, 78, 10–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, E.M.; Marcinkiewicz, C. Non-RGD-containing snake venom disintegrins, functional and structural relations. Toxicon 2011, 58, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Bazan-Socha, S.; Kisiel, D.G.; Young, B.; Theakston, R.D.G.; Calvete, J.J.; Sheppard, D.; Marcinkiewicz, C. Structural requirements of MLD-containing disintegrins for functional interaction with alpha 4 beta 1 and alpha 9 beta1 integrins. Biochemistry 2004, 43, 1639–1647. [Google Scholar] [CrossRef] [PubMed]
- Marcinkiewicz, C.; Taooka, Y.; Yokosaki, Y.; Calvete, J.J.; Marcinkiewicz, M.M.; Lobb, R.R.; Niewiarowski, S.; Sheppard, D. Inhibitory Effects of MLDG-containing Heterodimeric Disintegrins Reveal Distinct Structural Requirements for Interaction of the Integrin α9β1 with VCAM-1, Tenascin-C, and Osteopontin. J. Biol. Chem. 2000, 275, 31930–31937. [Google Scholar] [CrossRef] [PubMed]
- Kisiel, D.G.; Calvete, J.J.; Katzhendler, J.; Fertala, A.; Lazarovici, P.; Marcinkiewicz, C. Structural determinants of the selectivity of KTS-disintegrins for the α1β1 integrin. FEBS Lett. 2004, 577, 478–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wierzbicka-Patynowski, I.; Niewiarowski, S.; Marcinkiewicz, C.; Calvete, J.J.; Marcinkiewicz, M.M.; McLane, M.A. Structural requirements of echistatin for the recognition of α(v)β3 and α5β1 integrins. J. Biol. Chem. 1999, 274, 37809–37814. [Google Scholar] [CrossRef] [PubMed]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front. Microbiol. 2014, 5, 172. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-C.; Cheng, C.-H.; Shiu, J.-H.; Chang, Y.-T.; Chang, Y.-S.; Huang, C.-H.; Lee, J.-C.; Chuang, W.-J. Expression in Pichia pastoris and characterization of echistatin, an RGD-containing short disintegrin. Toxicon 2012, 60, 1342–1348. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.T.; Chou, L.J.; Chen, Y.C.; Chen, C.Y.; Pari, K.; Jen, C.J.; Lo, S.J.; Huang, S.L.; Lee, C.Y.; Chang, T.W. Expression in Pichia pastoris and characterization by circular dichroism and NMR of rhodostomin. Proteins Struct. Funct. Genet. 2001, 43, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, G.S.; Novo, J.B.; Clissa, P.B.; Della Casa, M.S.; Butera, D.; Da Silva, A.M.M. Engineered mammalian vector to express EGFP-tagged proteins as biomarkers. Mol. Biotechnol. 2012, 51, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Butera, D.; Fontes Piazza, R.M.; McLane, M.A.; Chammas, R.; Da Silva, A.M.M. Molecular engineering of an EGFP/disintegrin-based integrin marker. Toxicon 2005, 46, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Knight, L.C.; Romano, J.E.; Bright, L.T.; Agelan, A.; Kantor, S.; Maurer, A.H. Platelet binding and biodistribution of [99mTc]rBitistatin in animal species and humans. Nucl. Med. Biol. 2007, 34, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Demain, A.L.; Vaishnav, P. Production of recombinant proteins by microbes and higher organisms. Biotechnol. Adv. 2009, 27, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Young, C.L.; Britton, Z.T.; Robinson, A.S. Recombinant protein expression and purification: A comprehensive review of affinity tags and microbial applications. Biotechnol. J. 2012, 7, 620–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terpe, K. Overview of bacterial expression systems for heterologous protein production: From molecular and biochemical fundamentals to commercial systems. Appl. Microbiol. Biotechnol. 2006, 72, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Kelley, M.J.; Crowl, R.M.; Dennis, E.A. Renaturation of cobra venom phospholipase A2 expressed from a synthetic gene in Escherichia coli. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1992, 1118, 107–115. [Google Scholar] [CrossRef]
- Chang, T.-Y.; Mao, S.-H.; Guo, Y.-W. Cloning and expression of a cysteine-rich venom protein from Trimeresurus mucrosquamatus (Taiwan Habu). Toxicon 1997, 35, 879–888. [Google Scholar] [CrossRef]
- Chang, L.; Lin, J.; Wu, P.; Chang, C.; Hong, E. cDNA Sequence Analysis and Expression of κ-Bungarotoxin from Taiwan Banded Krait. Biochem. Biophys. Res. Commun. 1997, 230, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.H.; Hu, S.T.; Huang, T.F.; Chen, S.H.; Lee, Y.H.W.; Lo, S.C.J. Rhodostomin, an RGD-Containing Peptide Expressed from a Synthetic Gene in Escherichia coli, Facilitates the Attachment of Human Hepatoma Cells. Biochem. Biophys. Res. Commun. 1993, 190, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S.; Aitken, A.; Flynn, G.; Formstone, C.; Savidge, G.F. Modulation of RGD sequence motifs regulates disintegrin recognition of αIIbβ3 and α5β1 integrin complexes. Biochem. J. 1998, 335, 247–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauskot, A.; Cominetti, M.R.; Ramos, O.H.P.; Bechyne, I.; Renard, J.-M.; Hoylaerts, M.F.; Crepin, M.; Legrand, C.; Selistre-de-Araujo, H.S.; Bonnefoy, A. Hemostatic effects of recombinant DisBa-01, a disintegrin from Bothrops alternatus. Front. Biosci. 2008, 13, 6604–6616. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.E.; Lucena, S.E.; Reyes, S.; Soto, J.G.; Cantu, E.; Lopez-Johnston, J.C.; Guerrero, B.; Salazar, A.M.; Rodríguez-acosta, A.; Galán, J.A.; et al. Cloning, expression, and hemostatic activities of a disintegrin, r-mojastin 1, from the mohave rattlesnake (Crotalus scutulatus scutulatus). Thromb. Res. 2010, 126, e211–e219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, D.; Kang, I.; Kim, H.; Chung, K.; Kim, D.S.; Yun, Y. Cloning and characterization of novel disintegrins from Agkistrodon halys venom. Mol. Cells 1998, 8, 578–584. [Google Scholar] [PubMed]
