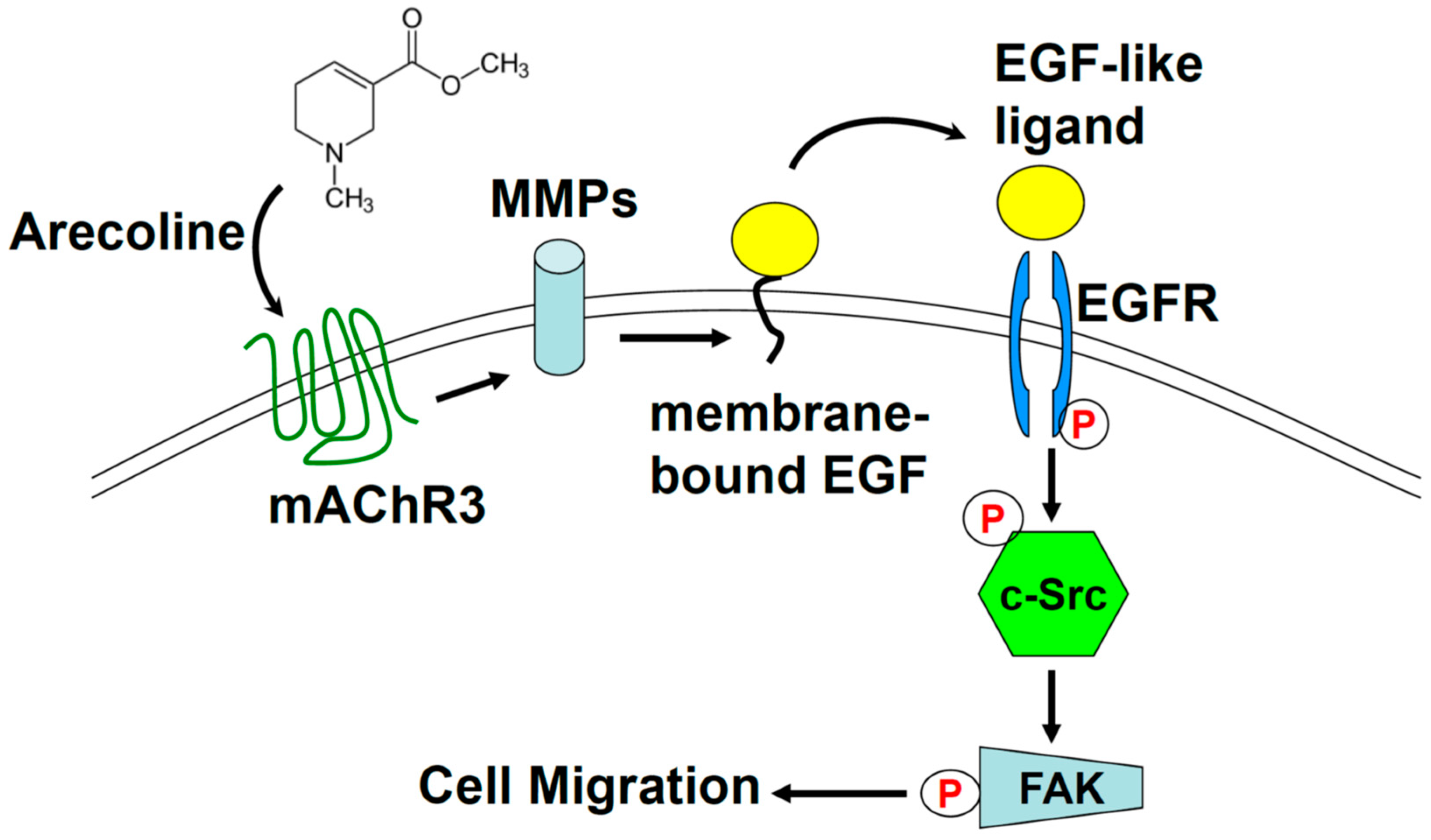
Arecoline Promotes Migration of A549 Lung Cancer Cells through Activating the EGFR/Src/FAK Pathway

 , , , and
, , , and 
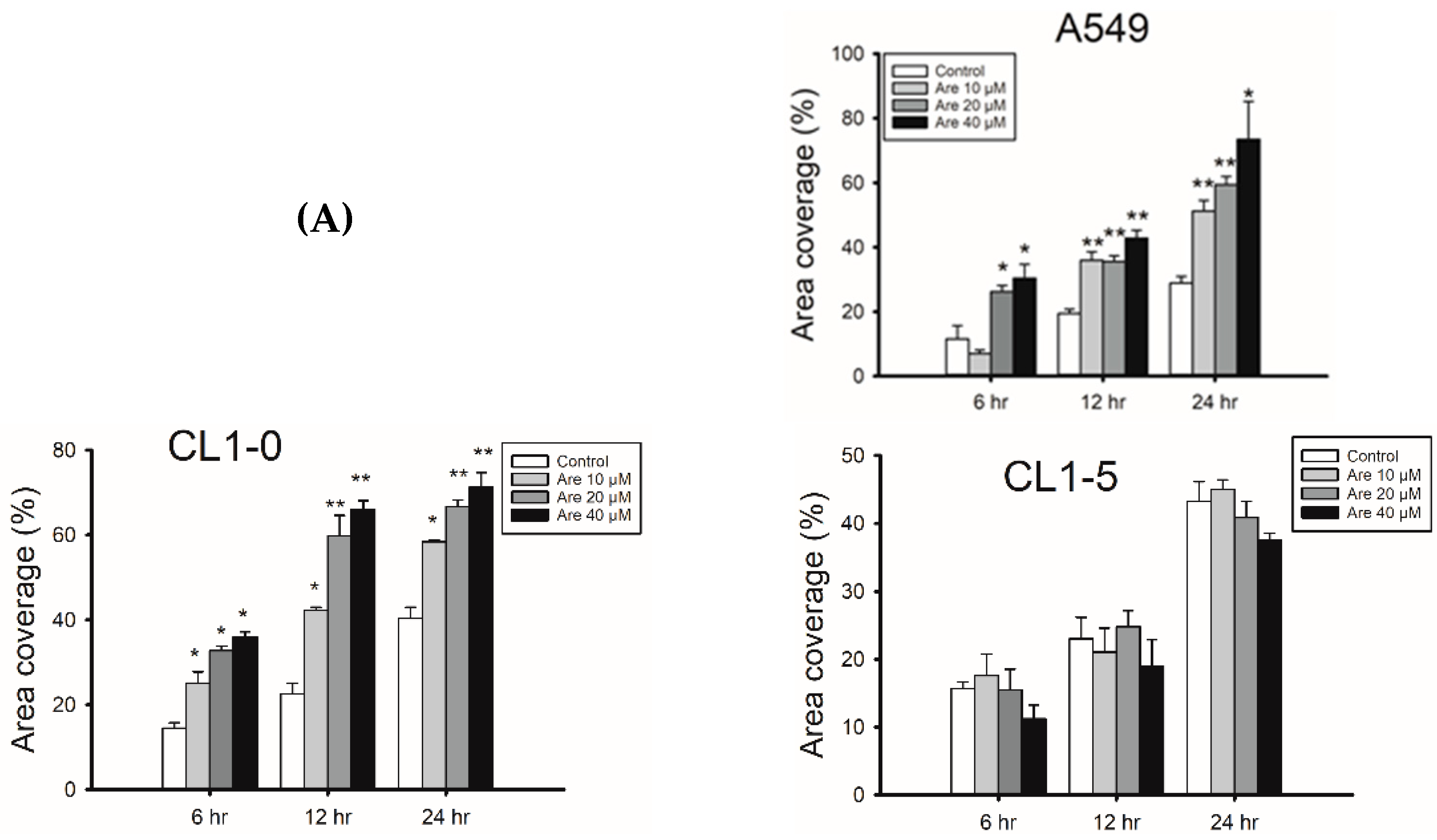{kind=link}
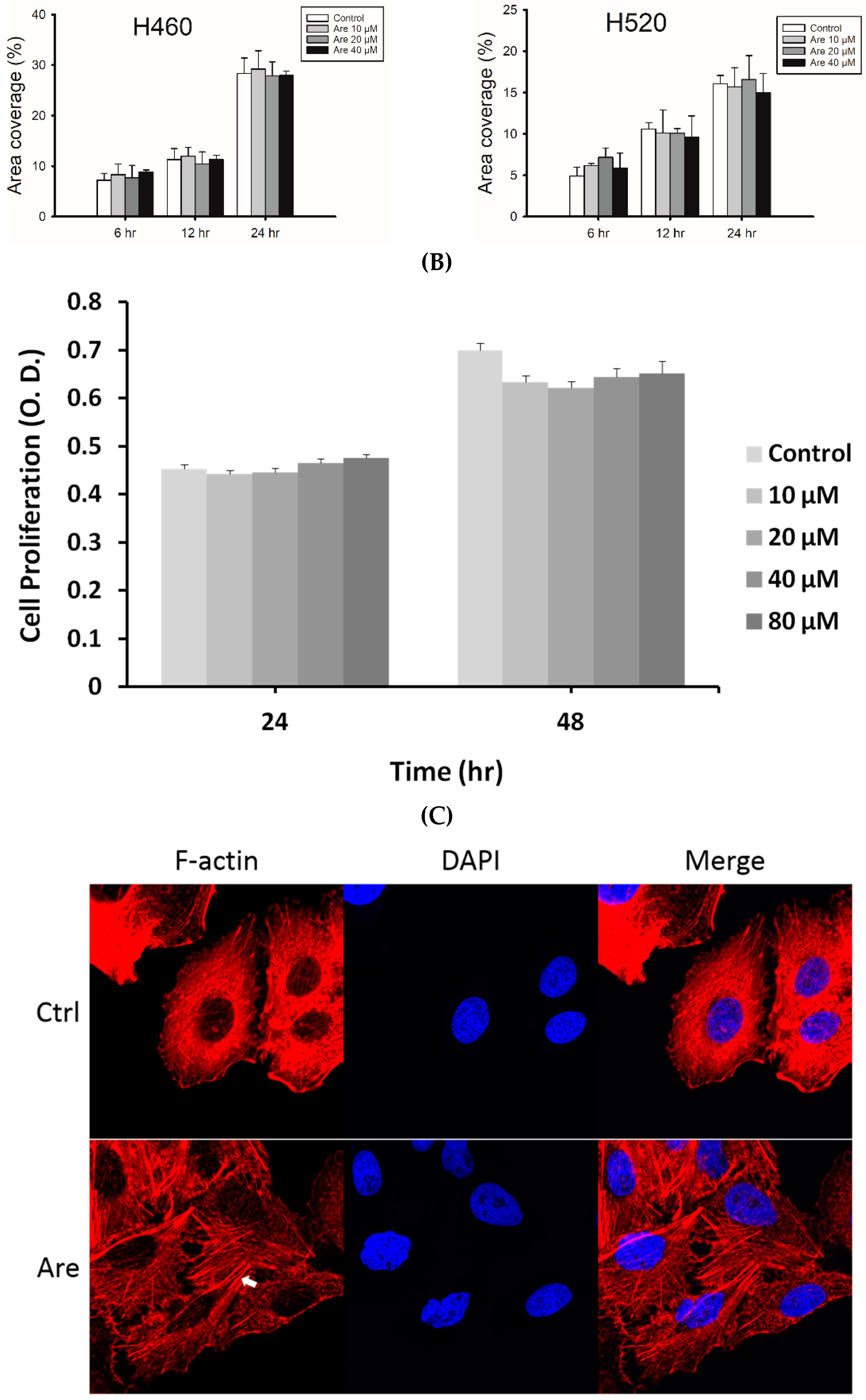{kind=link}
{kind=link}
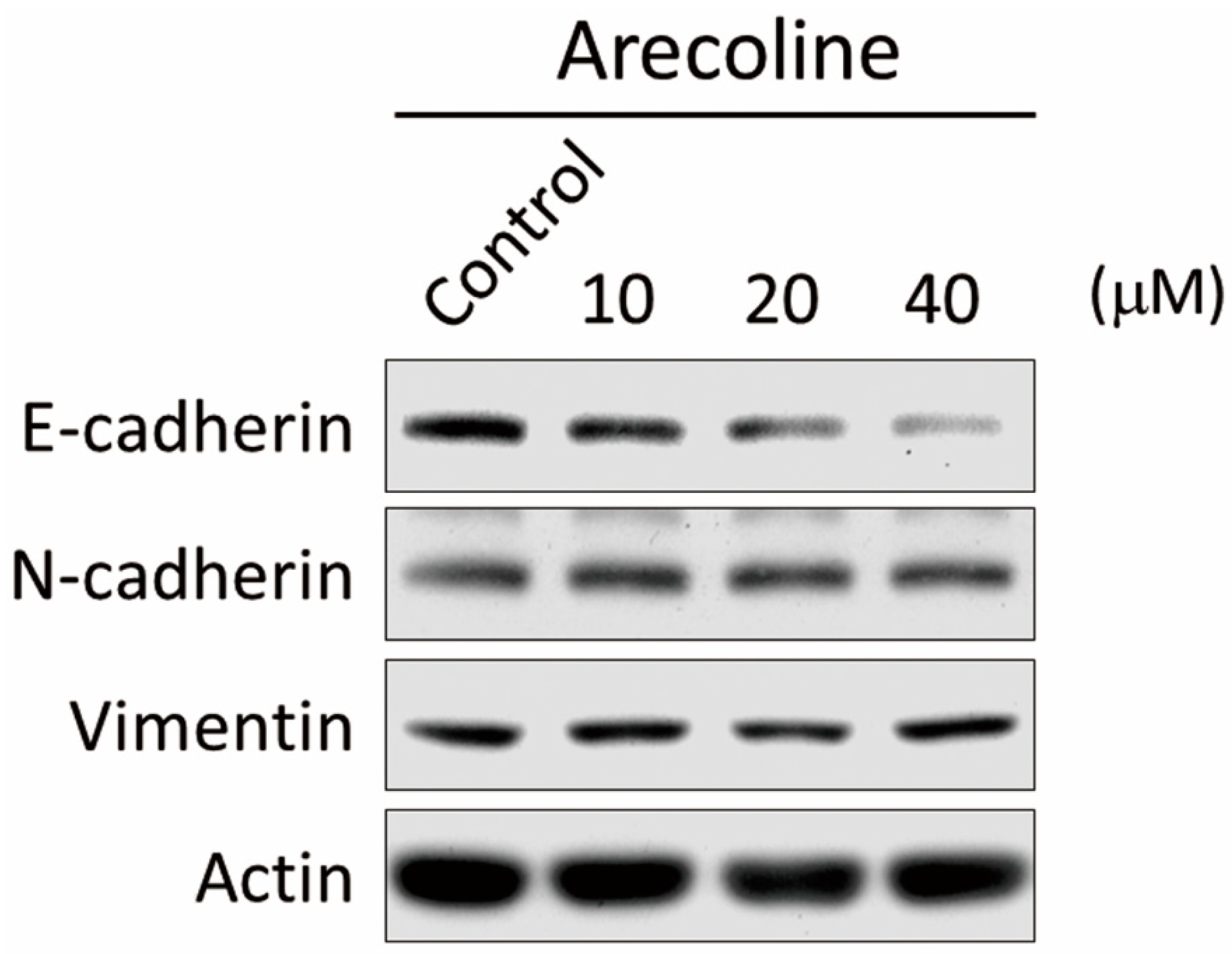{kind=link}
{kind=link}
