A Long-Term Time Series of Dinophysis acuminata Blooms and Associated Shellfish Toxin Contamination in Port Underwood, Marlborough Sounds, New Zealand

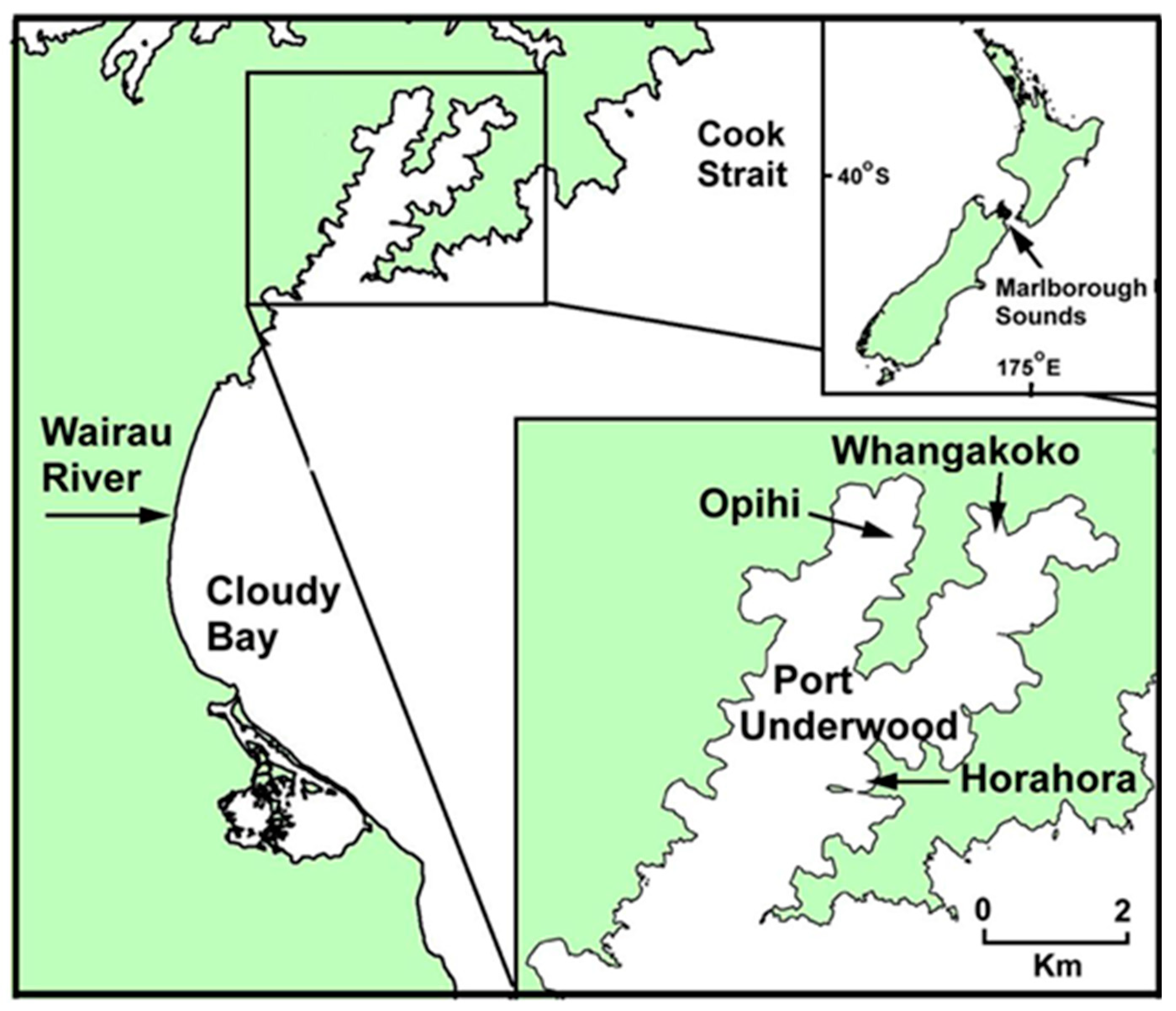{kind=link}
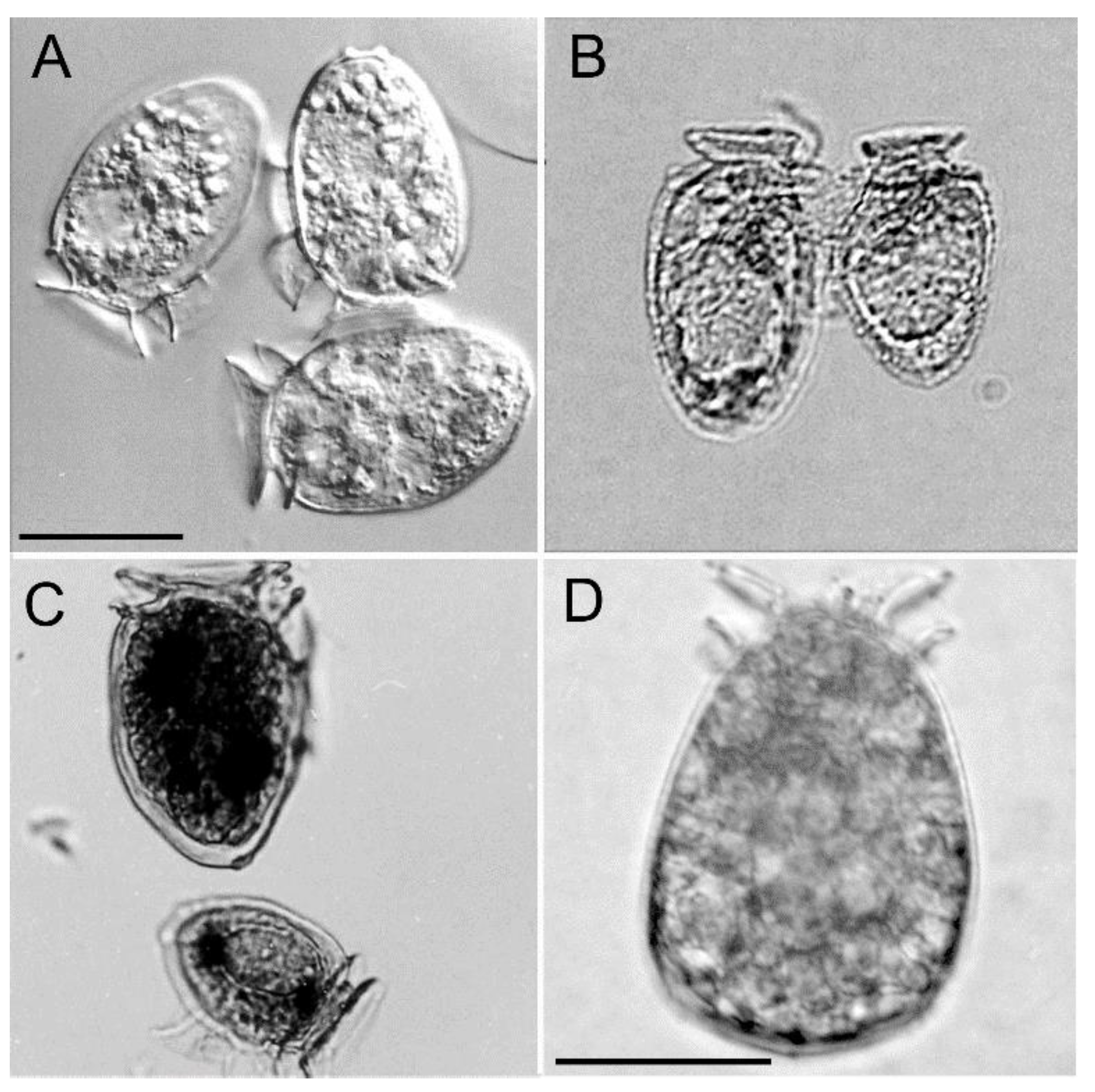{kind=link}
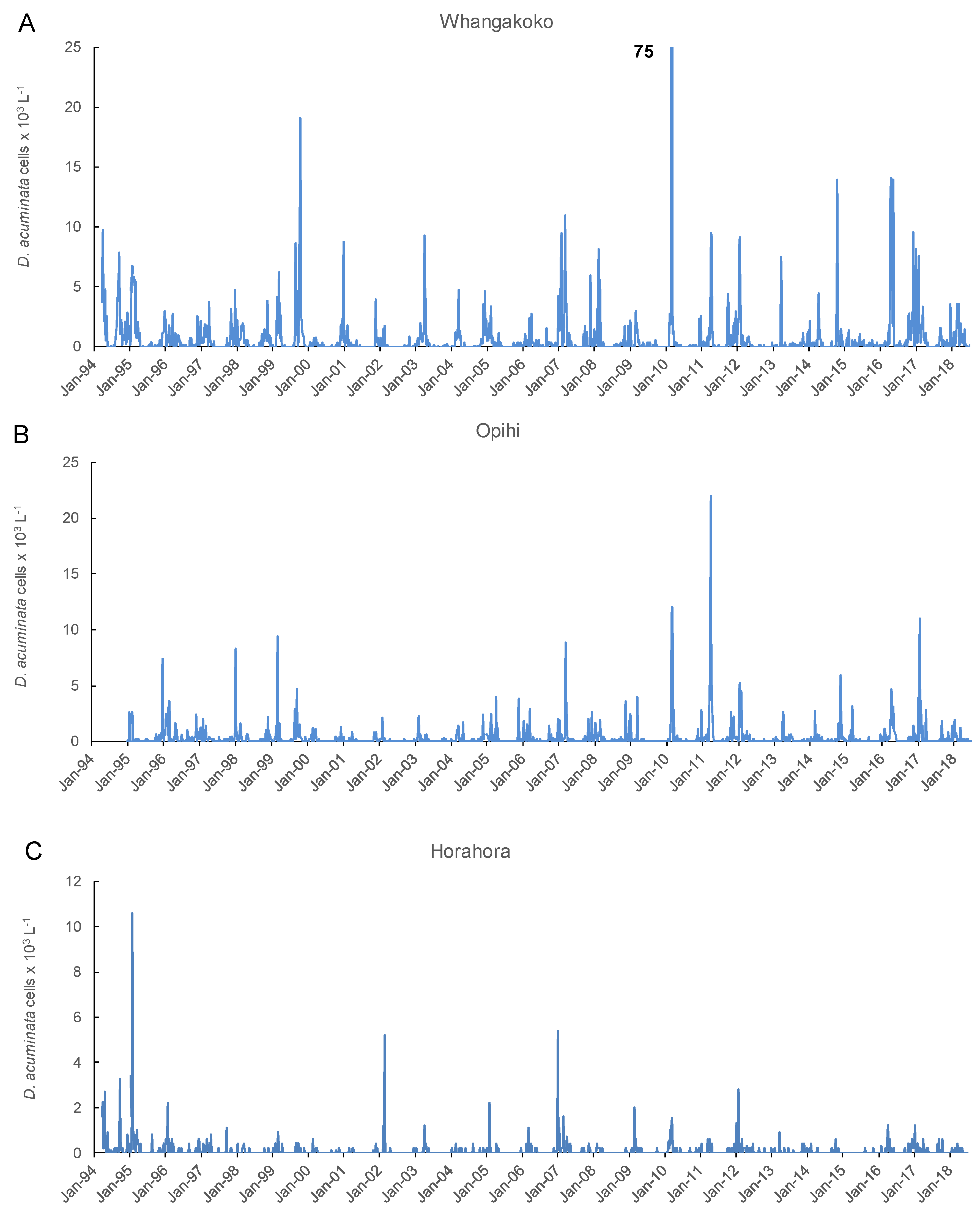{kind=link}
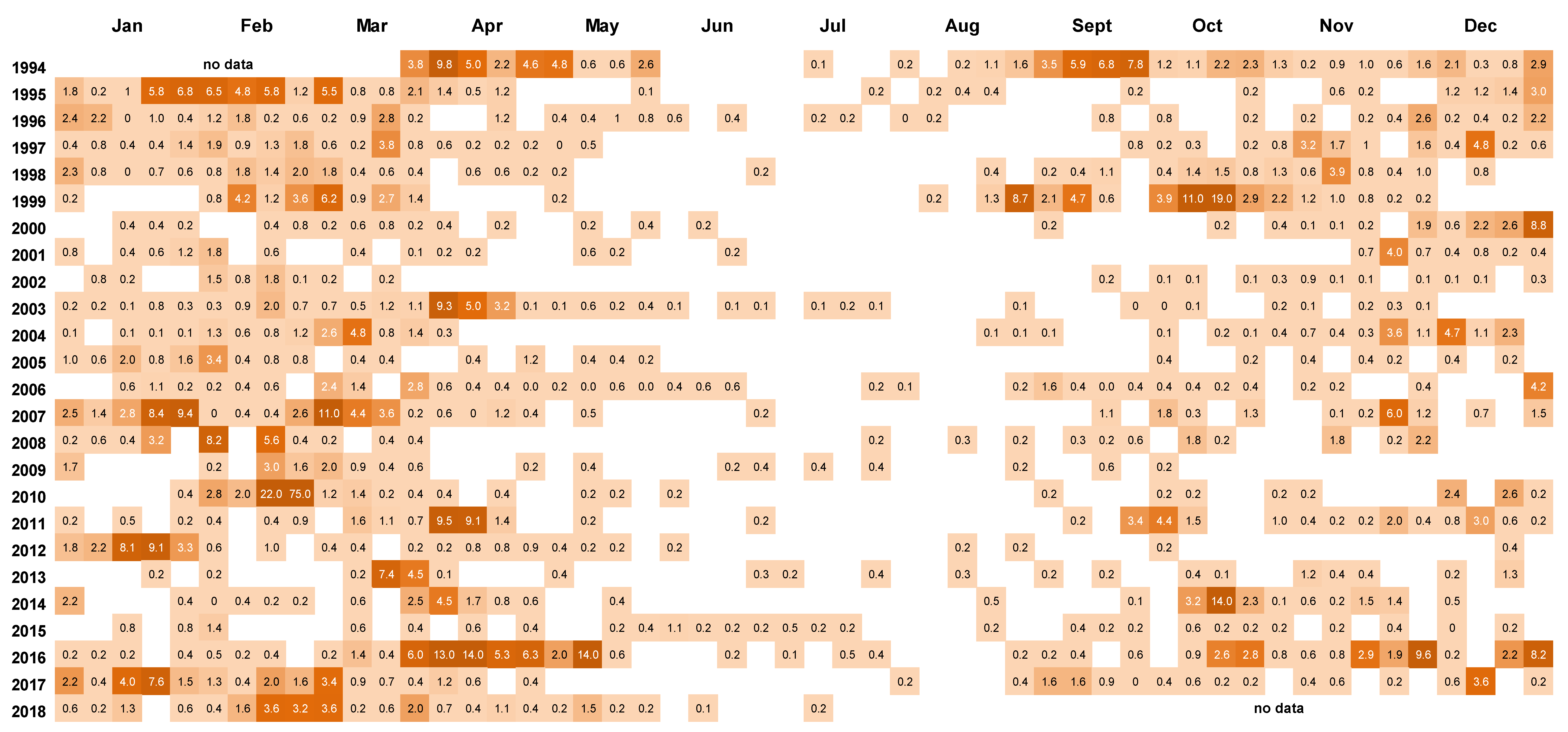{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Dinophysis acuminata Morphology
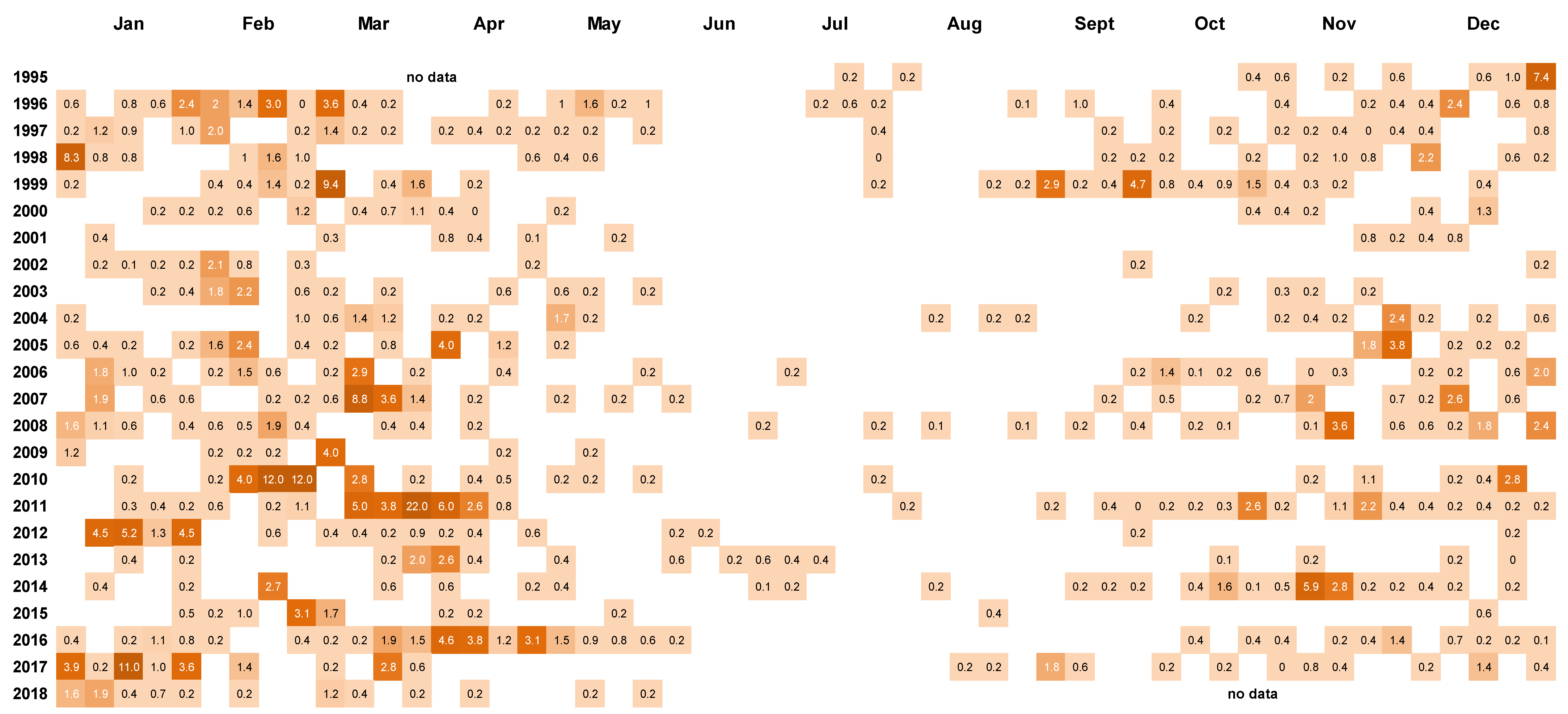
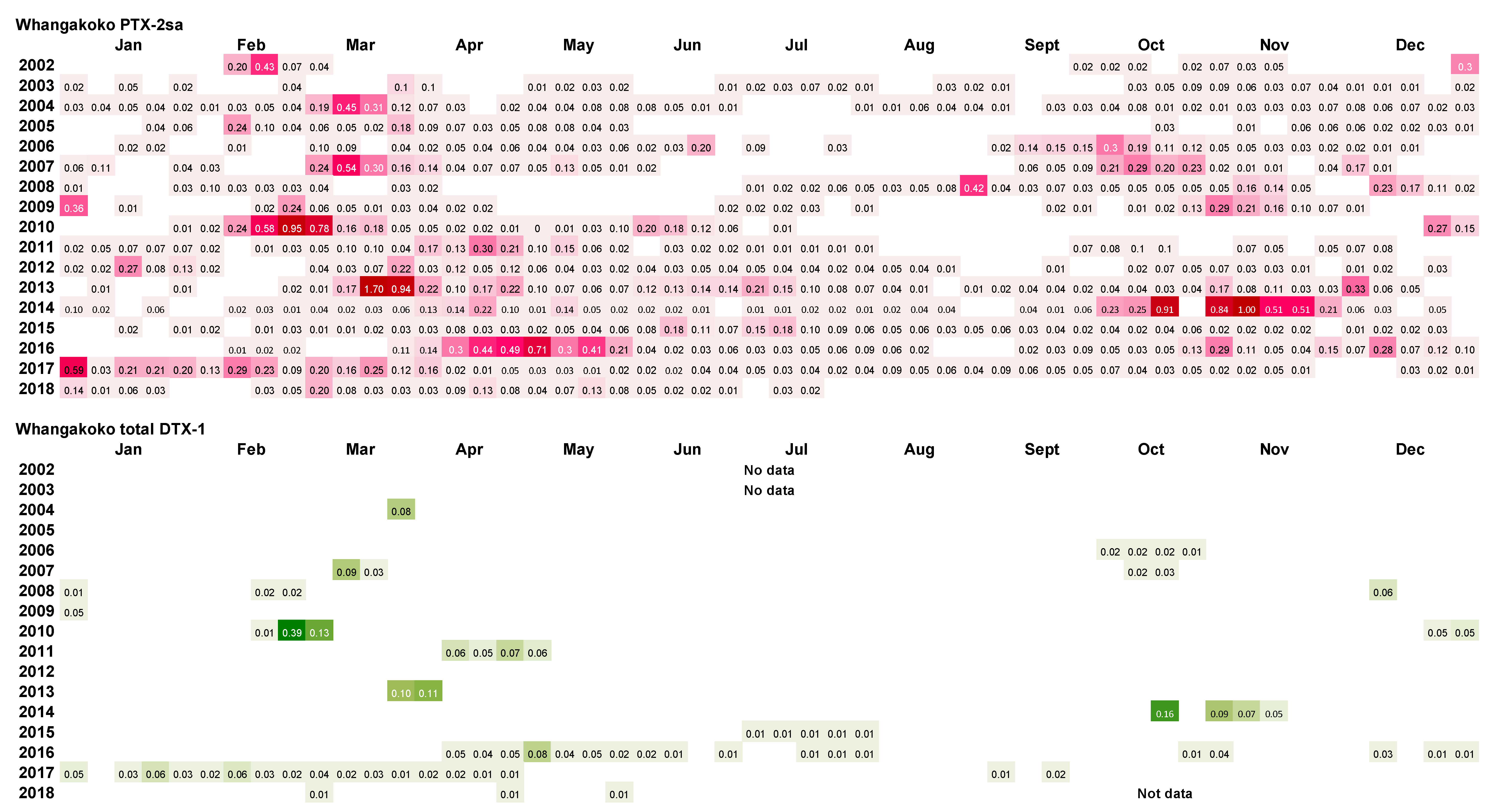
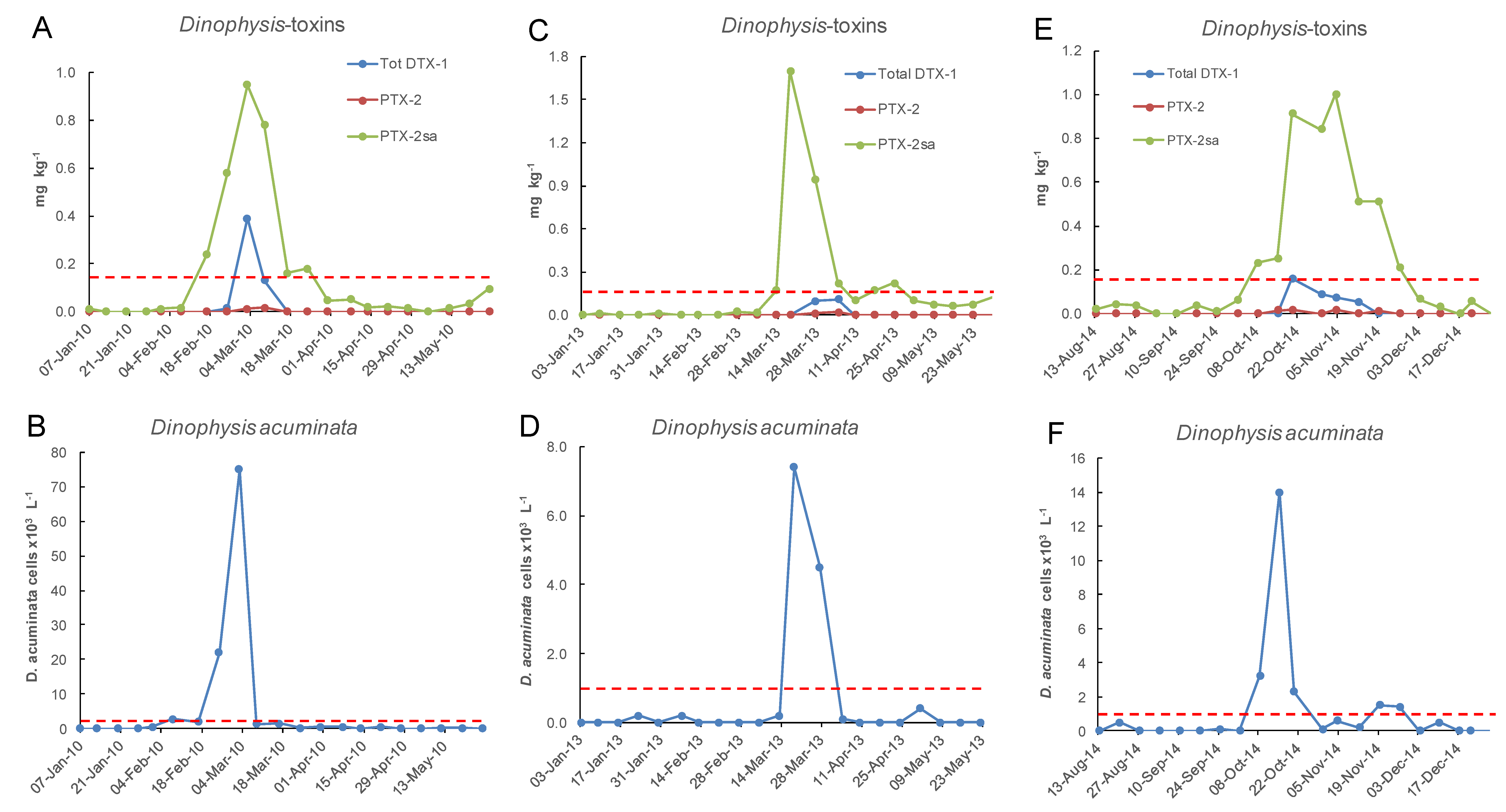
2.2. Frequency of Dinophysis acuminata Blooms
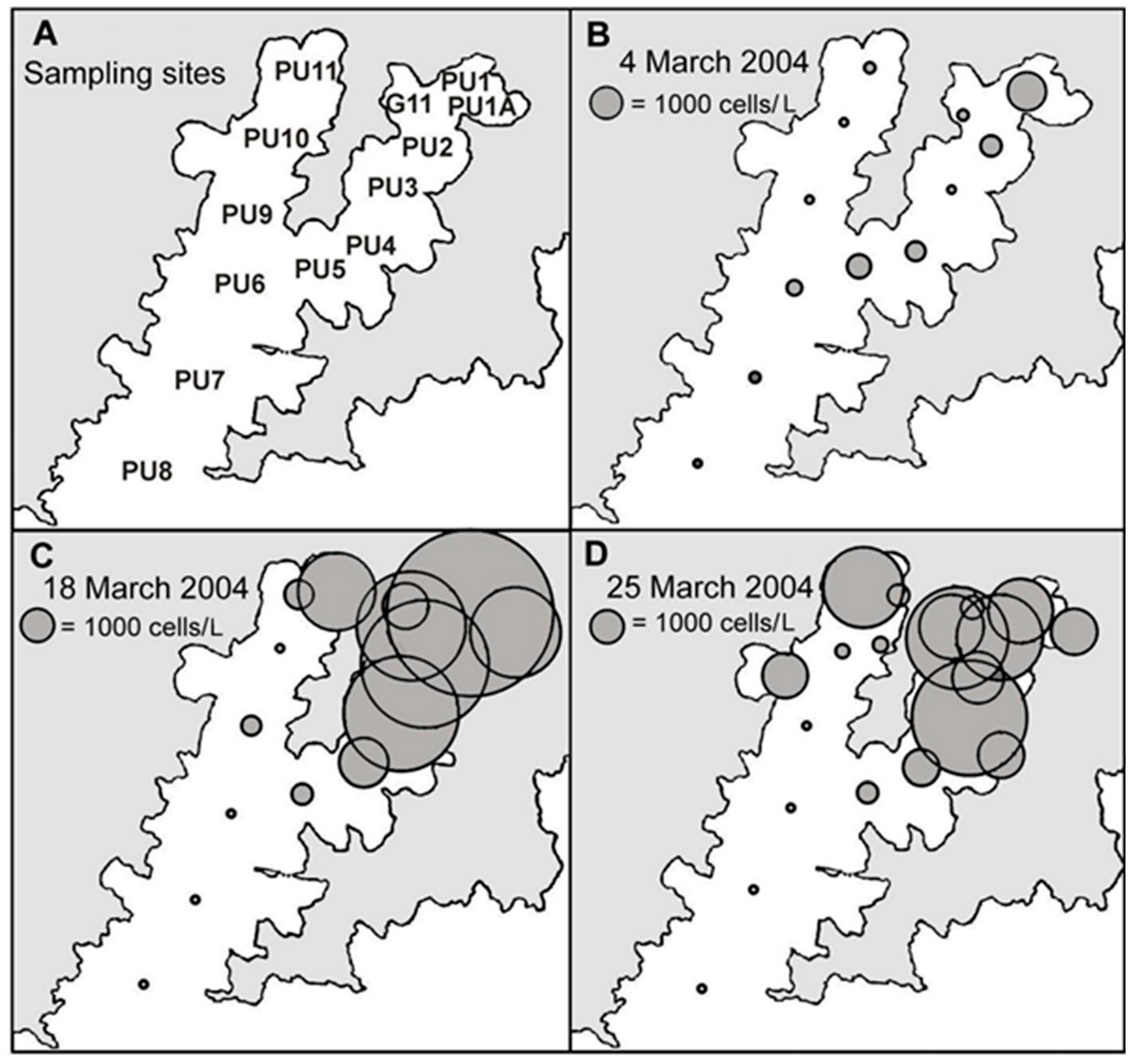
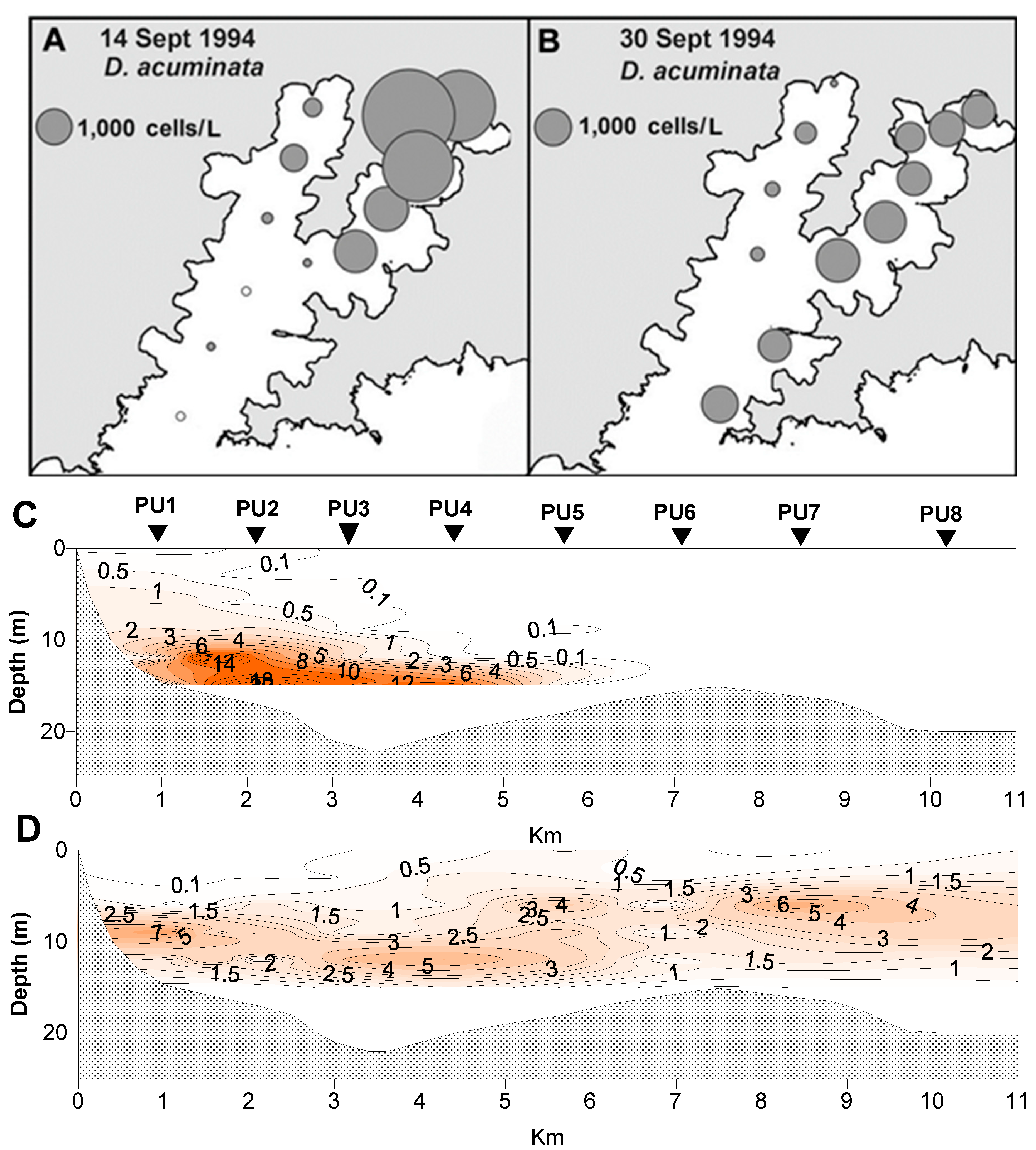
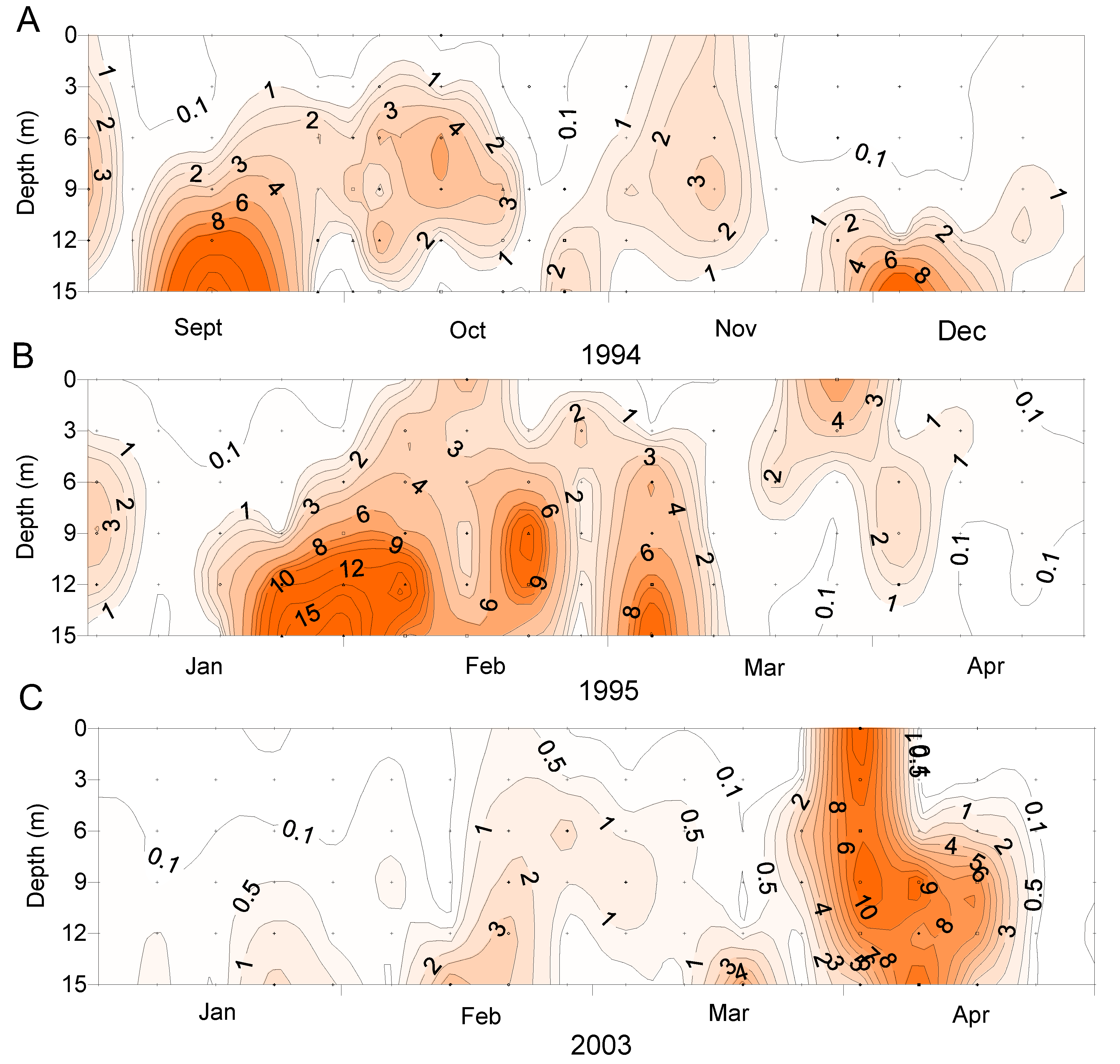
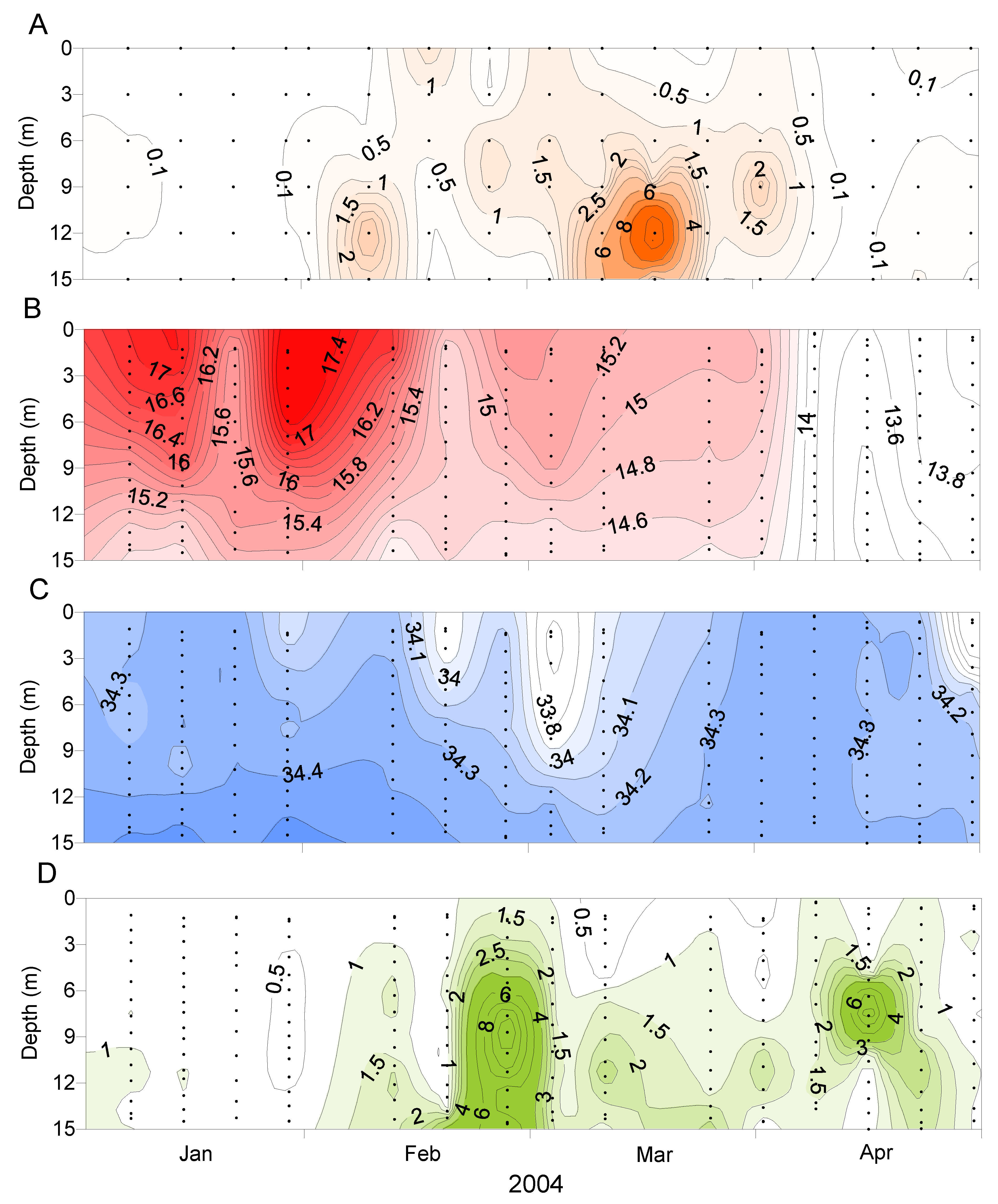
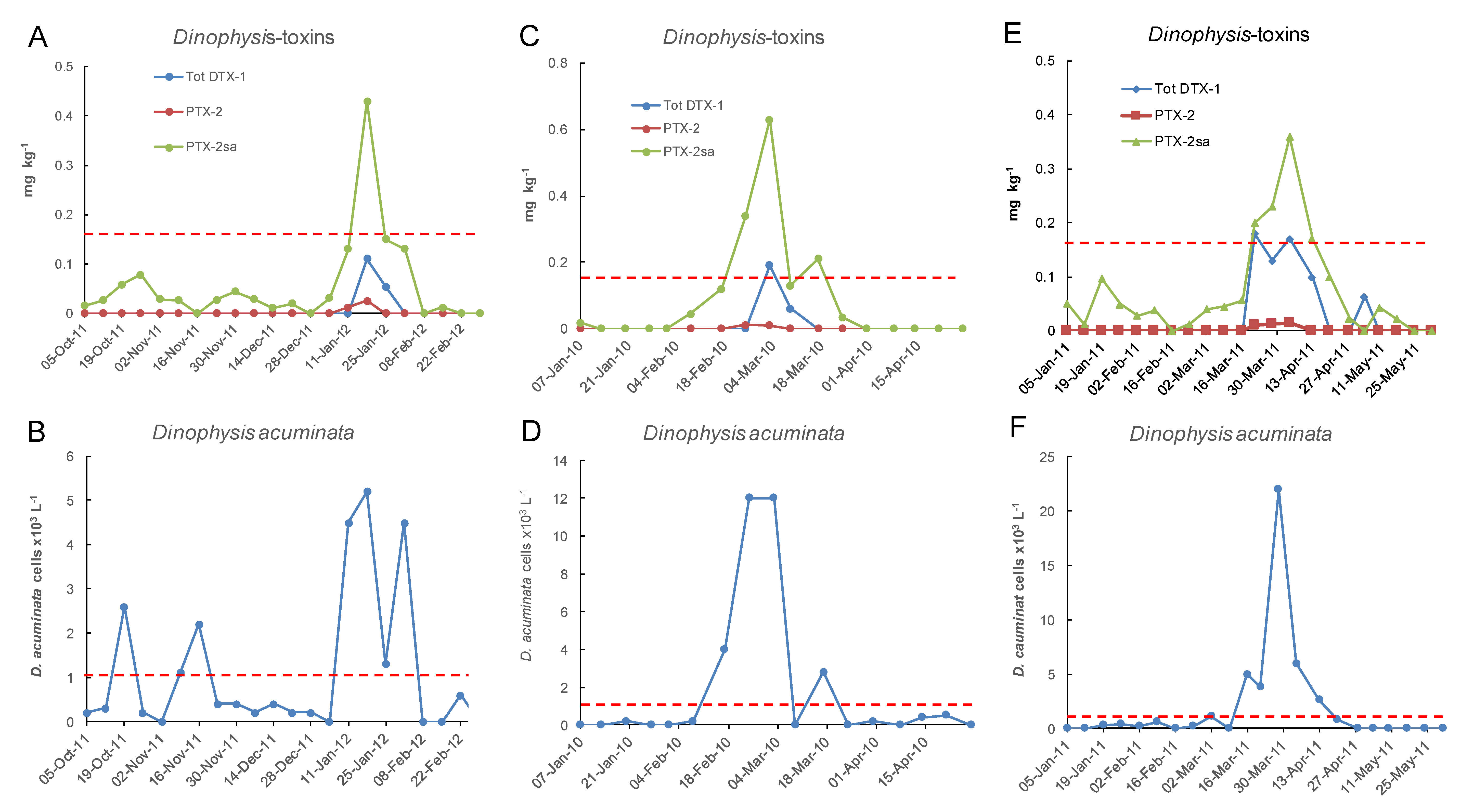
2.3. Spatial and Temporal Distribution of Blooms
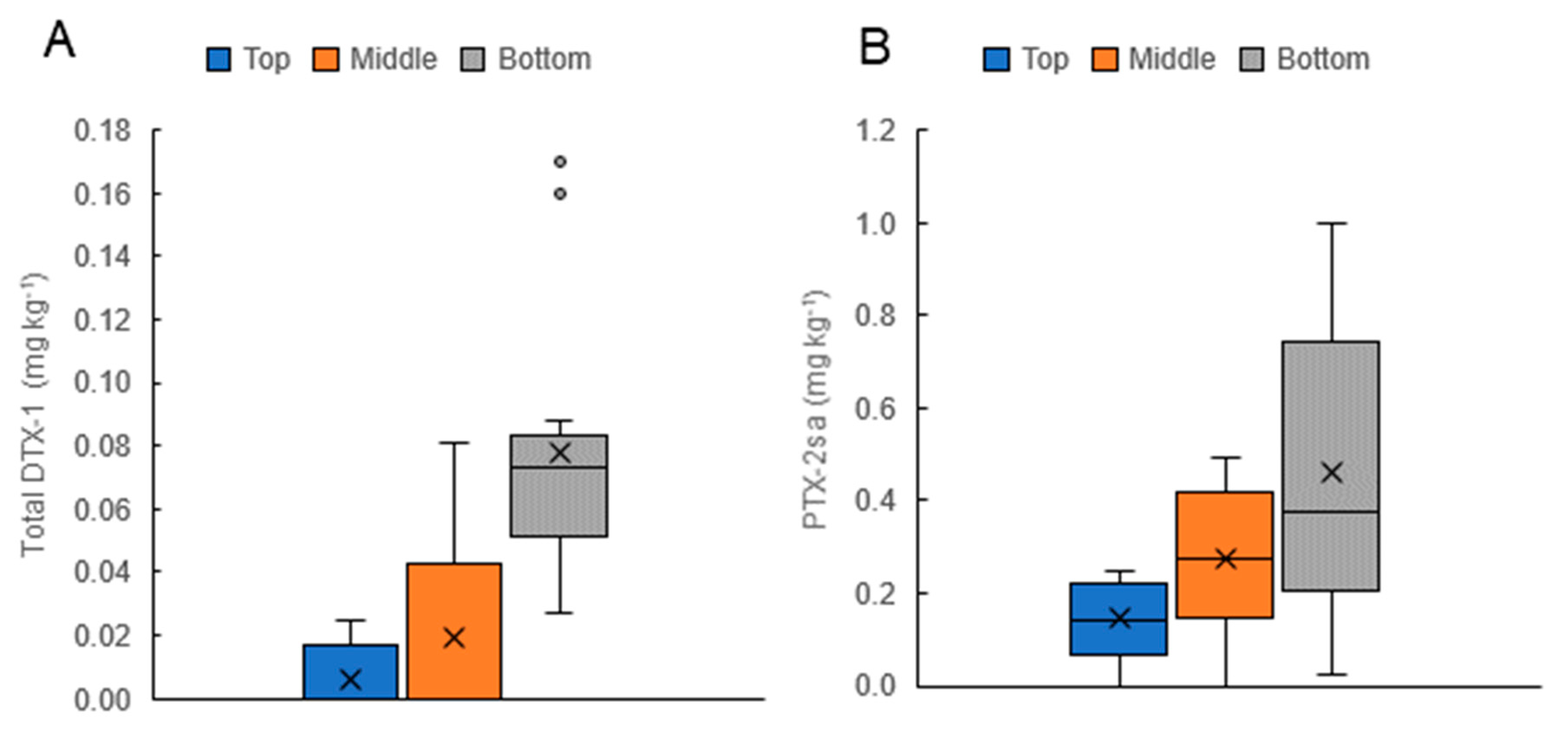
2.4. Toxins Originating from D. acuminata in Cultivated Mussels (Perna canaliculus)
3. Conclusions
4. Materials and Methods
Funding
Acknowledgments
Conflicts of Interest
References
