First Characterization of The Venom from Apis mellifera syriaca, A Honeybee from The Middle East Region


 ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
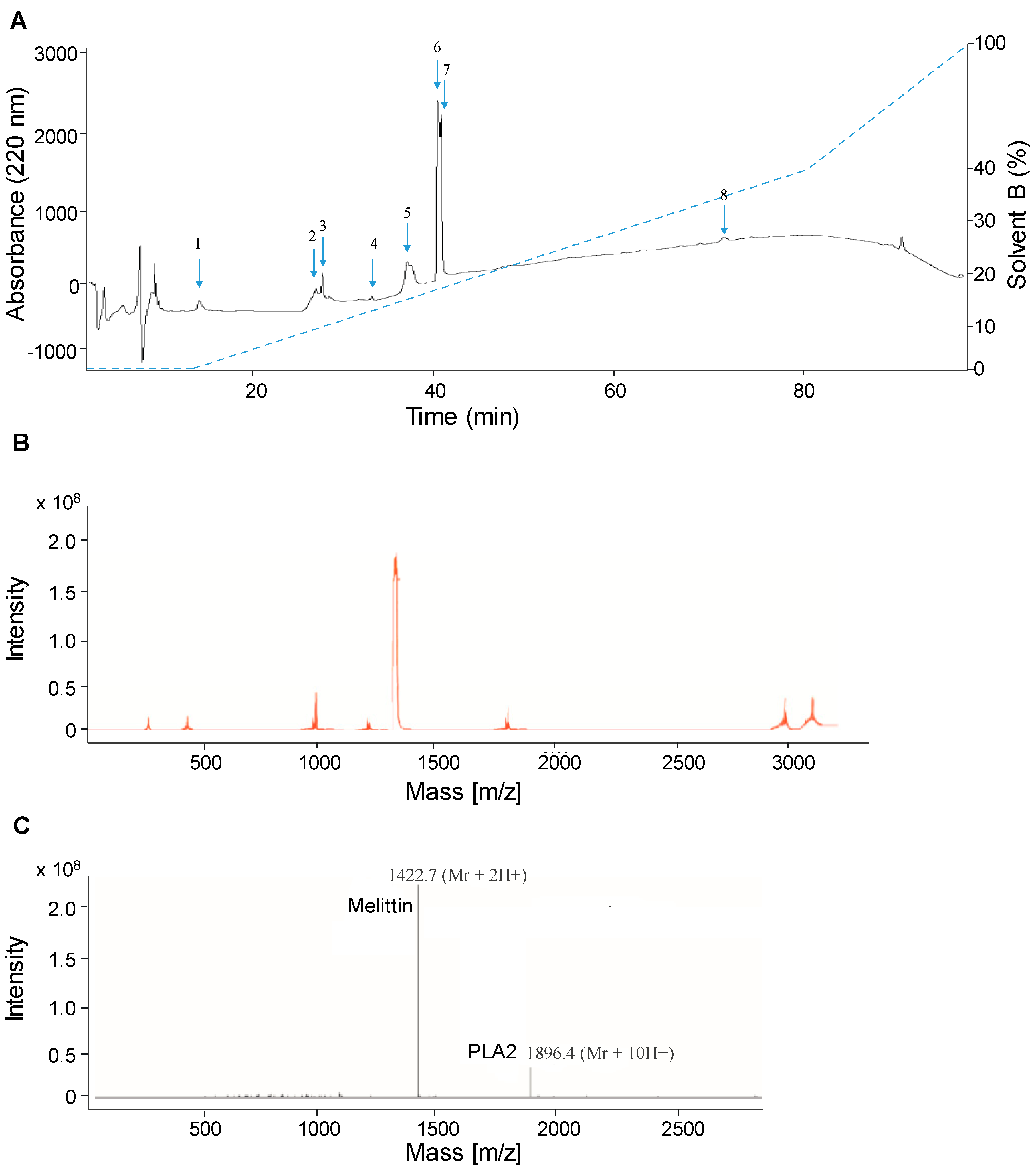
2.1. Separation and Analysis of Venom Compounds by LC-ESI-MS
2.2. Separation of Crude Venom Compounds by SPE
2.3. Antibacterial Activity
2.4. Hemolytic Activity
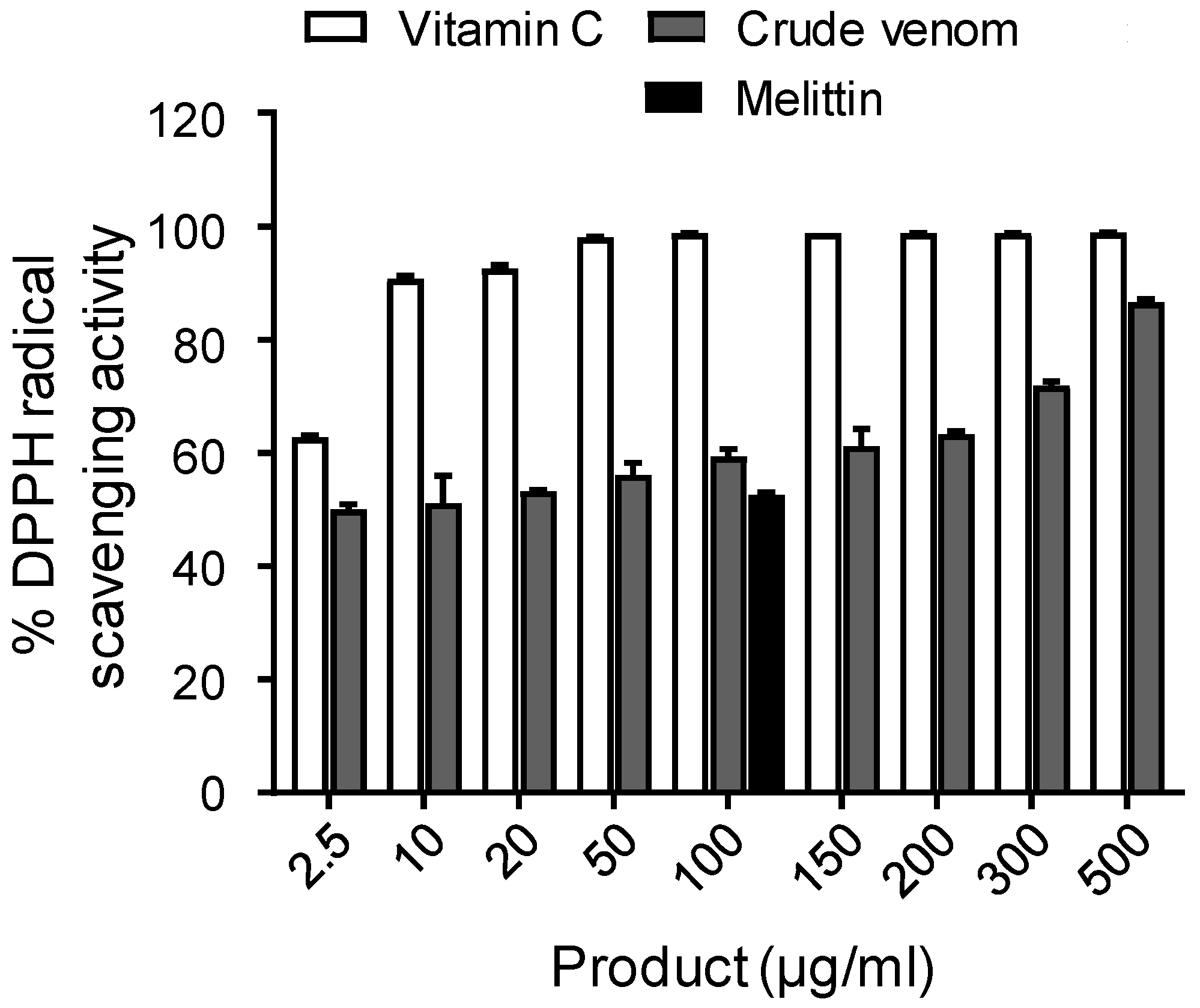
2.5. Antioxidant Activity
2.6. PLA2 Activity
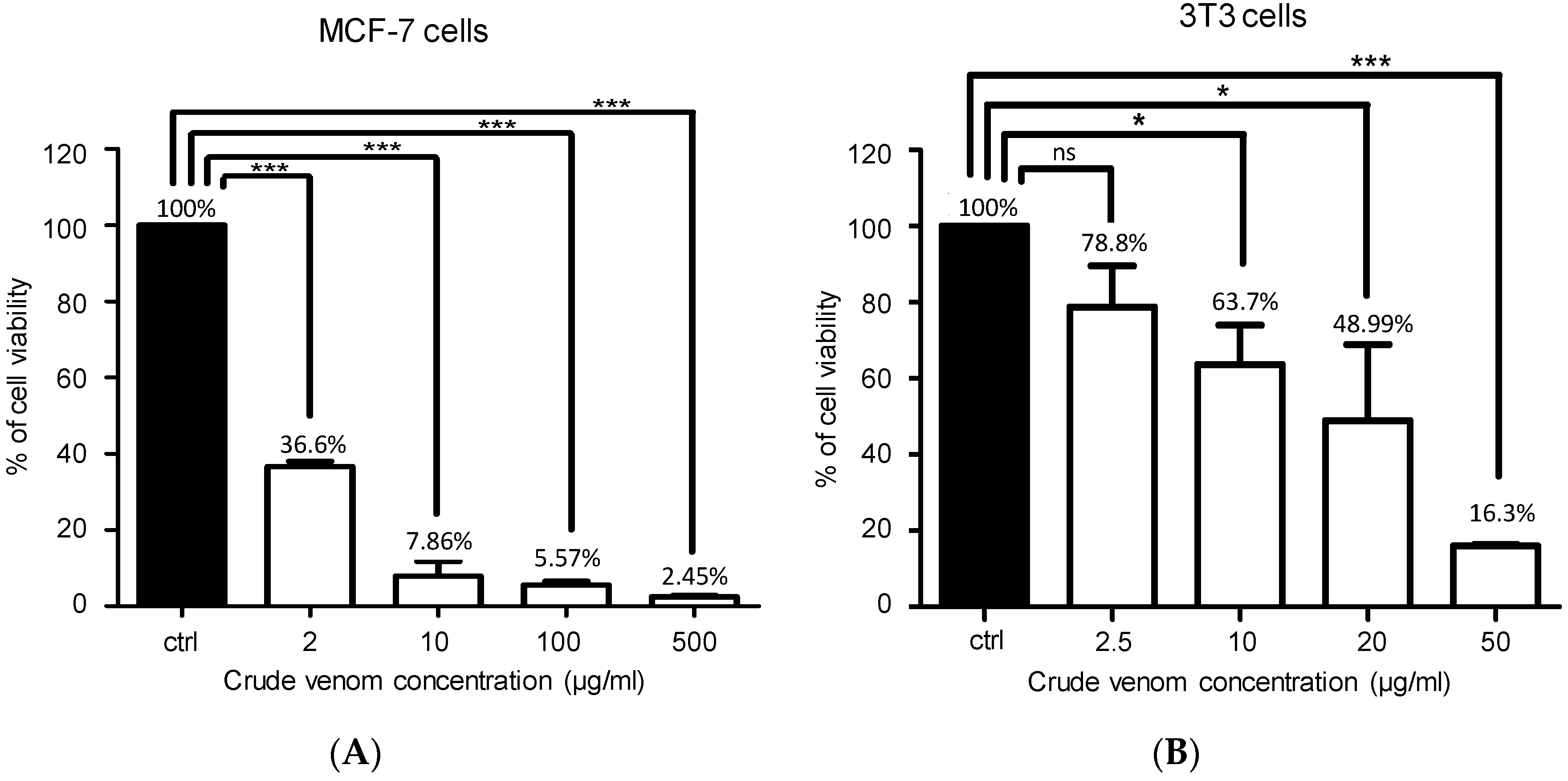
2.7. Cytotoxic Activity on MCF-7 and 3T3 Cancer Cells
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Materials
5.1.1. Bees
5.1.2. Venom
5.1.3. Reagents
5.2. Methods
5.2.1. Chemical Characterization of The Crude Venom by LC-ESI-MS