- Brown, M.C.; Eble, J.A.; Calvete, J.J.; Marcinkiewicz, C. Structural requirements of KTS-disintegrins for inhibition of alpha(1)beta(1) integrin. Biochem. J. 2009, 417, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Sanz, L.; Chen, R.-Q.; Pérez, A.; Hilario, R.; Juárez, P.; Marcinkiewicz, C.; Monleón, D.; Celda, B.; Xiong, Y.-L.; Pérez-Payá, E.; et al. cDNA Cloning and Functional Expression of Jerdostatin, a Novel RTS-disintegrin from Trimeresurus jerdonii and a Specific Antagonist of the α 1 β 1 Integrin. J. Biol. Chem. 2005, 280, 40714–40722. [Google Scholar] [CrossRef] [PubMed]
- Teklemariam, T.; Seoane, A.I.; Ramos, C.J.; Sanchez, E.E.; Lucena, S.E.; Perez, J.C.; Mandal, S.A.; Soto, J.G. Functional analysis of a recombinant PIII-SVMP, GST-acocostatin; an apoptotic inducer of HUVEC and HeLa, but not SK-Mel-28 cells. Toxicon 2011, 57, 646–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carey, C.M.; Bueno, R.; Gutierrez, D.A.; Petro, C.; Lucena, S.E.; Sanchez, E.E.; Soto, J.G. Recombinant rubistatin (r-Rub), an MVD disintegrin, inhibits cell migration and proliferation, and is a strong apoptotic inducer of the human melanoma cell line SK-Mel-28. Toxicon 2012, 59, 241–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berlec, A.; Strukelj, B. Current state and recent advances in biopharmaceutical production in Escherichia coli, yeasts and mammalian cells. J. Ind. Microbiol. Biotechnol. 2013, 40, 257–274. [Google Scholar] [CrossRef] [PubMed]
- Latifi, A.M.; Khajeh, K.; Farnoosh, G.; Hassanpour, K.; Khodi, S. The Cytoplasmic and Periplasmic Expression Levels and Folding of Organophosphorus Hydrolase Enzyme in Escherichia coli. Jundishapur J. Microbiol. 2015, 8, 17790. [Google Scholar] [CrossRef] [PubMed]
- Ke, N.; Berkmen, M. Production of Disulfide-Bonded Proteins in Escherichia coli. Curr. Protoc. Mol. Biol. 2014, 108, 16.1B.1–16.1B.21. [Google Scholar] [CrossRef] [PubMed]
- Ritz, D.; Beckwith, J. Roles of Thiol-Redox Pathways in Bacteria. Annu. Rev. Microbiol. 2001, 55, 21–48. [Google Scholar] [CrossRef] [PubMed]
- Prinz, W.A.; Åslund, F.; Holmgren, A.; Beckwith, J. The Role of the Thioredoxin and Glutaredoxin Pathways in Reducing Protein Disulfide Bonds in the Escherichia coli Cytoplasm. J. Biol. Chem. 1997, 272, 15661–15667. [Google Scholar] [CrossRef] [PubMed]
- Schein, C.H. Production of Soluble Recombinant Proteins in Bacteria. Nat. Biotechnol. 1989, 7, 1141–1149. [Google Scholar] [CrossRef]
- Fahnert, B.; Lilie, H.; Neubauer, P. Inclusion Bodies: Formation and Utilisation. In Physiological Stress Response in Bioprocesses. Advances in Biochemical Engineering; Springer: Heidelberg/Berlin, Germany, 2004; Volume 89, pp. 93–142. ISBN 978-3-540-20311-7. [Google Scholar]
- Ramos, O.H.P.; Kauskot, A.; Cominetti, M.R.; Bechyne, I.; Salla Pontes, C.L.; Chareyre, F.; Manent, J.; Vassy, R.; Giovannini, M.; Legrand, C.; et al. A novel αvβ3-blocking disintegrin containing the RGD motive, DisBa-01, inhibits bFGF-induced angiogenesis and melanoma metastasis. Clin. Exp. Metast. 2008, 25, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Kang, I.; Lee, Y.; Kim, D. A Novel Disintegrin Salmosin Inhibits Tumor Angiogenesis A Novel Disintegrin Salmosin Inhibits Tumor Angiogenesis 1. Cancer Res. 1999, 59, 3754–3760. [Google Scholar] [PubMed]
- Vallejo, L.F.; Rinas, U. Strategies for the recovery of active proteins through refolding of bacterial inclusion body proteins. Microb. Cell Fact. 2004, 3, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.M.; Sharma, A.; Upadhyay, A.K.; Singh, A.; Garg, L.C.; Panda, A.K. Solubilization of inclusion body proteins using n -propanol and its refolding into bioactive form. Protein Expr. Purif. 2012, 81, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Zhuravko, A.S.; Kononova, N.V.; Bobruskin, A.I. Features of the solubilization of interferon beta-1B from inclusion bodies. Russ. J. Bioorg. Chem. 2015, 41, 357–363. [Google Scholar] [CrossRef]
- Kaur, J.; Kumar, A.; Kaur, J. Strategies for optimization of heterologous protein expression in E. coli: Roadblocks and reinforcements. Int. J. Biol. Macromol. 2018, 106, 803–822. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Cui, X.; Ren, F.; Zhao, B. rAdinbitor, a novel disintegrin from Agkistrodon halys brevicaudus stejneger inhibits adhesion and proliferation of SMMC-7721 cells. Chin. Ger. J. Clin. Oncol. 2008, 7, 390–393. [Google Scholar] [CrossRef]
