Contamination of Wheat Cultivated in Various Regions of Poland during 2017 and 2018 Agricultural Seasons with Selected Trichothecenes and Their Modified Forms

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
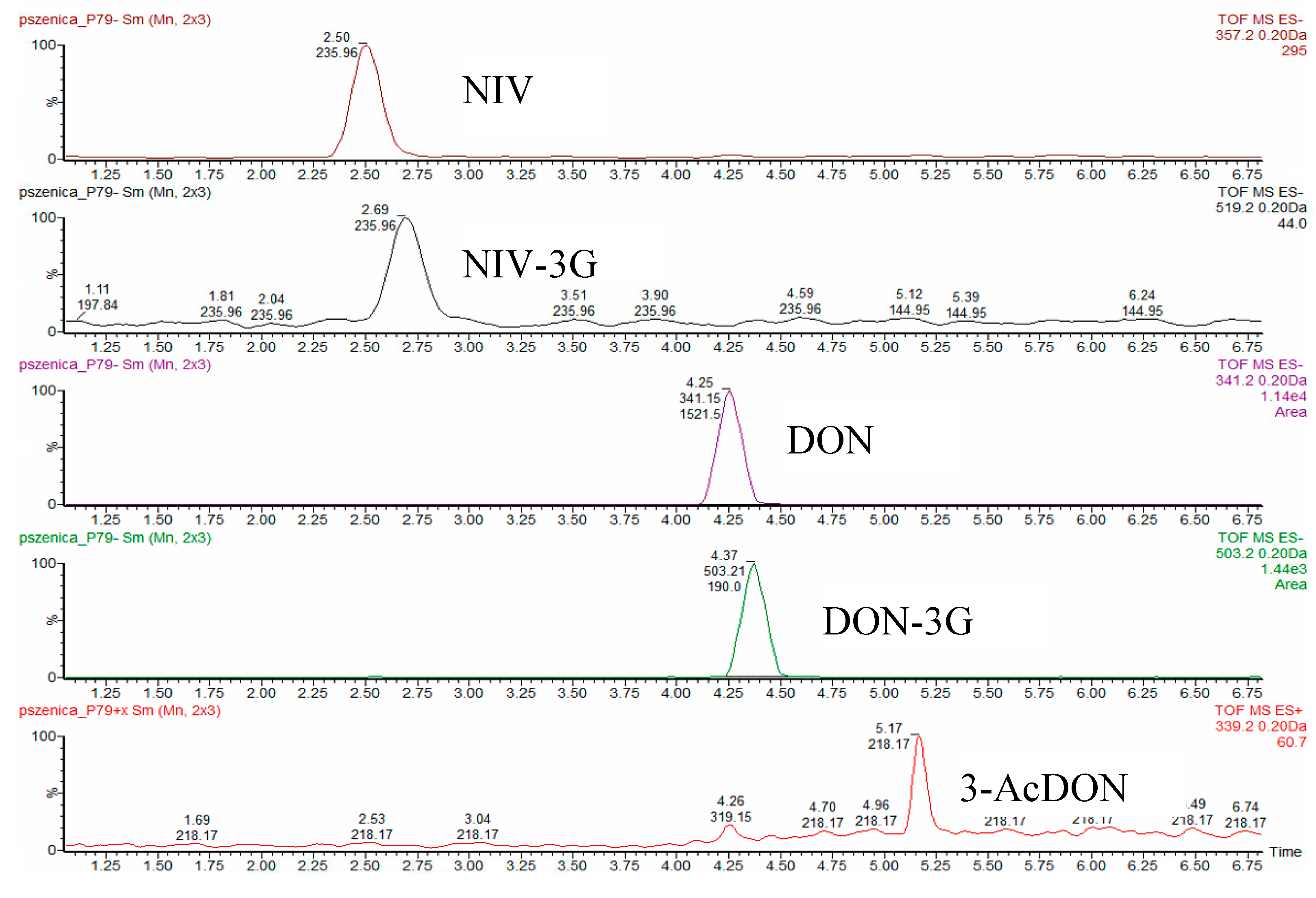
2.1. Method Validation
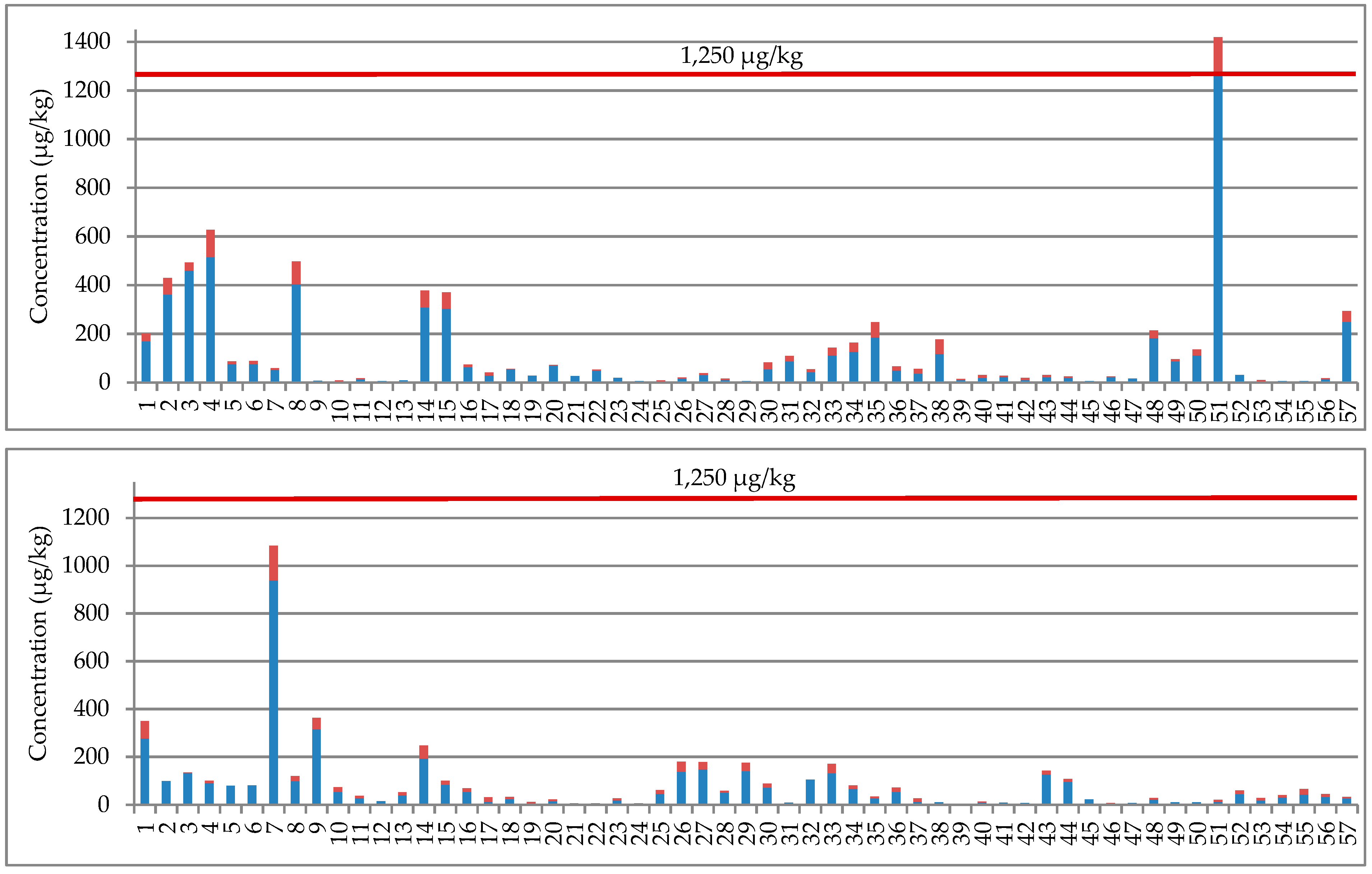
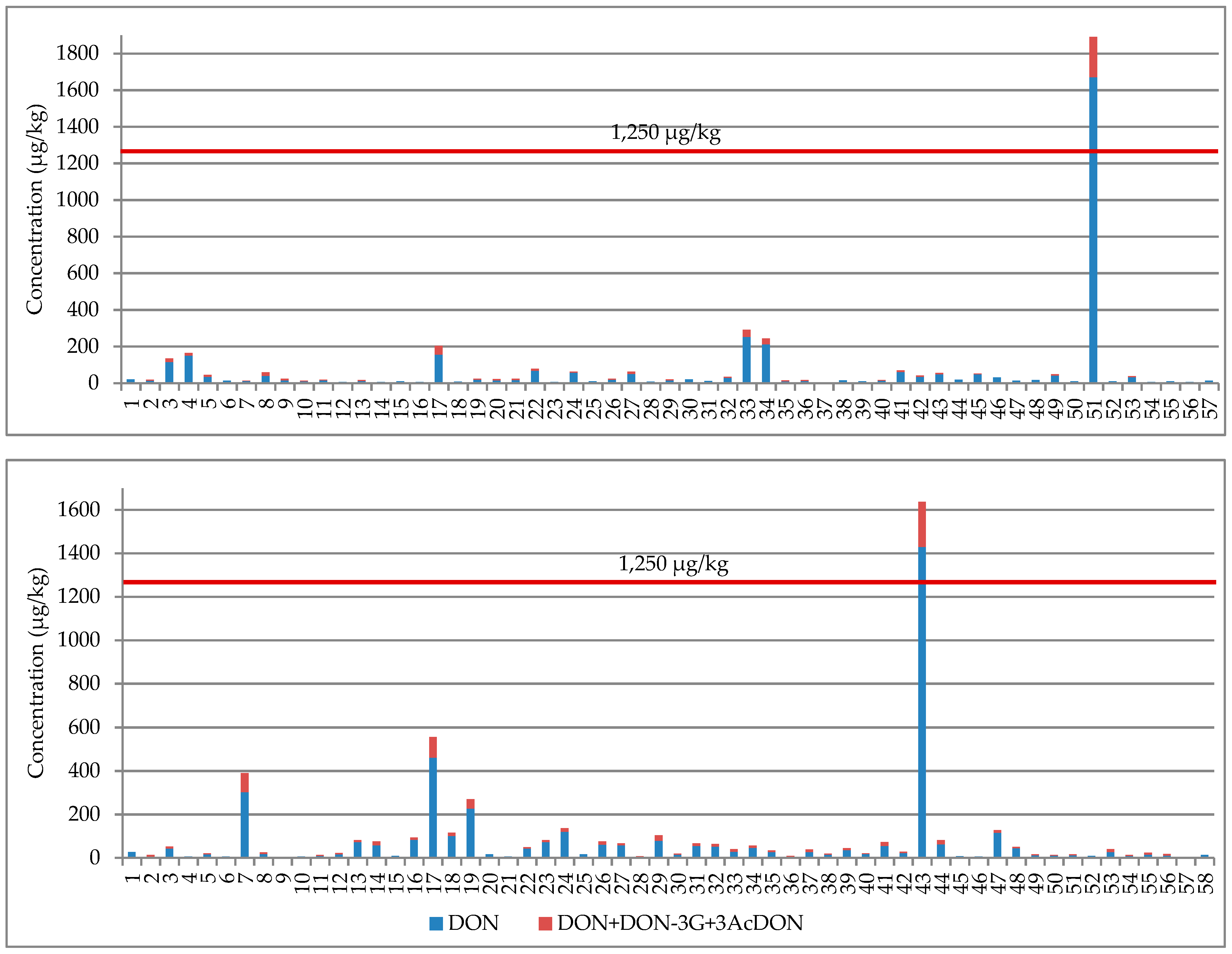
2.2. NIV, DON, Their Glucosides, and 3-AcDON in Wheat Grain
3. Conclusions
4. Materials and Methods
4.1. Reagents and Standards
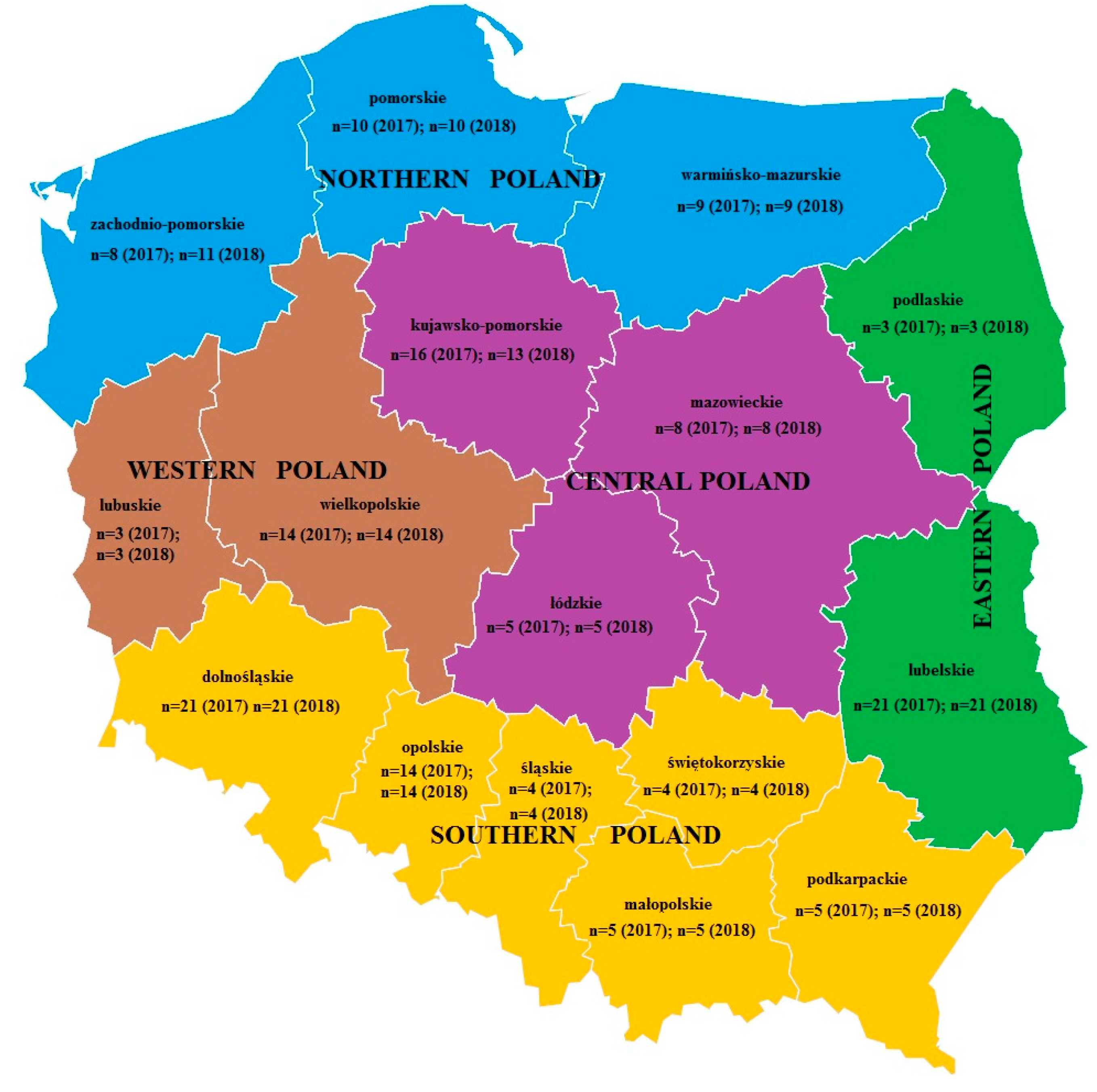
4.2. Research Material
4.3. LC-MS
4.4. Sample Preparation
4.5. Statistics
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dweba, C.; Figlan, S.; Shimelis, H.; Motaung, T.E.; Sydeham, S.; Mwadzingeni, L.; Tsilo, T.J. Fusarium head blight of wheat: Pathogenesis and control strategies. Crop Prot. 2017, 91, 114–122. [Google Scholar] [CrossRef]
- Dahl, B.; Wilson, W.W. Risk premiums due to Fusarium Head Blight (FHB) in wheat and barley. Agric. Syst. 2018, 162, 145–153. [Google Scholar] [CrossRef]
- Bruns, H.A. Controlling aflatoxin and fumonisin in maize by crop management. Toxin Rev. 2003, 22, 153–173. [Google Scholar] [CrossRef]
- Brakhage, A.A. Regulation of fungal secondary metabolism. Nat. Rev. Microbiol. 2013, 11, 21–32. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From simple to complex mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez-Vea, M.; Lizarraga, E.; González-Peñas, E.; López de Cerain, A. Co-occurrence of type-A and type-B trichothecenes in barley from a northern region of Spain. Food Control 2012, 25, 81–88. [Google Scholar] [CrossRef]