- Yasumoto, T.; Oshima, Y.; Yamaguchi, M. Occurrence of a new type of shellfish poisoning in the Tohoku district. Bull. Jpn. Soc. Sci. Fish. 1978, 44, 1249–1255. [Google Scholar] [CrossRef]
- Yasumoto, T.; Oshima, Y.; Sugawara, W.; Fukuyo, Y.; Oguri, H.; Igarashi, T.; Fujita, N. Identification of Dinophysis fortii as the causative organism of Diarrhetic Shellfish Poisoning. Bull. Jpn. Soc. Sci. Fish. 1980, 46, 1405–1411. [Google Scholar] [CrossRef]
- European Parliament Council. Regulation (EC) 853 of April laying down specific hygiene rules for food of animal origin. Off. J. Eur. Union 2004, 226, 22–80. [Google Scholar]
- MacKenzie, L.; Beuzenburg, V.; McNabb, P.; Holland, P.; Suzuki, T.; Neil, T.; Selwood, A. Pectenotoxin and okadaic acid based toxin profiles in Dinophysis acuta and D. acuminata from New Zealand. Harmful Algae 2004, 4, 75–85. [Google Scholar] [CrossRef]
- Tong, M.; Smith, J.L.; Richlen, M.; Steidinger, K.A.; Kulis, D.M.; Fux, E.; Anderson, D.M. Characterization and comparison of toxin producing isolates s of Diniophysis acuminata from New England and Canada. J. Phycol. 2015, 51, 66–81. [Google Scholar] [CrossRef] [PubMed]