- Sun, M.-Z.Z.; Cui, Y.; Guo, C.; Zhao, B.; Liu, S. rAdinbitor, a disintegrin from Agkistrodon halys brevicaudus stejneger, inhibits tumorigenicity of hepatocarcinoma via enhanced anti-angiogenesis and immunocompetence. Biochimie 2015, 116, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Knight, L.C.; Romano, J.E. Functional expression of bitistatin, a disintegrin with potential use in molecular imaging of thromboembolic disease. Protein Expr. Purif. 2005, 39, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, J.H.; Silva, C.A.; Assakura, M.T.; Camargo, A.C.M.; Serrano, S.M.T. Molecular cloning, functional expression, and molecular modeling of bothrostatin, a new highly active disintegrin from Bothrops jararaca venom. Biochem. Biophys. Res. Commun. 2005, 329, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Cassini-Vieira, P.; Deconte, S.R.A.; Tomiosso, T.C.; Campos, P.P.E.; Montenegro, C.D.F.; Selistre-de-Araújo, H.S.; Barcelos, L.S.; Andrade, S.P.A.; Araújo, F.D.A. DisBa-01 inhibits angiogenesis, inflammation and fibrogenesis of sponge-induced-fibrovascular tissue in mice. Toxicon 2014, 92, 81–89. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, C.R.; de Marqueti, R.C.; Cominetti, M.R.; Douat, E.S.V.; Ribeiro, J.U.; Pontes, C.L.S.; Borghi-Silva, A.; Selistre-de-Araujo, H.S. Effects of blocking αvβ3 integrin by a recombinant RGD disintegrin on remodeling of wound healing after induction of incisional hernia in rats. Acta Cir. Bras. 2015, 30, 134–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montenegro, C.F.; Salla-Pontes, C.L.; Ribeiro, J.U.; MacHado, A.Z.; Ramos, R.F.; Figueiredo, C.C.; Morandi, V.V.V.; Selistre-De-Araujo, H.S. Blocking αvβ3 integrin by a recombinant RGD disintegrin impairs VEGF signaling in endothelial cells. Biochimie 2012, 94, 1812–1820. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, L.C.; Massimino, L.C.; Durante, A.C.; Tansini, A.; Urbaczek, A.C.; Selistre-de-Araújo, H.S.; Carlos, I.Z. Recombinant disintegrin targets α(v) β(3) integrin and leads to mediator production. Cell Adhes. Migr. 2014, 8, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Hantgan, R.R.; Stahle, M.C.; Connor, J.H.; Lyles, D.S.; Horita, D.A.; Rocco, M.; Nagaswami, C.; Weisel, J.W.; McLane, M.A. The Disintegrin Echistatin Stabilizes Integrin αIIbβ3’s Open Conformation and Promotes Its Oligomerization. J. Mol. Biol. 2004, 342, 1625–1636. [Google Scholar] [CrossRef] [PubMed]
- Hantgan, R.R.; Stahle, M.C.; Connor, J.H.; Horita, D.A.; Rocco, M.; McLane, M.A.; Yakovlev, S.; Medved, L. Integrin αIIbβ3:ligand interactions are linked to binding-site remodeling. Protein Sci. 2006, 15, 1893–1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, J.; Paquette-Straub, C.; Sage, E.H.; Funk, S.E.; Patel, V.; Galileo, D.; McLane, M.A. Inhibition of melanoma cell motility by the snake venom disintegrin eristostatin. Toxicon 2007, 49, 899–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLane, M.A.; Zhang, X.; Tian, J.; Zelinskas, C.; Srivastava, A.; Hensley, B.; Paquette-Straub, C. Scratching below the surface: Wound healing and alanine mutagenesis provide unique insights into interactions between eristostatin, platelets and melanoma cells. Pathophysiol. Haemost. Thromb. 2006, 34, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Hailey, S.; Adams, E.; Penn, R.; Wong, A.; McLane, M.A. Effect of the disintegrin eristostatin on melanoma-natural killer cell interactions. Toxicon 2013, 61, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Della-Casa, M.S.; Junqueira-de-Azevedo, I.; Butera, D.; Clissa, P.B.; Lopes, D.S.; Serrano, S.M.T.; Pimenta, D.C.; Magalhães, G.S.; Ho, P.L.; Moura-da-Silva, A.M. “Insularin, a disintegrin from Bothrops insularis venom: Inhibition of platelet aggregation and endothelial cell adhesion by the native and recombinant GST-insularin proteins”. Toxicon 2011, 57, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Soler, R.; Lorente, C.; Company, B.; Sanz, L.; Juárez, P.; Pérez, A.; Zhang, Y.; Jin, Y.; Chen, R.; Eble, J.A.; et al. Recombinant expression of mutants of the Frankenstein disintegrin, RTS-ocellatusin. Evidence for the independent origin of RGD and KTS/RTS disintegrins. Toxicon 2012, 60, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Bolás, G.; de Rezende, F.F.; Lorente, C.; Sanz, L.; Eble, J.A.; Calvete, J.J. Inhibitory effects of recombinant RTS-jerdostatin on integrin α1β1 function during adhesion, migration and proliferation of rat aortic smooth muscle cells and angiogenesis. Toxicon 2014, 79, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Lima-dos-Santos, I.; Della-Casa, M.S.; Portes-Junior, J.A.; Calabria, P.A.L.; Magalhães, G.S.; Moura-da-Silva, A.M. Characterization of Neuwiedin, a new disintegrin from Bothrops neuwiedi venom gland with distinct cysteine pattern. Toxicon 2015, 104, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.H.; Tsai, W.J.; Lo, S.J. Glutathione S-transferase-rhodostomin fusion protein inhibits platelet aggregation and induces platelet shape change. Toxicon 1997, 35, 195–204. [Google Scholar] [CrossRef]
