Oxidative Damage and Nrf2 Translocation Induced by Toxicities of Deoxynivalenol on the Placental and Embryo on Gestation Day 12.5 d and 18.5 d

Abstract
:1. Introduction
2. Results
2.1. Effect of DON Exposure on Body Weight
2.2. Effect of DON Exposure on Embryonic Survival, Growth, and Development on GD 12.5 d and GD 18.5 d Embryos
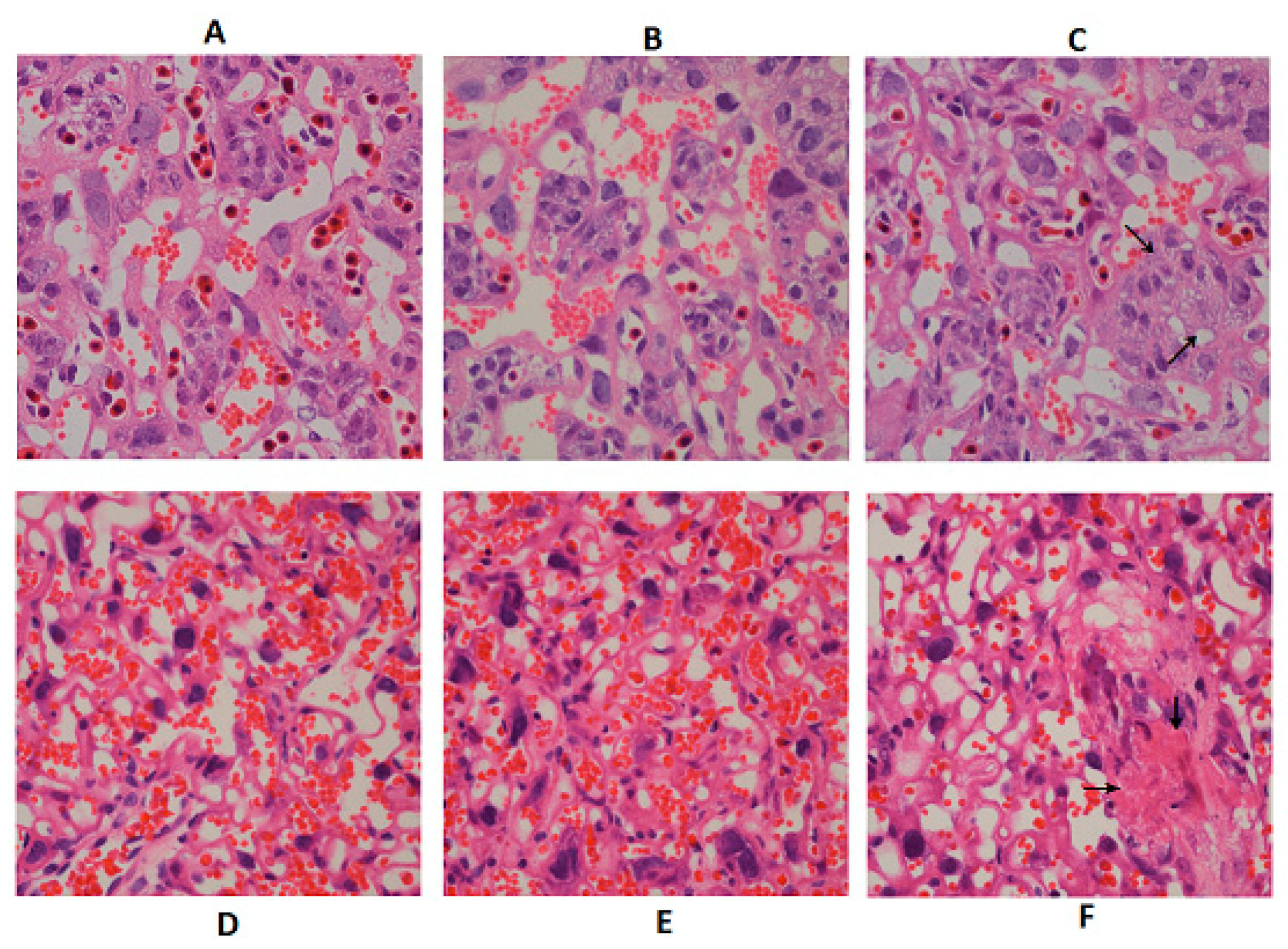
2.3. Effect of DON Exposure on Placenta Histopathological and Function Changes in GD 12.5 d and GD 18.5 d Maternal Mice
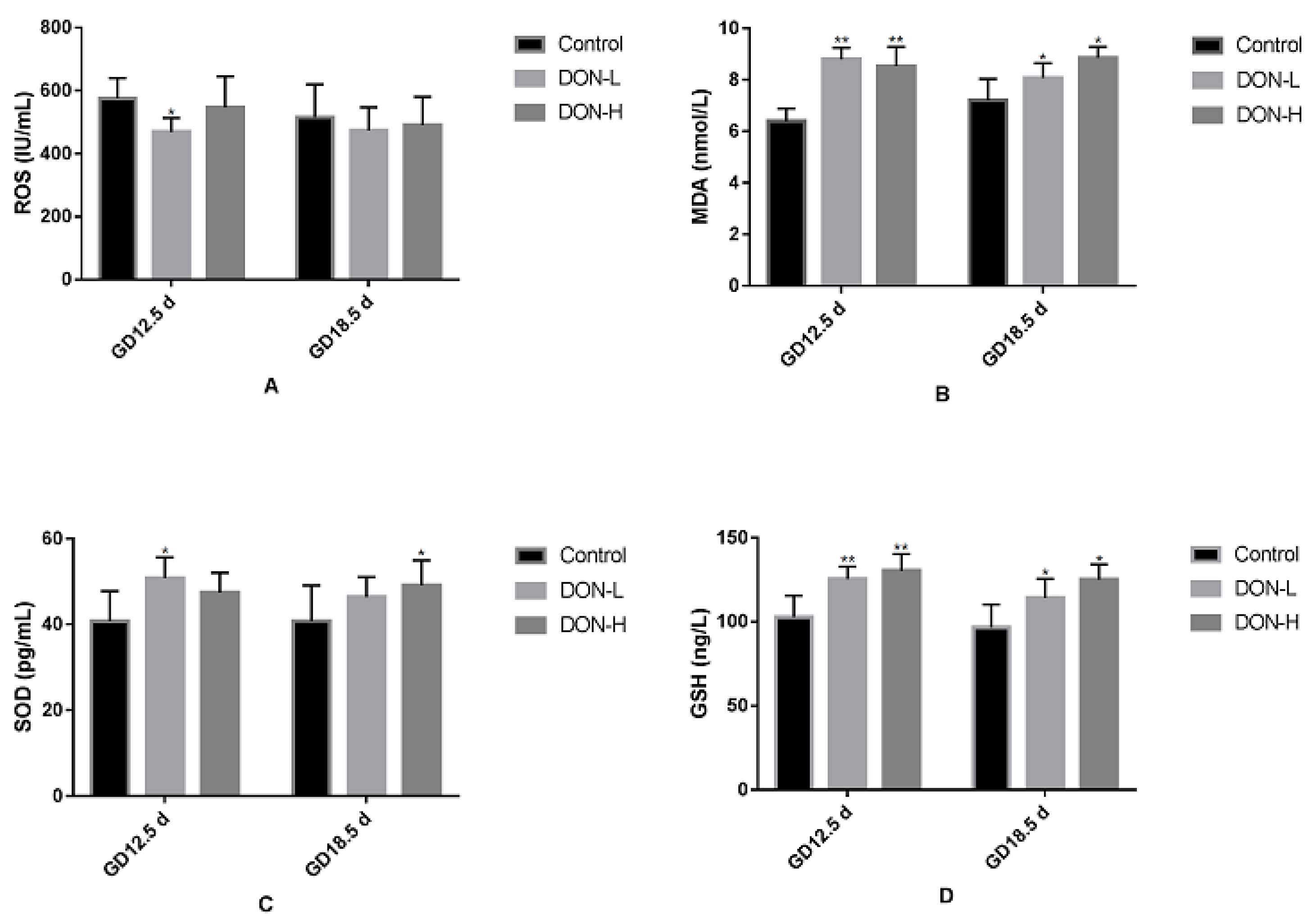
2.4. Effect of DON Exposure on the Levels of Oxidant and Antioxidant Biomarkers in the Placenta of GD 12.5 d and GD 18.5 d Maternal Mice
2.5. Effect of DON Exposure on the HO-1 Level in Maternal Placenta on GD 12.5 d and GD 18.5 d
2.6. Effect of DON Exposure on the Intracellular Translocation of Nrf2 in the Placenta of GD 12.5 d and GD 18.5 d Maternal Mice
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals and Treatment
4.3. Study Design
4.4. β-CG, ROS, MDA, SOD, GSH, and HO-1 in the Placenta
4.5. Placental Pathological Examination and Immunostaining
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| β-CG | β-chorionic gonadotropin |
| β-HCG | β-human chorionic gonadotropin |
| DON | Deoxynivalenol |
| DON-H | DON high-dose group |
| DON-L | DON low-dose group |
| GD | Gestation day |
| GSH | Glutathione |
| H&E | Hematoxylin and eosin |
| HO-1 | Heme oxygenase-1 |
| MDA | Malondialdehyde |
| MFI | Mean fluorescence intensity |
| Nrf2 | NF-E2-related factor 2 |
| ROS | Reactive oxygen species |
| SPF | Specific Pathogen Free |
| SOD | Superoxide Dismutase |
References
- Wu, Q.; Wang, X.; Nepovimova, E.; Wang, Y.; Yang, H.; Li, L.; Zhang, X.; Kuca, K. Antioxidant agents against trichothecenes: New hints for oxidative stress treatment. Oncotarget 2017, 8, 110708–110726. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Peng, Z.; Chen, L.; Nussler, A.K.; Liu, L.; Yang, W. Deoxynivalenol, gut microbiota and immunotoxicity: A potential approach? Food Chem. Toxicol. 2018, 112, 342–354. [Google Scholar] [CrossRef] [PubMed]
- Joint FAO/WHO Expert Committee on Food Additives. Evaluation of Certain Mycotoxins in Food: Fifty-Sixth Report of the Joint FAO/WHO Expert Committee on Food Additives; WHO: Geneva, Switzerland, 2002; pp. 1–62. [Google Scholar]
- Malachova, A.; Dzuman, Z.; Veprikova, Z.; Vaclavikova, M.; Zachariasova, M.; Hajslova, J. Deoxynivalenol, deoxynivalenol-3-glucoside, and enniatins: The major mycotoxins found in cereal-based products on the czech market. J. Agric. Food Chem. 2011, 59, 12990–12997. [Google Scholar] [CrossRef] [PubMed]