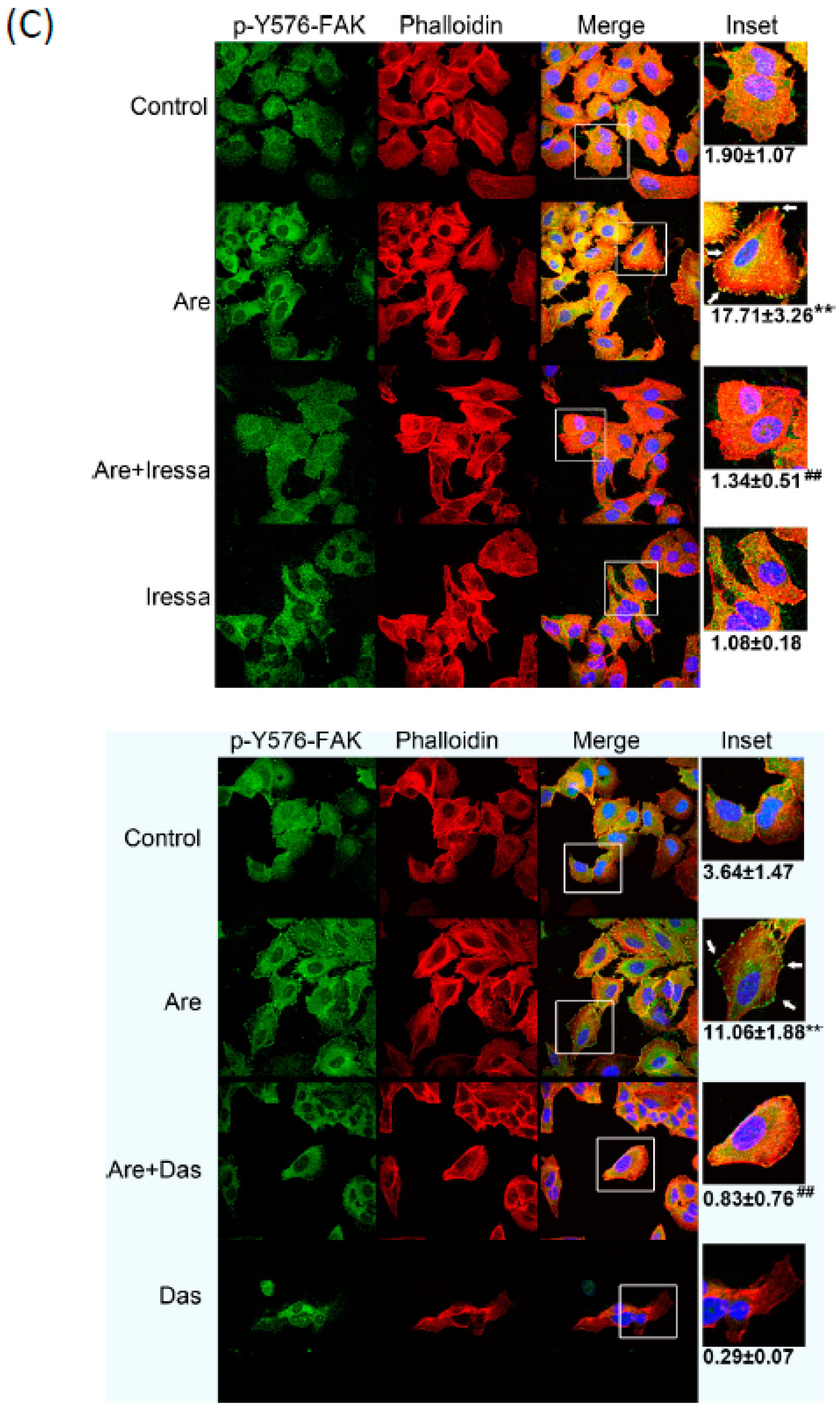{kind=link}
{kind=link}
{kind=link}
Abstract
Share and Cite
Chang, C.-H.; Chen, M.-C.; Chiu, T.-H.; Li, Y.-H.; Yu, W.-C.; Liao, W.-L.; Oner, M.; Yu, C.-T.R.; Wu, C.-C.; Yang, T.-Y.; et al. Arecoline Promotes Migration of A549 Lung Cancer Cells through Activating the EGFR/Src/FAK Pathway. Toxins 2019, 11, 185. https://doi.org/10.3390/toxins11040185