- Chang, H.H.; Lo, S.J. Full-spreading platelets induced by the recombinant rhodostomin are via binding to integrins and correlated with FAK phosphorylation. Toxicon 1998, 36, 1087–1099. [Google Scholar] [CrossRef]
- Tselepis, V.H.; Green, L.J.; Humphries, M.J. An RGD to LDV motif conversion within the disintegrin kistrin generates an integrin antagonist that retains potency but exhibits altered receptor specificity. Evidence for a functional equivalence of acidic integrin- binding motifs. J. Biol. Chem. 1997, 272, 21341–21348. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.H.; Lin, C.H.; Lo, S.J. Recombinant rhodostomin substrates induce transformation and active calcium oscillation in human platelets. Exp. Cell Res. 1999, 250, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Suntravat, M.; Jia, Y.; Lucena, S.E.; Sánchez, E.E.; Pérez, J.C. cDNA cloning of a snake venom metalloproteinase from the eastern diamondback rattlesnake (Crotalus adamanteus), and the expression of its disintegrin domain with anti-platelet effects. Toxicon 2013, 64, 43–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suntravat, M.; Barret, H.S.; Jurica, C.A.; Lucena, S.E.; Perez, J.C. Recombinant disintegrin (r-Cam-dis) from Crotalus adamanteus inhibits adhesion of human pancreatic cancer cell lines to laminin-1 and vitronectin. J. Venom Res. 2015, 6, 1–10. [Google Scholar] [PubMed]
- Suntravat, M.; Helmke, T.J.; Atphaisit, C.; Cuevas, E.; Lucena, S.E.; Uzcátegui, N.L.; Sánchez, E.E.; Rodriguez-Acosta, A. Expression, purification, and analysis of three recombinant ECD disintegrins (r-colombistatins) from P-III class snake venom metalloproteinases affecting platelet aggregation and SK-MEL-28 cell adhesion. Toxicon 2016, 122, 43–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seoane, A.I.; Tran, V.L.; Sanchez, E.E.; White, S.A.; Choi, J.L.; Gaytán, B.; Chavez, N.; Reyes, S.R.; Ramos, C.J.; Tran, L.H.; et al. The mojastin mutant Moj-DM induces apoptosis of the human melanoma SK-Mel-28, but not the mutant Moj-NN nor the non-mutated recombinant Moj-WN. Toxicon 2010, 56, 391–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucena, S.; Sanchez, E.E.; Perez, J.C. Anti-metastatic activity of the recombinant disintegrin, r-mojastin 1, from the Mohave rattlesnake. Toxicon 2011, 57, 794–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucena, S.; Castro, R.; Lundin, C.; Hofstetter, A.; Alaniz, A.; Suntravat, M.; Sánchez, E.E. Inhibition of pancreatic tumoral cells by snake venom disintegrins. Toxicon 2015, 93, 136–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos, C.J.; Gutierrez, D.A.; Aranda, A.S.; Koshlaychuk, M.A.; Carrillo, D.A.; Medrano, R.; McBride, T.D.; U, A.; Medina, S.M.; Lombardo, M.C.; et al. Functional characterization of six aspartate (D) recombinant mojastin mutants (r-Moj): A second aspartate amino acid carboxyl to the RGD in r-Moj-D_ peptides is not sufficient to induce apoptosis of SK-Mel-28 cells. Toxicon 2016, 118, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Lucena, S.E.; Jia, Y.; Soto, J.G.; Parral, J.; Cantu, E.; Brannon, J.; Lardner, K.; Ramos, C.J.; Seoane, A.I.; Sánchez, E.E. Anti-invasive and anti-adhesive activities of a recombinant disintegrin, r-viridistatin 2, derived from the Prairie rattlesnake (Crotalus viridis viridis). Toxicon 2012, 60, 31–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, I.C.; Kim, D.S.; Jang, Y.; Chung, K.H. Suppressive mechanism of salmosin, a novel disintegrin in B16 melanoma cell metastasis. Biochem. Biophys. Res. Commun. 2000, 275, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Minea, R.; Costa, F.; Chen, C.; Markland, F.S.; Swenson, S.; Costa, F.; Chen, T.C.; Markland, F.S. Development of a novel recombinant disintegrin, contortrostatin, as an effective anti-tumor and anti-angiogenic agent. Pathophysiol. Haemost. Thromb. 2005, 34, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Minea, R.; Helchowski, C.; Rubino, B.; Brodmann, K.; Swenson, S.; Markland, F. Development of a chimeric recombinant disintegrin as a cost-effective anti-cancer agent with promising translational potential. Toxicon 2012, 59, 472–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadokura, H.; Beckwith, J. Mechanisms of Oxidative Protein Folding in the Bacterial Cell Envelope. Antioxid. Redox Signal. 2010, 13, 1231–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Li, G.; Ren, X.; Herrler, G. Select what you need: A comparative evaluation of the advantages and limitations of frequently used expression systems for foreign genes. J. Biotechnol. 2007, 127, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, A.; Neubauer, P.; Myllyharju, J. High-level production of human collagen prolyl 4-hydroxylase in Escherichia coli. Matrix Biol. 2005, 24, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Kovalskaya, N.; Hammond, R.W. Expression and functional characterization of the plant antimicrobial snakin-1 and defensin recombinant proteins. Protein Expr. Purif. 2009, 63, 12–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgiou, G.; Telford, J.N.; Shuler, M.L.; Wilson, D.B. Localization of inclusion bodies in Escherichia coli overproducing beta-lactamase or alkaline phosphatase. Appl. Environ. Microbiol. 1986, 52, 1157–1161. [Google Scholar] [PubMed]