- Bryła, M.; Jędrzejczak, R.; Szymczyk, K.; Roszko, M.; Obiedziński, M.W. An LC-IT-MS/MS-based method to determine trichothecenes in grain products. Food Anal. Met. 2014, 7, 1056–1065. [Google Scholar] [CrossRef]
- Chelkowski, J.; Wisniewska, H.; Adamski, T.; Golinski, P.; Kaczmarek, Z.; Kostecki, M.; Perkowski, J.; Surma, M. Effects of Fusarium culmorum head blight on mycotoxin accumulation and yield traits in barley doubled haploids. J. Phytopathol. 2000, 148, 541–545. [Google Scholar] [CrossRef]
- Schaafsma, A.W.; Tamburic-Ilinic, L.; Miller, J.D.; Hooker, D.C. Agronomic considerations for reducing deoxynivalenol in wheat grain. Can. J. Pathol. 2001, 23, 279–285. [Google Scholar] [CrossRef]
- Doohan, F.M.; Brennan, J.; Cooke, B.M. Influence of climatic factors on Fusarium species pathogenic to cereals. Eur. J. Plant Pathol. 2003, 109, 755–768. [Google Scholar] [CrossRef]
- Bryła, M.; Waśkiewicz, A.; Podolska, G.; Szymczyk, K.; Jędrzejczak, R.; Damaziak, K.; Sułek, A. Occurrence of 26 mycotoxins in the grain of cereals cultivated in Poland. Toxins 2016, 8, 160. [Google Scholar] [CrossRef] [PubMed]
- Sutton, J.C. Epidemiology of wheat head blight and maize ear rot caused by Fusarium graminearum. Can. J. Plant Pathol. 1982, 4, 195–209. [Google Scholar] [CrossRef]
- Blaney, B.J.; Dodman, R.L. Production of zearalenone, deoxynivalenol, nivalenol, and acetylated derivated by Australian isolates of Fusarium graminearum and F. pseudograminearum in relation to source and culturing conditions. Aust. J. Agric. Res. 2002, 53, 1317–1326. [Google Scholar] [CrossRef]