- Draisci, R.; Lucentini, L.; Mascioni, A. Pectenotoxins and Yessotoxin: Chemistry, Toxicology, Pharmacology and Analysis. In Seafood and Freshwater Toxins: Pharmacology Physiology, and Detection; Botana, L.M., Ed.; Marcel Dekker Inc.: New York, NY, USA; Basel, Switzerland, 2000; pp. 289–324. [Google Scholar]
- Munday, R. Toxicology of the pectenotoxins. In Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection, 2nd ed.; Botana, L.M., Ed.; CRC Press: Boca Raton, FL, USA; London, UK; New York, NY, USA, 2008; pp. 371–380. [Google Scholar]
- Daiguji, M.; Satake, M.; James, K.J.; Bishop, A.; Mackenzie, L.; Naoki, H.; Yasumoto, T. Structures of new Pectenotoxin analogs, Pectenotoxin-2 Seco acid and 7-epi-Pectenotoxin-2 seco acid, isolated from a dinoflagellate and Greenshell mussels. Chem. Lett. 1998, 7, 653–654. [Google Scholar] [CrossRef]
- Suzuki, T.; Mackenzie, L.; Stirling, D.; Adamson, J. Pectenotoxin-2 Seco Acid: A toxin converted from Pectenotoxin-2 by the New Zealand Greenshell mussel Perna canaliculus. Toxicon 2001, 39, 507–514. [Google Scholar] [CrossRef]