- Majander, K.; Anton, L.; Antikainen, J.; Lång, H.; Brummer, M.; Korhonen, T.K.; Westerlund-Wikström, B. Extracellular secretion of polypeptides using a modified Escherichia coli flagellar secretion apparatus. Nat. Biotechnol. 2005, 23, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.-G.; Xia, X.-X.; Choi, J.H.; Lee, S.Y. Proteome-based identification of fusion partner for high-level extracellular production of recombinant proteins in Escherichia coli. Biotechnol. Bioeng. 2008, 101, 587–601. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Wang, S.; Li, H.; Yu, H.; Qi, Q. Identification of a heterologous cellulase and its N-terminus that can guide recombinant proteins out of Escherichia coli. Microb. Cell Fact. 2015, 14, 49. [Google Scholar] [CrossRef] [PubMed]
- Schmoldt, H.-U.; Wentzel, A.; Becker, S.; Kolmar, H. A fusion protein system for the recombinant production of short disulfide bond rich cystine knot peptides using barnase as a purification handle. Protein Expr. Purif. 2005, 39, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Wan, E.W.M.; Baneyx, F. TolAIII co-overexpression facilitates the recovery of periplasmic recombinant proteins into the growth medium of Escherichia coli. Protein Expr. Purif. 1998. [Google Scholar] [CrossRef] [PubMed]
- Lobstein, J.; Emrich, C.A.; Jeans, C.; Faulkner, M.; Riggs, P.; Berkmen, M. SHuffle, a novel Escherichia coli protein expression strain capable of correctly folding disulfide bonded proteins in its cytoplasm. Microb. Cell Fact. 2012, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Marblestone, J.G. Comparison of SUMO fusion technology with traditional gene fusion systems: Enhanced expression and solubility with SUMO. Protein Sci. 2006, 15, 182–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butt, T.R.; Edavettal, S.C.; Hall, J.P.; Mattern, M.R. SUMO fusion technology for difficult-to-express proteins. Protein Expr. Purif. 2005, 43, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Shimokawa-Falcão, L.; Caporrino, M.; Barbaro, K.; Della-Casa, M.; Magalhães, G. Toxin Fused with SUMO Tag: A New Expression Vector Strategy to Obtain Recombinant Venom Toxins with Easy Tag Removal inside the Bacteria. Toxins 2017, 9, 82. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Hirz, M.; Pichler, H.; Schwab, H. Protein expression in Pichia pastoris: Recent achievements and perspectives for heterologous protein production. Appl. Microbiol. Biotechnol. 2014, 98, 5301–5317. [Google Scholar] [CrossRef] [PubMed]
- Cregg, J.M.; Cereghino, J.L.; Shi, J.; Higgins, D.R. Recombinant Protein Expression in Pichia pastoris. Mol. Biotechnol. 2000. [Google Scholar] [CrossRef]
- Daly, R.; Hearn, M.T.W. Expression of heterologous proteins in Pichia pastoris: A useful experimental tool in protein engineenring and production. J. Mol. Recognit. 2005, 18, 119–138. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.-Y.; Sohn, Y.-D.; Chung, K.-H.; Kim, D.-S. Structural and functional significance of disulfide bonds in saxatilin, a 7.7 kDa disintegrin. Biochem. Biophys. Res. Commun. 2002, 293, 530–536. [Google Scholar] [CrossRef]
- Hong, S.Y.; Lee, H.; You, W.K.; Chung, K.H.; Kim, D.S.; Song, K. The snake venom disintegrin salmosin induces apoptosis by disassembly of focal adhesions in bovine capillary endothelial cells. Biochem. Biophys. Res. Commun. 2003, 302, 502–508. [Google Scholar] [CrossRef]
- Higuchi, D.A.; Almeida, M.C.; Barros, C.C.; Sanchez, E.F.; Pesquero, P.R.; Lang, E.A.S.; Samaan, M.; Araujo, R.C.; Pesquero, J.B.; Pesquero, J.L. Leucurogin, a new recombinant disintegrin cloned from Bothrops leucurus (white-tailed-jararaca) with potent activity upon platelet aggregation and tumor growth. Toxicon 2011, 58, 123–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singhamatr, P.; Rojnuckarin, P. Molecular cloning of albolatin, a novel snake venom metalloprotease from green pit viper (Trimeresurus albolabris), and expression of its disintegrin domain. Toxicon 2007, 50, 1192–1200. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-T.T.; Tang, C.-H.H.; Chuang, W.-J.J.; Wang, S.-M.M.; Huang, T.-F.F.; Fu, W.-M.M. Inhibition of adipogenesis by RGD-dependent disintegrin. Biochem. Pharmacol. 2005, 70, 1469–1478. [Google Scholar] [CrossRef] [PubMed]