- Hooker, D.C.; Schaafsma, A.W.; Tamburic-Ilincic, L. Using weather variables pre- and post-heading to predict deoxynivalenol content in winter wheat. Plant Dis. 2002, 86, 611–619. [Google Scholar] [CrossRef]
- Cromey, M.G.; Shorter, S.C.; Lauren, D.R.; Sinclair, K.I. Cultivar and crop management influences on fusarium head blight and mycotoxins in spring wheat (Triticum aestivum) in New Zealand. N. Z. J. Crop Hortic. Sci. 2002, 30, 235–247. [Google Scholar] [CrossRef]
- Parry, D.W.; Jenkinson, P.; McLeod, L. Fusarium ear blight (scab) in small grain cereals—A review. Plant Pathol. 1995, 44, 207–238. [Google Scholar] [CrossRef]
- Scott, L.; Leath, S.; Hagler, W.; Murphy, J. Variation in Fusarium graminearum associated with wheat scab in north Carolina. In National Fusarium Head Blight Forum. Chapter 1—Epidemiology and Disease Management; Michigan State University (USA): East Lansing, MI, USA, 1998; p. 15. [Google Scholar]
- Eriksen, G.S.; Pettersson, H. Toxicological evaluation of trichothecenes in animal feed. Anim. Feed Sci. Technol. 2004, 114, 205–239. [Google Scholar] [CrossRef]
- Commission Regulation (EC) No 1881/2006 of 19 December 2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32006R1881 (accessed on 31 January 2019).
- Wu, W.; Flannery, B.M.; Sugita-Konishi, Y.; Watanabe, M.; Zhang, H.; Pestka, J.J. Comparison of murine anorectic responses to the 8-ketotrichothecenes 3-acetyl-deoxynivalenol, 15-acetyl-deoxynivalenol, fusarenon X and nivalenol. Food Chem. Toxicol. 2012, 50, 2056–2061. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer (IARC). Some Naturally Occurring Substances: Food Items and Constituents, Heterocyclic Aromatic Amines and Mycotoxins. Monograph on the Evaluation of Carcinogenic Risks to Humans; World Health Organization, International Agency for Research on Cancer: Lyon, France, 1993; pp. 397–444. [Google Scholar]