- MacKenzie, L.A.; Selwood, A.I.; Marshall, C. Isolation and characterization of an enzyme from the GreenshellTM mussel Perna canaliculus that hydrolyses pectenotoxins and esters of okadaic acid. Toxicon 2012, 60, 406–419. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, L.; Holland, P.; McNabb, P.; Beuzenberg, V.; Selwood, A.; Suzuki, T. Complex toxin profiles in phytoplankton and Greenshell mussels (Perna canaliculus) revealed by LC-MS/MS analysis. Toxicon 2002, 40, 1321–1330. [Google Scholar] [CrossRef]
- Mackenzie, L. An Evaluation of the Risk to Consumers of Pectenotoxn-2 Seco Acid (PTX2-SA) Contamination of GreenshellTM Mussels; A Report for Marlborough Sounds Shellfish Quality Programme; Cawthron Report No. 750; Cawthron: Nelson, New Zealand, 2002; 50p. [Google Scholar]
- MacKenzie, A.L. Does Dinophysis (Dinophyceae) have a sexual life cycle? J. Phycol. 1992, 28, 399–406. [Google Scholar] [CrossRef]
- Reguera, B.; González-Gill, S. Small cell and intermediate cell formation in species of Dinophysis (Dinophyceae, Dinophysiales). J. Phycol. 2001, 37, 318–333. [Google Scholar] [CrossRef]