- Dennis, M.S.; Carter, P.; Lazarus, R.A. Binding Interactions of Kistrin With Platelet Glycoprotein IIb-IIIa: Analysus by Site-Directed Mutagenesis. Proteins Struct. Funct. Bioinform. 1993, 15, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-C.; Chuang, W.-J.; Chung, C.-H.; Chang, C.-H.; Peng, H.-C.; Huang, T.-F. Snake Venom Disintegrin Inhibits the Activation of Toll-Like Receptors and Alleviates Sepsis through Integrin alphaVbeta3 Blockade. Sci. Rep. 2016, 6, 23387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.-T.; Shiu, J.; Huang, C.; Chen, Y.-C.; Chen, C.; Chang, Y.; Chuang, W. Effects of the RGD loop and C-terminus of rhodostomin on regulating integrin αIIbβ3 recognition. PLoS ONE 2017, 12, e0175321. [Google Scholar] [CrossRef] [PubMed]
- Sohn, Y.D.; Hong, S.Y.; Cho, K.S.; Choi, W.S.; Song, S.W.; Bae, J.S.; Kim, D.S.; Chung, K.H. Acute and repeated dose toxicity studies of recombinant saxatilin, a disintegrin from the Korean snake (Gloydius saxatilis). Toxicon 2008, 51, 406–417. [Google Scholar] [CrossRef] [PubMed]
- Sohn, Y.D.; Cho, K.S.; Sun, S.A.; Sung, H.J.; Kwak, K.W.; Hong, S.Y.; Kim, D.S.; Chung, K.H. Suppressive effect and mechanism of saxatilin, a disintegrin from Korean snake (Gloydius saxatilis), in vascular smooth muscle cells. Toxicon 2008, 52, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Kwon, I.; Hong, S.Y.; Kim, Y.D.; Nam, H.S.; Kang, S.; Yang, S.H.; Heo, J.H. Thrombolytic effects of the snake venom disintegrin saxatilin determined by novel assessment methods: A FeCl3-induced thrombosis model in mice. PLoS ONE 2013, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Setlow, J.K. Genetic Engineering; Genetic Engineering: Principles and Methods (Book 27); Setlow, J.K., Ed.; Springer: Boston, MA, USA, 2002; Volume 27, ISBN 978-1-4613-5210-5. [Google Scholar]
- Chen, C.-Y.; Shiu, J.-H.; Hsieh, Y.-H.; Liu, Y.-C.; Chen, Y.-C.; Jeng, W.-Y.; Tang, M.-J.; Lo, S.J.; Chuang, W.-J. Effect of D to E mutation of the RGD motif in rhodostomin on its activity, structure, and dynamics: Importance of the interactions between the D residue and integrin. Proteins 2009, 76, 808–821. [Google Scholar] [CrossRef] [PubMed]
- Shiu, J.-H.; Chen, C.-Y.; Chen, Y.-C.; Chang, Y.-T.; Huang, C.-H.; Chuang, W.-J. Effect of P to A mutation of the N-terminal residue adjacent to the Rgd motif on rhodostomin: Importance of dynamics in integrin recognition. PLoS ONE 2012, 7, e28833. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, D.A.; Aranda, A.S.; Carrillo, D.A.R.; Koshlaychuk, M.A.; Sanchez, E.E.; Lucena, S.E.; Soto, J.G.; Marceau, K.; Ruttle, P.L.; Shirtcliff, E.A.; et al. Functional analysis of four single (RGDWL, RGDWM, RGDWP, RGDMN) and two double (RGDNM, RGDMP) mutants: The importance of methionine (M) in the functional potency of recombinant mojastin (r-Moj). Toxicon 2015, 57, 742–768. [Google Scholar] [CrossRef] [PubMed]
- Mezö, G.; Hudecz, F. Synthesis of Linear, Branched, and Cyclic Peptide Chimera. In Peptide Synthesis and Applications; Humana Press: Totowa, NJ, USA, 2005; Volume 298, pp. 63–76. ISBN 978-1-58829-317-6. [Google Scholar]
- Jing, J.; Lu, S. Inhibition of Platelet Aggregation of a Mutant Proinsulin Chimera Engineered by Introduction of a Native Lys-Gly-Asp-containing Sequence. Biotechnol. Lett. 2005, 27, 1259–1265. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.E.; Galán, J.A.; Russell, W.K.; Soto, J.G.; Russell, D.H.; Pérez, J.C. Isolation and characterization of two disintegrins inhibiting ADP-induced human platelet aggregation from the venom of Crotalus scutulatus scutulatus (Mohave Rattlesnake). Toxicol. Appl. Pharmacol. 2006, 212, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Clarke, E.A.; Trikha, M.; Markland, M.S.; Bugge, J.S. Structurally distinct disintegrins contortrostatin and multiquamatin differentially regulate platelet tyrosine phosphorylation. J. Biol. Chem. 1994, 269, 21940–21943. [Google Scholar]
- Scarborough, R.M.; Naughton, M.A.; Teng, W.; Rose, J.W.; Phillips, D.R.; Nannizzi, L.; Arfsten, A.; Campbell, A.M.; Charo, I.F. Design of potent and specific integrin antagonists: Peptide antagonists with high specificity for glycoprotein IIb-IIIa. J. Biol. Chem. 1993, 268, 1066–1073. [Google Scholar] [PubMed]
- Lynch, J.J.; Cook, J.J.; Sitko, G.R.; Holahan, M.A.; Ramjit, D.R.; Mellott, M.J.; Stranieri, M.T.; Stabilito, I.I.; Zhang, G.; Lynch, R.J. Nonpeptide glycoprotein IIb/IIIa inhibitors. 5. Antithrombotic effects of MK-0383. J. Pharmacol. Exp. Ther. 1995, 272, 20–32. [Google Scholar] [PubMed]
- Lin, E.; Wang, Q.; Swenson, S.; Jadvar, H.; Groshen, S.; Ye, W.; Markland, F.S.; Pinski, J. The disintegrin contortrostatin in combination with docetaxel is a potent inhibitor of prostate cancer in vitro and in vivo. Prostate 2010, 70, 1359–1370. [Google Scholar] [CrossRef] [PubMed]