- Abbas, H.K.; Yoshizawa, T.; Shier, W.T. Cytotoxicity and phytotoxicity of trichothecene mycotoxins produced by Fusarium spp. Toxicon 2013, 74, 68–75. [Google Scholar] [CrossRef]
- Berthiller, F.; Krska, R.; Domig, K.J.; Kneifel, W.; Juge, N.; Schuhmacher, R.; Adam, G. Hydrolytic fate of deoxynivalenol-3-glucoside during digestion. Toxicol. Lett. 2011, 206, 264–267. [Google Scholar] [CrossRef]
- Dall’Erta, A.; Cirlini, M.; Dall’Asta, M.; Del Rio, D.; Galaverna, G.; Dall’Asta, C. Masked mycotoxins are efficiently hydrolyzed by human colonic microbiota releasing their aglycones. Chem. Res. Toxicol. 2013, 26, 305–312. [Google Scholar] [CrossRef]
- Gratz, S.W.; Duncan, G.; Richardson, A.J. The human fecal microbiota metabolizes deoxynivalenol and deoxynivalenol-3-glucoside and may be responsible for urinary deepoxy-deoxynivalenol. Appl. Environ. Microbiol. 2013, 79, 1821–1825. [Google Scholar] [CrossRef]
- De Angelis, E.; Monaci, L.; Visconti, A. Investigation on the stability of deoxynivalenol and DON-3 glucoside during gastro-duodenal in vitro digestion of a naturally contaminated bread model food. Food Control 2014, 43, 270–275. [Google Scholar] [CrossRef]
- Evaluations of the Joint FAO/WHO Expert Committee on Food Additives (JECFA). Available online: http://apps.who.int/food-additives-contaminants-jecfa-database/chemical.aspx?chemID=2947 (accessed on 31 January 2019).