- Koike, K.; Nishiyama, A.; Saitoh, K.; Imai, K.; Koike, K.; Kobiyama, A.; Ogata, T. Mechanism of gamete fusion in Dinophysis fortii (Dinophyceae, Dinophyta): Light microscopic and ultrastructural observations. J. Phycol. 2006, 42, 1247–1256. [Google Scholar] [CrossRef]
- Park, M.G.; Kim, S.; Kim, H.S.; Myung, G.; Kang, Y.G.; Yih, W. First successful culture of the marine dinoflagellate Dinophysis acuminata. Aquat. Microb. Ecol. 2006, 45, 101–106. [Google Scholar] [CrossRef]
- Reguera, B.; Velo-Suárez, L.; Raine, R.; Park, M.G. Harmful Dinophysis species: A review. Harmful Algae 2012, 14, 87–106. [Google Scholar] [CrossRef]
- Velo-Suárez, L.; González-Gil, S.; Pazos, Y.; Reguera, B. The growth season of Dinophysis acuminata in an upwelling system embayment: A conceptual model based on in situ measurements. Deep-Sea Res. 2014, 101, 141–151. [Google Scholar] [CrossRef]
- Maestrini, S.Y. Bloom dynamics and ecophysiology of Dinophysis spp. In Physiological Ecology of Harmful Algal Blooms. NATO AI Series G. Ecological Science; Anderson, D.M., Cembella, A.D., Hallegraeff, G.M., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1998; pp. 243–266. [Google Scholar]