- Ritter, M.R.; Zhou, Q.; Markland, F.S. Contortrostatin, a Homodimeric Disintegrin, Actively Disrupts Focal Adhesion and Cytoskeletal Structure and Inhibits Cell Motility Through a Novel Mechanism. Cell Commun. Adhes. 2001, 8, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Sherwin, R.P.; Parrish, C.; Richters, V.; Groshen, S.G.; Tsao-Wei, D.; Markland, F.S. Contortrostatin, a dimeric disintegrin from Agkistrodon contortrix contortrix, inhibits breast cancer progression. Breast Cancer Res. Treat. 2000, 61, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Swenson, S.; Ramu, S.; Markland, F.S. Anti-angiogenesis and RGD-containing snake venom disintegrins. Curr. Pharm. Des. 2007, 13, 2860–2871. [Google Scholar] [CrossRef] [PubMed]
- Marcinkiewicz, C.; Weinreb, P.H.; Calvete, J.J.; Kisiel, D.G.; Mousa, S.A.; Tuszynski, G.P.; Lobb, R.R. Obtustatin: A potent selective inhibitor of alpha1beta1 integrin in vitro and angiogenesis in vivo. Cancer Res. 2003, 63, 2020–2023. [Google Scholar] [PubMed]
- Staniszewska, I.; Walsh, E.M.; Rothman, V.L.; Gaathon, A.; Tuszynski, G.P.; Calvete, J.J.; Lazarovici, P.; Marcinkiewicz, C. Effect of VP12 and viperistatin on inhibition of collagen receptors-dependent melanoma metastasis. Cancer Biol. Ther. 2009, 8, 1507–1516. [Google Scholar] [CrossRef] [PubMed]
- Momic, T.; Katzehendler, J.; Benny, O.; Lahiani, A.; Cohen, G.; Noy, E.; Senderowitz, H.; Eble, J.A.; Marcinkiewicz, C.; Lazarovici, P. Vimocin and Vidapin, cyclic KTS peptides, dual antagonists of α1β1/α2β1 integrins with antiangiogenic activity. J. Pharmacol. Exp. Ther. 2014, 506–519. [Google Scholar] [CrossRef] [PubMed]
- Lazarovici, P.; Momic, T.; Katzehendler, J.; Shai, E.; Noy, E.; Senderowitz, H.; Eble, J.A.; Marcinkiewicz, C.; Varon, D. Vipegitide: A folded peptidomimetic partial antagonist of α2β1 integrin with antiplatelet aggregation activity. Drug Des. Devel. Ther. 2015, 9, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Coelho, A.L.J.; De Freitas, M.S.; Mariano-Oliveira, A.; Oliveira-Carvalho, A.L.; Zingali, R.B.; Barja-Fidalgo, C. Interaction of disintegrins with human neutrophils induces cytoskeleton reorganization, focal adhesion kinase activation, and extracellular-regulated kinase-2 nuclear translocation, interfering with the chemotactic function. FASEB J. 2001, 15, 1643–1645. [Google Scholar] [CrossRef] [PubMed]
- Coelho, A.L.J.; De Freitas, M.S.; Mariano-Oliveira, A.; Rapozo, D.C.M.; Pinto, L.F.R.; Niewiarowski, S.; Zingali, R.B.; Marcinkiewicz, C.; Barja-Fidalgo, C. RGD- and MLD-disintegrins, jarastatin and EC3, activate integrin-mediated signaling modulating the human neutrophils chemotaxis, apoptosis and IL-8 gene expression. Exp. Cell Res. 2004, 292, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Neto, E.H.; Coelho, A.L.J.; Sampaio, A.L.F.; Henriques, M.d.G.M.O.; Marcinkiewicz, C.; De Freitas, M.S.; Barja-Fidalgo, C. Activation of human T lymphocytes via integrin signaling induced by RGD-disintegrins. Biochim. Biophys. Acta 2007, 1773, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Saldanha-Gama, R.F.; Moraes, J.A.; Mariano-Oliveira, A.; Coelho, A.L.; Walsh, E.M.; Marcinkiewicz, C.; Barja-Fidalgo, C. alpha(9)beta(1) integrin engagement inhibits neutrophil spontaneous apoptosis: Involvement of Bcl-2 family members. Biochim. Biophys. Acta 2010, 1803, 848–857. [Google Scholar] [CrossRef] [PubMed]
- Moraes, J.A.; Frony, A.C.; Dias, A.M.; Renovato-Martins, M.; Rodrigues, G.; Marcinkiewicz, C.; Assreuy, J.; Barja-Fidalgo, C. Alpha1beta1 and integrin-linked kinase interact and modulate angiotensin II effects in vascular smooth muscle cells. Atherosclerosis 2015, 243, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Moraes, J.A.; Frony, A.C.; Dias, A.M.; Renovato-Martins, M.; Rodrigues, G.; Marcinkiewicz, C.; Assreuy, J.; Barja-Fidalgo, C. Data in support of alpha1beta1 and integrin-linked kinase interact and modulate angiotensin II effects in vascular smooth muscle cells. Data Br. 2016, 6, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.C.; Chuang, W.J.; Chang, C.H.; Tseng, Y.L.; Peng, H.C.; Huang, T.F. Improvements in endotoxemic syndromes using a disintegrin, rhodostomin, through integrin αvβ3-dependent pathway. J. Thromb. Haemost. 2011, 9, 593–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.I.; Kim, K.S.; Kim, H.S.; Kim, D.S.; Jang, Y.; Chung, K.H.; Park, Y.S. Inhibitory effect of the salmosin gene transferred by cationic liposomes on the progression of B16BL6 tumors. Cancer Res. 2003, 63, 6458–6462. [Google Scholar] [PubMed]
- Swenson, S.; Costa, F.; Minea, R.; Sherwin, R.P.; Ernst, W.; Fujii, G.; Yang, D.; Markland, F.S. Intravenous liposomal delivery of the snake venom disintegrin contortrostatin limits breast cancer progression. Mol. Cancer Ther. 2004, 3, 499–511. [Google Scholar] [PubMed]