- Jia, H.; Zhou, J.; Xue, S.; Li, G.; Yan, H.; Ran, C.; Zhang, Y.; Shi, J.; Jia, L.; Wang, X.; et al. A journey to understand wheat Fusarium head blight resistance in the Chinese wheat landrace Wangshuibai. Crop J. 2018, 6, 48–59. [Google Scholar] [CrossRef]
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium toxins in cereals: Occurrence, legislation, factors promoting the appearance and their management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef] [PubMed]
- Atanasova-Penichon, V.; Barreau, C.; Richard-Forget, F. Antioxidant Secondary Metabolites in Cereals: Potential Involvement in Resistance to Fusarium and Mycotoxin Accumulation. Front. Microbiol. 2016, 7, 566. [Google Scholar] [CrossRef] [PubMed]
- Mesterhazy, A. Role of deoxynivalenol in aggressiveness of Fusarium graminearum and F. culmorum and in resistance to Fusarium head blight. Eur. J. Plant Pathol. 2002, 108, 675–684. [Google Scholar] [CrossRef]
- Rychlik, M.; Humpf, H.; Marko, D.; Dänicke, S.; Mally, A.; Berthiller, F.; Klaffke, H.; Lorenz, N. Proposal of a comprehensive definition of modified and other forms of mycotoxins including “masked” mycotoxins. Mycotoxin Res. 2014, 30, 197–205. [Google Scholar] [CrossRef]
- Berthiller, F.; Crews, C.; Dall’Asta, C.; De Saeger, S.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef]
- Commission Regulation (EC) No 401/2006 of 23 February 2006 laying down the methods of sampling and analysis for the official control of the levels of mycotoxins in foodstuffs. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/HTML/?uri=CELEX:32006R0401&from=en (accessed on 31 January 2019).
- Yoshinari, T.; Sakuda, S.; Furihata, K.; Furusawa, H.; Ohnishi, T.; Sugita-Konishi, Y.; Ishizaki, N.; Terajima, J. Structural determination of a nivalenol glucoside and development of an analytical method for the simultaneous determination of nivalenol and deoxynivalenol, and their glucosides, in wheat. J. Agric. Food Chem. 2014, 62, 1174–1180. [Google Scholar] [CrossRef]
- Geng, Z.; Yang, D.; Zhou, M.; Zhang, P.; Wang, D.; Liu, F.; Zhu, Y.; Zhang, M. Determination of deoxynivalenol-3-glucoside in cereals by hydrophilic interaction chromatography with ultraviolet detection. Food Anal. Methods 2014, 7, 1139–1146. [Google Scholar] [CrossRef]
- Trombete, F.; Barros, A.; Vieira, M.; Saldanha, T.; Venâncio, A.; Fraga, M. Simultaneous determination of deoxynivalenol, deoxynivalenol-3-glucoside and nivalenol in wheat grains by HPLC-PDA with immunoaffinity column cleanup. Food Anal. Methods 2016, 9, 2579–2586. [Google Scholar] [CrossRef]
- Bryła, M.; Ksieniewicz-Woźniak, E.; Waśkiewicz, A.; Szymczyk, K.; Jędrzejczak, R. Natural occurrence of nivalenol, deoxynivalenol, and deoxynivalenol-3-glucoside in Polish winter wheat. Toxins 2018, 10, 81. [Google Scholar] [CrossRef] [PubMed]
- Trenberth, K.E.; Caron, J.M. The Southern Oscillation Revisited: Sea Level Pressures, Surface, Temperatures, and Precipitation. J. Clim. 2000, 13, 4358–4365. [Google Scholar] [CrossRef]
- Zhang, H.; Van der Lee, T.; Waalwijk, C.; Chen, W.; Xu, J.; Zhang, Y.; Feng, J. Population analysis of the Fusarium graminearum species complex from wheat in China show a shift to more aggressive isolates. PLoS ONE 2012, 7, e31722. [Google Scholar] [CrossRef] [PubMed]
- Steinmüller, R.; Ziller, K.; Frenzel, W. Ein Korn sieht Rot: Besatzanalyse oder Mykotoxin Nachweis. Mühle Mischfutter 2004, 141, 377–382. [Google Scholar]
- Beyer, M.; Pogoda, F.; Pallez, M.; Lazic, J.; Hoffmann, L.; Pasquali, M. Evidence for a reversible drought induced shift in the species composition of mycotoxin producing Fusarium head blight pathogens isolated from symptomatic wheat heads. Int. J. Food Microbiol. 2014, 182–183, 51–56. [Google Scholar] [CrossRef]
- Lindblad, M.; Gidlund, A.; Sulyok, M.; Börjesson, T.; Krska, R.; Olsen, M.; Fredlund, E. Deoxynivalenol and other selected Fusarium toxins in Swedish wheat—Occurrence and correlation to specific Fusarium species. Int. J. Food Microbiol. 2013, 167, 284–291. [Google Scholar] [CrossRef]