- Pleadin, J.; Zadravec, M.; Persi, N.; Vulic, A.; Jaki, V.; Mitak, M. Mould and mycotoxin contamination of pig feed in northwest croatia. Mycotoxin Res. 2012, 28, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Tran, S.-T.; Smith, T.K.; Girgis, G.N. A survey of free and conjugated deoxynivalenol in the 2008 corn crop in ontario, canada. J. Sci. Food Agric. 2012, 92, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Chen, L.; Peng, Z.; Wang, D.; Song, Y.; Wang, H.; Yao, P.; Yan, H.; Nussler, A.K.; Liu, L.; et al. Embryotoxicity caused by DON-induced oxidative stress mediated by Nrf2/HO-1 pathway. Toxins 2017, 9, 188. [Google Scholar] [CrossRef] [PubMed]
- Sobrova, P.; Adam, V.; Vasatkova, A.; Beklova, M.; Zeman, L.; Kizek, R. Deoxynivalenol and its toxicity. Interdiscip. Toxicol. 2010, 3, 94–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khera, K.S.; Whalen, C.; Angers, G.; Vesonder, R.F.; Kuiper-Goodman, T. Embryotoxicity of 4-deoxynivalenol (vomitoxin) in mice. Bull. Environ. Contam. Toxicol. 1982, 29, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Chen, L.; Peng, Z.; Nussler, A.K.; Wu, Q.; Liu, L.; Yang, W. Mechanism of deoxynivalenol effects on the reproductive system and fetus malformation: Current status and future challenges. Toxicol. In Vitro 2017, 41, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Debouck, C.; Haubruge, E.; Bollaerts, P.; van Bignoot, D.; Brostaux, Y.; Werry, A.; Rooze, M. Skeletal deformities induced by the intraperitoneal administration of deoxynivalenol (vomitoxin) in mice. Int. Orthop. 2001, 25, 194–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, T.F.; Sprando, R.L.; Black, T.N.; Olejnik, N.; Eppley, R.M.; Hines, F.A.; Rorie, J.; Ruggles, D.I. Effects of deoxynivalenol (DON, vomitoxin) on in utero development in rats. Food Chem. Toxicol. 2006, 44, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Al-Gubory, K.H.; Fowler, P.A.; Garrel, C. The roles of cellular reactive oxygen species, oxidative stress and antioxidants in pregnancy outcomes. Int. J. Biochem. Cell Biol. 2010, 42, 1634–1650. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.-H.; Wang, X.; Yang, W.; Nussler, A.K.; Xiong, L.-Y.; Kuca, K.; Dohnal, V.; Zhang, X.-J.; Yuan, Z.-H. Oxidative stress-mediated cytotoxicity and metabolism of T-2 toxin and deoxynivalenol in animals and humans: An update. Arch. Toxicol. 2014, 88, 1309–1326. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.C.; Martel, F. Oxidative stress in pregnancy and fertility pathologies. Cell Biol. Toxicol. 2014, 30, 301–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keyse, S.M.; Tyrrell, R.M. Heme oxygenase is the major 32-kDa stress protein induced in human skin fibroblasts by UVA radiation, hydrogen peroxide, and sodium arsenite. Proc. Natl. Acad. Sci. USA 1989, 86, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Foresti, R.; Clark, J.E.; Green, C.J.; Motterlini, R. Thiol compounds interact with nitric oxide in regulating heme oxygenase-1 induction in endothelial cells. Involvement of superoxide and peroxynitrite anions. J. Biol. Chem. 1997, 272, 18411–18417. [Google Scholar] [CrossRef] [PubMed]
- Baker, H.M.; Anderson, B.F.; Baker, E.N. Dealing with iron: Common structural principles in proteins that transport iron and heme. Proc. Natl. Acad. Sci. USA 2003, 100, 3579–3583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonen, A.L. The Keap1-Nrf2 pathway: Mechanisms of activation and dysregulation in cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Mitsuishi, Y.; Motohashi, H.; Yamamoto, M. The Keap1-Nrf2 system in cancers: Stress response and anabolic metabolism. Front. Oncol. 2012, 2, 200. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.C.; Nguyen, T.; Pickett, C.B. Phosphorylation of Nrf2 at Ser-40 by protein kinase C regulates antioxidant response element-mediated transcription. J. Biol. Chem. 2002, 277, 42769–42774. [Google Scholar] [CrossRef] [PubMed]
- Katika, M.R.; Hendriksen, P.J.; van Loveren, H.; Peijnenburg, A.A.C.M. Characterization of the modes of action of deoxynivalenol (DON) in the human Jurkat T-cell line. J. Immunotoxicol. 2015, 12, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Butler, S.A.; Iles, R.K. The free monomeric beta subunit of human chorionic gonadotrophin (hCGβ) and the recently identified homodimeric beta-beta subunit (hCGββ) both have autocrine growth effects. Tumor Biol. 2004, 25, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Redman, C.W. Current topic: Pre-eclampsia and the placenta. Placenta 1991, 12, 301–308. [Google Scholar] [CrossRef]
- Barros, J.S.; Baptista, M.G.; Bairos, V.A. Human chorionic gonadotropin in human placentas from normal and preeclamptic pregnancies. Arch. Gynecol. Obstet. 2002, 266, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Beck, T.; Schweikhart, G.; Stolz, E. Immunohistochemical location of HPL, SP1 and β-HCG in normal placentas of varying gestational age. Arch. Gynecol. 1986, 239, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Kurman, R.J.; Main, C.S.; Chen, H.-C. Intermediate trophoblast: A distinctive form of trophoblast with specific morphological, biochemical and functional features. Placenta 1984, 5, 349–369. [Google Scholar] [CrossRef]