- Janib, S.M.; Gustafson, J.A.; Minea, R.O.; Swenson, S.D.; Liu, S.; Pastuszka, M.K.; Lock, L.L.; Cui, H.; Markland, F.S.; Conti, P.S.; et al. Multimeric disintegrin protein polymer fusions that target tumor vasculature. Biomacromolecules 2014, 15, 2347–2358. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Disintegrin | Motif | Snake Venom ** | Plasmid | Origin of Gene | Expressed | Assay with Fusion Protein or Tag | Yield (mg/L) | Integrin | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| Acocostatin | ECD | A. c. contortrix | pGEX-KG | cDNA | E. coli BL21 | GST | NM | NE | [62] |
| Adinbitor | RGD | A. h. b. stejneger | pET23b | cDNA | E. coli BL21 | His-Tag | NM | NE | [51,77,78] |
| Bitistatin | RGD | B. arietans | pGEX-KT, pET-5a | S | E. coli BL21 | GST, no | 1–2 (free) 12 ± 3 (free) | αIIbβ3 * | [79] |
| Bothrostatin | RGD | B. jararaca | pGEX-4T | cDNA | E. coli BL21 | GST, no | 6 (free) | NE | [80] |
| DisBa-01 | RGD | B. alternatus | pET28a | cDNA | E. coli BL21 | His-Tag | NM | αIIbβ3, αvβ3 | [57,71,81,82,83,84] |
| Echistatin and mutants | RGD | E. carinatus | pGEX-KG | S | E. coli BL21 | No | NM | αIIbβ3, αvβ3 *, α5β1 * | [42,85,86] |
| Elegantin and mutant | RGD | T. elegans | pGEX-3X | S | E. coli (DE3) pLysS | GST | NM | αIIbβ3, α5β1 | [56] |
| Eristostatin and mutants | RGD | E. macmahoni | pGEX-KG, pET 39b | S | E. coli BL21 | No | NM | αIIbβ3 *, αvβ3 *, α5β1 * | [42,87,88,89] |
| Insularin | RGD | B. insularis | pGEX-4-T1 | cDNA | E. coli BL21 | GST | 50 (fused) | αIIbβ3 *, αvβ3 * | [90] |
| Jerdostatin and mutant | RTS | P. jerdonii | pET32a | cDNA | E. coli Origami B, E. coli BL21 | No | 0.5–2 | α1β1 | [61,91,92] |
| Neuwiedin | RGD | B. neuwiedi | pMST3 | cDNA | E. coli C43 | No | 8 | αIIbβ3 *, αvβ3 * | [93] |
| Obtustatin and mutant | KTS | V. l. obtusa | pGEX-4-T1 | S | E. coli BL21 | No | NM | α1β1 | [60] |
| Rhodostomin and mutant | RGD | C. rhodostoma | pGEX-2KS, pGEX-2T | S cDNA | E. coli K38, E. coli RR1, E. coli DH5α | GST | NM | αIIbβ3, αvβ3, α4β1 α5β1 | [55,94,95,96,97] |
| Rubistatin | MVD | C. r. ruber | pET32b | cDNA | E. coli origami2 | No | NM | NE | [63] |
| r-Cam-dis | RGD | C. adamanteus | pET-43.1a, pGEX-4T-1 | cDNA | E. coli BL21 | No | 1 | αIIbβ3, αvβ3, αvβ5, αvβ6, α2β1, α6β1 | [98,99] |
| r-colombis-tatin | ECD | B. colombiensis | pGEX-4-T1 | cDNA | E. coli BL21 | No | NM | NE | [100] |
| r-mojastin 1 and mutant | RGD | C. s. scutulatus | pGEX-4-T1, pGEX-KG | cDNA | E. coli BL21 | GST, no | 0.8 (free) | αIIbβ3 *, αv *, αvβ3 *, αvβ5 *, α3β1 *, α6 *, β1 *, β3 * | [37,58,101,102,103,104] |
| r-ocellatusin and mutant | RGD RTS | E. ocellatus | pET32a | cDNA | E. coli BL21 | No | 0.5–1 | NE | [91] |
| r-viridistatin 2 | RGD | C. v. viridis | pGEX-4-T1 | cDNA | E. coli BL21 | No | NM | αvβ3, αvβ5, α3β1 | [37,103,105] |
| Salmosin 1 | RGD | A. h. brevicandus | pMA-PRK153, ΔpMA | cDNA | E. coli MC1061 | PRK | NM | αIIbβ3, αvβ3 | [59,72,106] |
| Vicrostatin | RGD | chimeric recombinant | pET32a | NM | Origami B (DE3) pLysS | No | 20 | αvβ3, αvβ5, α5β1 | [107,108] |
| Disintegrin | Motif | Snake Venom ** | Plasmid | Origin of Gene | Expressed | Assay with Fusion Protein or Tag | Yield (mg/L) | Integrin | References |
|---|---|---|---|---|---|---|---|---|---|
| Albolatin | KGD | T. albolabris | pPICZα A | cDNA | P. pastoris X33 | His-Tag | 3.3 | NE | [129] |
| Echistatin (Ech) | RGD | Echis carinatus | pPICZα A | S | P. pastoris X33 | His-Tag | 2–7 | αIIbβ3 | [44] |
| Leucurogin | ECD | Bothrops leucurus | pPIC9 | cDNA | P. pastoris X33 | No | NM | NE | [127] |
| Rhodostomin mutant | RGD | Calloselasma rhodostoma | pPICZαA | NM | P. pastoris X33 | No | NM | NE | [130] |
| Rhodostomin and mutant | RGD | Calloselasma rhodostoma | pPICZαA | S NM | P. pastoris X33 | No | 10–25 | αIIbβ3, α5β1 αvβ3 | [44,131,132,133] |
| Salmosin 1 | RGD | Agkistrodon h. brevicandus | pPIC9 | cDNA | P. pastoris | No | NM | αvβ3 | [127] |
| Saxatilin and mutant | RGD | Gloydius saxatilis | pPIC9 | cDNA | P. pastoris GS115 | No | 150 | αIIbβ3, αvβ3 | [126,134,135,136] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
David, V.; Succar, B.B.; De Moraes, J.A.; Saldanha-Gama, R.F.G.; Barja-Fidalgo, C.; Zingali, R.B. Recombinant and Chimeric Disintegrins in Preclinical Research. Toxins 2018, 10, 321. https://doi.org/10.3390/toxins10080321
David V, Succar BB, De Moraes JA, Saldanha-Gama RFG, Barja-Fidalgo C, Zingali RB. Recombinant and Chimeric Disintegrins in Preclinical Research. Toxins. 2018; 10(8):321. https://doi.org/10.3390/toxins10080321
Chicago/Turabian StyleDavid, Victor, Barbara Barbosa Succar, João Alfredo De Moraes, Roberta Ferreira Gomes Saldanha-Gama, Christina Barja-Fidalgo, and Russolina Benedeta Zingali. 2018. "Recombinant and Chimeric Disintegrins in Preclinical Research" Toxins 10, no. 8: 321. https://doi.org/10.3390/toxins10080321
APA StyleDavid, V., Succar, B. B., De Moraes, J. A., Saldanha-Gama, R. F. G., Barja-Fidalgo, C., & Zingali, R. B. (2018). Recombinant and Chimeric Disintegrins in Preclinical Research. Toxins, 10(8), 321. https://doi.org/10.3390/toxins10080321