- Juan, C.; Ritieni, A.; Mañes, J. Occurrence of Fusarium mycotoxins in Italian cereal and cereal products from organic farming. Food Chem. 2013, 141, 1747–1755. [Google Scholar] [CrossRef]
- Alkadri, D.; Rubert, J.; Prodi, A.; Pisi, A.; Manes, J.; Soler, C. Natural co-occurrence of mycotoxins in wheat grains from Italy and Syria. Food Chem. 2014, 157, 111–118. [Google Scholar] [CrossRef]
- Rafai, P.; Bata, A.; Jakab, L.; Vanyi, A. Evaluation of mycotoxin contaminated cereals for their use in animal feeds in Hungary. Food Addit. Contam. 2000, 17, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Schollenberger, M.; Jara, H.T.; Suchy, S.; Drochner, W.; Müler, H.M. Fusarium toxins in wheat flour collected in an area in southwest Germany. Int. J. Food Microbiol. 2002, 72, 85–89. [Google Scholar] [CrossRef]
- Edwards, S.G. Fusarium mycotoxin content of UK organic and conventional wheat. Food Addit. Contam. A 2009, 26, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Hajšlova, J.; Lancova, K.; Sehnalova, M.; Krplova, A.; Zachariašova, M.; Moravcova, H.; Nedělnik, J.; Markova, J.; Ehrenbergerova, J. Occurrence of trichothecene mycotoxins in cereals harvested in the Czech Republic. Czech J. Food Sci. 2007, 25, 339–350. [Google Scholar] [CrossRef]
- Pleadin, J.; Vahčić, N.; Perši, N.; Ševelj, D.; Markov, K.; Frece, J. Fusarium mycotoxins’ occurrence in cereals harvested from Croatian fields. Food Control 2013, 32, 49–54. [Google Scholar] [CrossRef]
- Fels-Klerx, H.J.V.D.; Klemsdal, S.; Hietaniemi, V.; Lindblad, M.; Ioannou-Kakouri, E.; Asselt, E.D.V. Mycotoxin contamination of cereal grain commodities in relation to climate in North West Europe. Food Addit. Contam. Part A 2012, 29, 1581–1592. [Google Scholar] [CrossRef] [PubMed]
- Vogelgsang, S.; Musa, T.; Bänziger, I.; Kägi, A.; Bucheli, T.D.; Wettstein, F.E.; Pasquali, M.; Forrer, H.-R. Fusarium Mycotoxins in Swiss Wheat: A survey of growers’ samples between 2007 and 2014 shows strong year and minor geographic effects. Toxins 2017, 9, 246. [Google Scholar] [CrossRef]
- Yoshizawa, T. Thirty-five years of research on deoxynivalenol, a trichothecene mycotoxin: With special reference to its discovery and co-occurrence with nivalenol in Japan thirty-five years of research on deoxynivalenol in Japan. Food Saf. 2013, 1, 12–31. [Google Scholar]
- Mirocha, C.J.; Abbas, H.K.; Windels, C.E.; Xie, W. Variation in deoxynivalenol, 15-acetyl-deoxynivalenol, 3-acetyl-deoxynivalenol and zearalenone production by Fusarium graminearum isolates. Appl. Environ. Microbiol. 1989, 55, 1315–1316. [Google Scholar]
- Bakan, B.; Giraud-Deville, C.; Pinson, L.; Richard-Molard, D.; Fournier, E.; Brygoo, Y. Identification by PCR of Fusarium culmorum strains producing large and small amounts of deoxynivalenol. Appl. Environ. Microbiol. 2002, 68, 5472–5479. [Google Scholar] [CrossRef]
- Walker, S.L.; Leath, S.; Hagler, W.M.; Murphy, J.P. Variation among isolates of Fusarium graminearum associated with Fusarium head bligh in north Carolinia. Plant Dis. 2001, 85, 404–410. [Google Scholar] [CrossRef]
- Champeil, A.; Doré, T.; Fourbet, J.F. Fusarium head blight: Epidemiological origin of the effects of cultural practices on head blight attacks and the production of mycotoxins by Fusarium in wheat grains. Plant Sci. 2004, 166, 1389–1415. [Google Scholar] [CrossRef]
- Shou, X.; Kleijer, G. Effects of Fusarium metabolites on growth of callus and seedling in triticale (abstract). Acta Prataculturae Sin. 2001, 10, 78–85. [Google Scholar]
- Audenaert, K.; Callewaert, E.; Höfte, M.; Saeger, S.D.; Haesaert, G. Hydrogen peroxide induced by the fungicide prothioconazole triggers deoxynivalenol (DON) production by Fusarium graminearum. BMC Microbiol. 2010, 10, 112. [Google Scholar] [CrossRef] [PubMed]