- Raine, R.; McDermott, G.; Silke, J.; Lyons, K.; Cusack, C. A simple short-range model for the prediction of harmful algal events in the bays of Southwest Ireland. J. Mar. Syst. 2010, 83, 150–157. [Google Scholar] [CrossRef]
- McNabb, P.; Holland, P. Using LCMS to manage shellfish harvesting and protect public health. In Molluscan Shellfish Safety; Villalba, A., Reguera, B., Romalde, J., Beiras, R., Eds.; Xunta de Glaicia & IOC of UNESCO: Santiago de Compostela, Spain, 2003; pp. 179–186. [Google Scholar]
- McNabb, P.; Selwood, A.I.; Holland, P.T. Multi-residue method for determination of algal toxins in shellfish: Single laboratory validation and inter-laboratory study. J. AOAC Int. 2005, 88, 761–772. [Google Scholar] [PubMed]
- Torgersen, T.; Sanvik, M.; Lundve, B.; Lindegart, S. Profiles and levels of fatty acid esters of okadaic acid group toxins and pectenotoxins during toxin depuration. Part II: Blue mussels (Mytilus edulis) and flat oyster (Ostrea edulis). Toxicon 2008, 52, 418–427. [Google Scholar] [CrossRef]
- Blanco, J.; Alvarez, G.; Rengel, J.; Diaz, R.; Marinao, C.; Martin, H.; Uribe, E. Accumulation and biotransformation of Dinophysis toxins by the surf clam Mesodesma donacium. Toxins 2018, 10, 314. [Google Scholar] [CrossRef] [PubMed]