- Prater, M.R.; Laudermilch, C.L.; Liang, C.; Holladay, S.D. Placental oxidative stress alters expression of murine osteogenic genes and impairs fetal skeletal formation. Placenta 2008, 29, 802–808. [Google Scholar] [CrossRef] [PubMed]
- Myatt, L. Review: Reactive oxygen and nitrogen species and functional adaptation of the placenta. Placenta 2010, 31, S66–S69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, T.; Tao, H.; Goto, M.; Yamada, H.; Suzuki, M.; Wu, Y.; Xiao, N.; He, Q.; Guo, W.; Cai, Z.; et al. Lipid peroxidation-induced DNA adducts in human gastric mucosa. Carcinogenesis 2013, 34, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Finkel, T. Oxidant signals and oxidative stress. Curr. Opin. Cell Biol. 2003, 15, 247–254. [Google Scholar] [CrossRef]
- Lee, H.M.; Choi, K.C. Cigarette smoke extract and isoprene resulted in the induction of apoptosis and autophagy in human placenta choriocarcinoma JEG-3 cells. Environ. Toxicol. 2018, 33, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Sun, F.; Que, S.; Li, Y.; Yang, S.; Liu, G. Prepregnancy maternal diabetes combined with obesity impairs placental mitochondrial function involving Nrf2/ARE pathway and detrimentally alters metabolism of offspring. Obes. Res. Clin. Pract. 2018, 12, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, S.; Rodriguez, L.; Otero, P.; Panadero, M.I.; Garcia, A.; Barbas, C.; Roglans, N.; Ramos, S.; Goya, L.; Laguna, J.C.; et al. Fructose during pregnancy provokes fetal oxidative stress: The key role of the placental heme oxygenase-1. Mol. Nutr. Food Res. 2016, 60, 2700–2711. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Guo, T.; Li, G.; Sun, S.; He, S.; Cheng, B.; Shi, B.; Shan, A. Dietary resveratrol improves antioxidant status of sows and piglets and regulates antioxidant gene expression in placenta by Keap1-Nrf2 pathway and Sirt1. J. Anim. Sci. Biotechnol. 2018, 9, 34. [Google Scholar] [CrossRef] [PubMed]
- Gurusinghe, S.; Cox, A.G.; Rahman, R.; Chan, S.T.; Muljadi, R.; Singh, H.; Leaw, B.; Mockler, J.C.; Marshall, S.A.; Murthi, P.; et al. Resveratrol mitigates trophoblast and endothelial dysfunction partly via activation of nuclear factor erythroid 2-related factor-2. Placenta 2017, 60, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Hobson, S.R.; Gurusinghe, S.; Lim, R.; Alers, N.O.; Miller, S.L.; Kingdom, J.C.; Wallace, E.M. Melatonin improves endothelial function in vitro and prolongs pregnancy in women with early-onset preeclampsia. J. Pineal Res. 2018, e12508. [Google Scholar] [CrossRef] [PubMed]