- Marín, P.; de Ory, A.; Cruz, A.; Magan, N.; González-Jaén, M.T. Potential effects of environmental conditions on the efficiency of the antifungal tebuconazole controlling Fusarium verticillioides and Fusarium proliferatum growth rate and fumonisin biosynthesis. Int. J. Food Microbiol. 2013, 165, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Moya-Elizondo, E.A.; Jacobsen, B.J. Integrated management of Fusarium crown rot of wheat using fungicide seed treatment, cultivar resistance, and induction of systemic acquired resistance (SAR). Biol. Control 2016, 92, 153–163. [Google Scholar] [CrossRef]
- Podolska, G.; Bryła, M.; Sułek, A.; Waśkiewicz, A.; Szymczyk, K.; Jędrzejczak, R. Influence of the cultivar and nitrogen fertilisation level on the mycotoxin contamination in winter wheat. Qual. Assur. Saf. Crop. 2017, 9, 451–461. [Google Scholar] [CrossRef]
- Desmarchelier, A.; Seefelder, W. Survey of deoxynivalenol and deoxynivalenol-3-glucoside in cereal-based products by liquid chromatography electrospray ionization tandem mass spectrometry. World Mycotoxin J. 2011, 4, 29–35. [Google Scholar] [CrossRef]
- Palacios, S.A.; Erazo, J.G.; Ciasca, B.; Lattanzio, V.M.T.; Reynoso, M.M.; Farnochi, M.C.; Torres, A.M. Occurrence of deoxynivalenol and deoxynivalenol-3-glucoside in durum wheat from Argentina. Food Chem. 2017, 230, 728–734. [Google Scholar] [CrossRef]
- Dall’Asta, C.; Dall’Erta, A.; Mantovani, P.; Massi, A.; Galaverna, G. Occurrence of deoxynivalenol and deoxynivalenol-3-glucoside in durum wheat. World Mycotoxin J. 2013, 6, 83–91. [Google Scholar] [CrossRef]
- Dong, F.; Wang, S.; Yu, M.; Sun, Y.; Xu, J.; Shi, J. Natural occurrence of deoxynivalenol and deoxynivalenol-3-glucoside in various wheat cultivars grown in Jiangsu province, China. World Mycotoxin J. 2017, 10, 285–293. [Google Scholar] [CrossRef]
- Ovando-Martínez, M.; Ozsisli, B.; Anderson, J.; Whitney, K.; Ohm, J.-B.; Simsek, S. Analysis of deoxynivalenol and deoxynivalenol-3-glucoside in hard red spring wheat inoculated with Fusarium graminearum. Toxins 2013, 5, 2522–2532. [Google Scholar] [CrossRef] [PubMed]
- Amarasinghe, C.C.; Simsek, S.; Brule-Babel, A.; Fernando, W.G.D. Analysis of deoxynivalenol and deoxynivalenol-3-glucosides content in Canadian spring wheat cultivars inoculated with Fusarium graminearum. Food Addit. Contam. A 2016, 33, 1254–1264. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Recovery (R) and Relative Standard Deviation (RSD) | NIV | NIV-3G | DON | DON-3G | 3-AcDON | 15-AcDON |
|---|---|---|---|---|---|---|
| fortification level for n = 4 (µg/kg) | 88.5 | 52.9 | 96.2 | 9.7 | 97.8 | 97.8 |
| R (%) | 71.7 | 85.0 | 88.9 | 104.3 | 70.5 | <10 |
| RSD (%) | 12.8 | 20.2 | 19.8 | 19.0 | 7.2 | - |
| fortification level for n = 4 (µg/kg) | 176.9 | 105.8 | 192.3 | 19.3 | 195.6 | 195.6 |
| R (%) | 77.6 | 83.5 | 98.0 | 88.0 | 71.8 | <10 |
| RSD (%) | 8.9 | 9.4 | 12.5 | 13.2 | 7.3 | - |
| fortification level for n = 4 (µg/kg) | 265.4 | 158.6 | 288.5 | 29.0 | 293.4 | 293.4 |
| R (%) | 81.3 | 91.4 | 97.2 | 92.1 | 72.7 | <10 |
| RSD (%) | 11.3 | 12.9 | 9.5 | 11.1 | 10.4 | - |
| fortification level for n = 4 (µg/kg) | 530.7 | 317.3 | 576.9 | 57.9 | 586.8 | 586.8 |
| R (%) | 78.3 | 82.7 | 99.9 | 84.5 | 69.5 | <10 |
| RSD (%) | 5.7 | 4.5 | 7.8 | 12.1 | 7.3 | - |
| Wheat Samples | Concentration (μg/kg) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NIV | NIV-3G | NIV-3G/NIV Molar Ratio | DON | DON-3G | DON-3G/DON Molar Ratio | 3-AcDON | NIV | NIV-3G | NIV-3G/NIV Molar Ratio | DON | DON-3G | DON-3G/DON Molar Ratio | 3-AcDON | |||
| Season 2017 | Season 2018 | |||||||||||||||
| Northern Poland n = 27 in 2017 n = 30 in 2018 | Positive samples (%) | 12 (44%) | 5 (19%) | - | 27 (100%) | 21 (78%) | - | 9 (33%) | 12 (40%) | 3 (10%) | 15 (50%) | 8 (27%) | 4 (13%) | |||
| Average | 16.4 ab | 8.2 a | 20% | 143.3 a | 37.5 a | 17% | 3.2 a | 11.8 a | 5.7 a | 19% | 11.4 a | 5.4 a | 23% | 3.8 ab | ||
| Median | 13.3 | 7.0 | 22% | 75.5 | 30.1 | 15% | 3.2 | 8.6 | 5.2 | 19% | 9.5 | 4.7 | 25% | 3.8 | ||
| Min–Max | 8.4–45.0 | 5.0–14.7 | 20–36% | 18.3–515.1 | 5.6–107.6 | 5–34% | 2.3–4.0 | 8.0–22.5 | 5.0–6.8 | 18–20% | 5.1–29.7 | 4.2–8.6 | 13–30% | 3.0–4.4 | ||
| Western Poland n = 17 in 2017 n = 17 in 2018 | Positive samples (%) | 3 (18%) | - | - | 16 (94%) | 13 (76%) | - | 3 (18%) | 10 (59%) | 3 (18%) | 6 (35%) | 1 (6%) | 1 (6%) | |||