- Abal, P.; Louzao, M.C.; Suzuki, T.; Watanabe, R.; Vilarino, N.; Carrera, C.; Botana, A.M.; Vieytes, M.R.; Botana, L.M. Toxic action re-evaluation of Okadaic acid, Dinophysistoxin-1 and Dinophysistoxin-2: Toxicity equivalency factors based on the oral toxicity study. Cell. Physiol. Biochem. 2018, 49, 743–757. [Google Scholar] [CrossRef]
- Kamiyama, T.; Nagai, S.; Miyamura, K. Effect of temperature on production of okadaic acid, dinophysistoxin-1 and pectenotoxin-2 by Dinophysis acuminata in culture experiments. Aquat. Microb. Ecol. 2010, 60, 193–202. [Google Scholar] [CrossRef]
- Pizarro, G.; Paz, B.; González-Gil, S.; Franco, J.M.; Reguera, B. Seasonal variability of lipophilic toxins during a Dinophysis acuta bloom in Western Iberia: Differences between picked cells and plankton concentrates. Harmful Algae 2009, 8, 926–937. [Google Scholar] [CrossRef]
- Hannah, D.J.; Till, D.G.; Deverall, T.; Jones, P.D.; Fry, J.M. Extraction of lipid-soluble marine biotoxins. J. AOAC Int. 1995, 78, 480–483. [Google Scholar]