Chang C-H, Chen M-C, Chiu T-H, Li Y-H, Yu W-C, Liao W-L, Oner M, Yu C-TR, Wu C-C, Yang T-Y, et al. Arecoline Promotes Migration of A549 Lung Cancer Cells through Activating the EGFR/Src/FAK Pathway. Toxins. 2019; 11(4):185. https://doi.org/10.3390/toxins11040185
Chicago/Turabian StyleChang, Chih-Hsiang, Mei-Chih Chen, Te-Huan Chiu, Yu-Hsuan Li, Wan-Chen Yu, Wan-Ling Liao, Muhammet Oner, Chang-Tze Ricky Yu, Chun-Chi Wu, Tsung-Ying Yang, and et al. 2019. "Arecoline Promotes Migration of A549 Lung Cancer Cells through Activating the EGFR/Src/FAK Pathway" Toxins 11, no. 4: 185. https://doi.org/10.3390/toxins11040185
APA StyleChang, C.-H., Chen, M.-C., Chiu, T.-H., Li, Y.-H., Yu, W.-C., Liao, W.-L., Oner, M., Yu, C.-T. R., Wu, C.-C., Yang, T.-Y., Teng, C.-L. J., Chiu, K.-Y., Chen, K.-C., Wang, H.-Y., Yue, C.-H., Lai, C.-H., Hsieh, J.-T., & Lin, H. (2019). Arecoline Promotes Migration of A549 Lung Cancer Cells through Activating the EGFR/Src/FAK Pathway. Toxins, 11(4), 185. https://doi.org/10.3390/toxins11040185