| Average | 11.5 ab | - | - | 77.9 a | 18.6 ab | 14% | 5.7 b | 14.5 ab | 5.9 a | 17% | 7.8 ab | 4.7 | 34% | 2.3 | ||
| Median | 11.1 | - | - | 69.1 | 14.9 | 14% | 5.8 | 11.1 | 5.2 | 20% | 8.3 | 4.7 | 34% | 2.3 | ||
| Min–Max | 10.8–12.7 | - | - | 9.9–148.1 | 4.4–43.1 | 8–21% | 2.5–8.9 | 8.0-26.6 | 5.0–7.4 | 12–20% | 5.1–10.7 | - | - | 2.3 | ||
| Central Poland n = 29 in 2017 n =26 in 2018 | Positive samples (%) | 17 (59%) | 6 (21%) | - | 28 (97%) | 19 (66%) | - | 10 (34%) | 13 (50%) | 4 (15%) | 18 (69%) | 9 (35%) | 5 (19%) | |||
| Average | 19.1 ab | 8.8 a | 20% | 164.5 a | 35.2 ab | 15% | 3.6 a | 51.0 b | 16.5 a | 13% | 15.5 ab | 5.6 a | 21% | 6.5 a | ||
| Median | 15.6 | 7.5 | 20% | 81.5 | 16.5 | 13% | 3.1 | 11.5 | 10.6 | 12% | 9.2 | 4.4 | 21% | 4.9 | ||
| Min–Max | 8.1–73.7 | 6.2–13.5 | 12–25% | 7.4–1260.9 | 4.4–150.3 | 3–34% | 2.4–6.8 | 8.0–405.4 | 5.0-39.6 | 6–22% | 5.3–57.1 | 4.0–10.2 | 7–54% | 2.9–16.1 | ||
| Eastern Poland n = 24 in 2017 n = 24 in 2018 | Positive samples (%) | 17 (71%) | 4 (17%) | - | 19 (79%) | 9 (38%) | - | 5 (21%) | 19 (79%) | 5 (21%) | 16 (67%) | 6 (25%) | 2 (8%) | |||
| Average | 14.6 a | 7.2 a | 22% | 111.4 a | 29.5 ab | 11% | 2.7 a | 22.6 ab | 7.0 a | 15% | 30.2 ab | 18.2 a | 24% | 3.7 ab | ||
| Median | 10.8 | 7.5 | 20% | 18.5 | 6.2 | 11% | 2.6 | 20.9 | 6.7 | 16% | 7.6 | 5.7 | 21% | 3.7 | ||
| Min–Max | 8.1–34.3 | 6.0–7.9 | 15–35% | 5.2–1670.7 | 4.0–217.2 | 6–19% | 2.0-3.4 | 8.2–74.5 | 6.3–8.1 | 6–19% | 5.0–303.1 | 4.0–82.1 | 10–40% | 2.4–4.9 | ||
| Southern Poland n = 53 in 2017 n = 53 in 2018 | Positive samples (%) | 24 (45%) | 5 (9%) | - | 28 (53%) | 22 (42%) | - | 6 (11%) | 36 (68%) | 5 (9%) | 22 (42%) | 17 (32%) | 17 (32%) | 7 (13%) | ||
| Average | 31.5 b | 10.7 a | 11% | 91.7 a | 18.4 b | 14% | 4.6 ab | 19.7 ab | 8.0 a | 15% | 55.1 b | 15.1 a | 21% | 3.3 b | ||
| Median | 20.2 | 7.3 | 10% | 39.5 | 10.4 | 14% | 4.0 | 16.6 | 6.9 | 14% | 13.7 | 4.9 | 21% | 2.7 | ||
| Min–Max | 8.5–185.6 | 5.5–26.8 | 7–14% | 5.8–1428.7 | 4.0–199.6 | 7–27% | 2.5–9.0 | 8.0–61.9 | 5.9–11.0 | 11–22% | 5.0–461.7 | 4.0–87.7 | 10–31% | 2.4 | ||
| TOTAL | Positive samples (%) | 73 (49%) | 22 (15%) | - | 138 (92%) | 97 (65%) | - | 38 (25%) | 89 (59%) | 20 (13%) | 91 (61%) | 50 (33%) | 24 (16%) | |||
| Average | 21.4 a | 8.7 a | 19.5% | 109.2 a | 25.8 a | 15% | 3.6 a | 23.4 a | 8.8 a | 15.5% | 32.4 b | 12.3 b | 21.9% | 3.9 a | ||
| Median | 15.0 | 7.4 | 20.0% | 45.7 | 11.6 | 14% | 3.2 | 14.6 | 6.7 | 15.8% | 10.0 | 4.8 | 21.3% | 3.0 | ||
| Min–Max | 8.1–185.6 | 5.0–26.8 | 7–36% | 5.2–1670.7 | 4.0–217.2 | 3–34% | 2.0–9.0 | 8.0–405.4 | 5.0–39.6 | 6–22% | 5.0–461.7 | 4.0–87.7 | 7–54% | 2.2–16.1 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bryła, M.; Ksieniewicz-Woźniak, E.; Yoshinari, T.; Waśkiewicz, A.; Szymczyk, K. Contamination of Wheat Cultivated in Various Regions of Poland during 2017 and 2018 Agricultural Seasons with Selected Trichothecenes and Their Modified Forms. Toxins 2019, 11, 88. https://doi.org/10.3390/toxins11020088
Bryła M, Ksieniewicz-Woźniak E, Yoshinari T, Waśkiewicz A, Szymczyk K. Contamination of Wheat Cultivated in Various Regions of Poland during 2017 and 2018 Agricultural Seasons with Selected Trichothecenes and Their Modified Forms. Toxins. 2019; 11(2):88. https://doi.org/10.3390/toxins11020088
Chicago/Turabian StyleBryła, Marcin, Edyta Ksieniewicz-Woźniak, Tomoya Yoshinari, Agnieszka Waśkiewicz, and Krystyna Szymczyk. 2019. "Contamination of Wheat Cultivated in Various Regions of Poland during 2017 and 2018 Agricultural Seasons with Selected Trichothecenes and Their Modified Forms" Toxins 11, no. 2: 88. https://doi.org/10.3390/toxins11020088
APA StyleBryła, M., Ksieniewicz-Woźniak, E., Yoshinari, T., Waśkiewicz, A., & Szymczyk, K. (2019). Contamination of Wheat Cultivated in Various Regions of Poland during 2017 and 2018 Agricultural Seasons with Selected Trichothecenes and Their Modified Forms. Toxins, 11(2), 88. https://doi.org/10.3390/toxins11020088