- Mountfort, D.O.; Suzuki, T.; Truman, P. Protein phosphatase inhibition assay adapted for determination of total DSP in contaminated mussels. Toxicon 2001, 39, 383–390. [Google Scholar] [CrossRef]














© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mackenzie, L.A. A Long-Term Time Series of Dinophysis acuminata Blooms and Associated Shellfish Toxin Contamination in Port Underwood, Marlborough Sounds, New Zealand. Toxins 2019, 11, 74. https://doi.org/10.3390/toxins11020074
Mackenzie LA. A Long-Term Time Series of Dinophysis acuminata Blooms and Associated Shellfish Toxin Contamination in Port Underwood, Marlborough Sounds, New Zealand. Toxins. 2019; 11(2):74. https://doi.org/10.3390/toxins11020074
Chicago/Turabian StyleMackenzie, Lincoln A. 2019. "A Long-Term Time Series of Dinophysis acuminata Blooms and Associated Shellfish Toxin Contamination in Port Underwood, Marlborough Sounds, New Zealand" Toxins 11, no. 2: 74. https://doi.org/10.3390/toxins11020074
APA StyleMackenzie, L. A. (2019). A Long-Term Time Series of Dinophysis acuminata Blooms and Associated Shellfish Toxin Contamination in Port Underwood, Marlborough Sounds, New Zealand. Toxins, 11(2), 74. https://doi.org/10.3390/toxins11020074