- Chiapella, G.; Flores-Martin, J.; Ridano, M.E.; Reyna, L.; de Potas, G.M.; Panzetta-Dutari, G.M.; Genti-Raimondi, S. The organophosphate chlorpyrifos disturbs redox balance and triggers antioxidant defense mechanisms in JEG-3 cells. Placenta 2013, 34, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Zhang, C.; Xin, Y.; Huang, Z.; Tan, Y.; Huang, Y.; Wang, Y.; Feng, W.; Li, X.; Li, W.; et al. Protective effect of FGF21 on type 1 diabetes-induced testicular apoptotic cell death probably via both mitochondrial- and endoplasmic reticulum stress-dependent pathways in the mouse model. Toxicol. Lett. 2013, 219, 65–76. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | N | Resorbed Embryos (%) | Dead Embryos (%) | Live Embryos (%) |
|---|---|---|---|---|
| E12.5 d | ||||
| Control | 6 | 2.0 (1/51) | 0 (0/51) | 98.0 (50/51) |
| DON-L | 6 | 17.6 ** (9/51) | 2.0 (1/51) | 80.4 (41/51) |
| DON-H | 6 | 28.0 ** (14/50) | 6.0 (3/50) | 66.0 (33/50) |
| E18.5 d | ||||
| Control | 6 | 2.1 (1/48) | 0 (0/48) | 97.9 (47/48) |
| DON-L | 6 | 10.2 (5/49) | 0 (0/49) | 89.8 (44/49) |
| DON-H | 6 | 18.8 ** (9/48) | 2.1 (1/48) | 79.1 (38/48) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, M.; Wei, Z.-Y.; Xu, Z.-H.; Pan, J.-Q.; Chen, J.-H. Oxidative Damage and Nrf2 Translocation Induced by Toxicities of Deoxynivalenol on the Placental and Embryo on Gestation Day 12.5 d and 18.5 d. Toxins 2018, 10, 370. https://doi.org/10.3390/toxins10090370
Yu M, Wei Z-Y, Xu Z-H, Pan J-Q, Chen J-H. Oxidative Damage and Nrf2 Translocation Induced by Toxicities of Deoxynivalenol on the Placental and Embryo on Gestation Day 12.5 d and 18.5 d. Toxins. 2018; 10(9):370. https://doi.org/10.3390/toxins10090370
Chicago/Turabian StyleYu, Miao, Zhi-Yuan Wei, Zhou-Heng Xu, Jia-Qi Pan, and Jian-Huan Chen. 2018. "Oxidative Damage and Nrf2 Translocation Induced by Toxicities of Deoxynivalenol on the Placental and Embryo on Gestation Day 12.5 d and 18.5 d" Toxins 10, no. 9: 370. https://doi.org/10.3390/toxins10090370
APA StyleYu, M., Wei, Z.-Y., Xu, Z.-H., Pan, J.-Q., & Chen, J.-H. (2018). Oxidative Damage and Nrf2 Translocation Induced by Toxicities of Deoxynivalenol on the Placental and Embryo on Gestation Day 12.5 d and 18.5 d. Toxins, 10(9), 370. https://doi.org/10.3390/toxins10090370