5.2.2. Separation of Crude Venom Compounds using Solid Phase Extraction (SPE)
5.2.3. Antibacterial Activity
5.2.4. Hemolytic Activity
5.2.5. Antioxidant Activity Assay
5.2.6. Measurement of PLA2 Activity
5.2.7. Cytotoxic Activity Assay on MCF-7 and 3T3 Cancer Cells
5.2.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, R.; Zhang, L.; Fang, Y.; Han, B.; Lu, X.; Zhou, T.; Feng, M.; Li, J. Proteome and phosphoproteome analysis of honeybee (Apis mellifera) venom collected from electrical stimulation and manual extraction of the venom gland. BMC Genom. 2013, 14, 766. [Google Scholar] [CrossRef] [PubMed]
- Sobral, F.; Sampaio, A.; Falcão, S.; Queiroz, M.J.R.; Calhelha, R.C.; Vilas-Boas, M.; Ferreira, I.C. Chemical characterization, antioxidant, anti-inflammatory and cytotoxic properties of bee venom collected in Northeast Portugal. Food Chem. Toxicol. 2016, 94, 172–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zolfagharian, H.; Mohajeri, M.; Babaie, M. Bee Venom (Apis Mellifera) an Effective Potential Alternative to Gentamicin for Specific Bacteria Strains. J. Pharm. 2016, 19, 225–230. [Google Scholar] [CrossRef]
- Matysiak, J.; Schmelzer, C.E.; Neubert, R.H.; Kokot, Z.J. Characterization of honeybee venom by MALDI-TOF and nanoESI-QqTOF mass spectrometry. J. Pharm. Biomed. Anal. 2011, 54, 273–278. [Google Scholar] [CrossRef]
- Han, S.; Lee, K.; Yeo, J.; Kim, W.; Park, K. Biological effects of treatment of an animal skin wound with honeybee (Apis melifera. L) venom. J. Plast. Reconstruct. Aesthet. Surg. 2011, 64, e67–e72. [Google Scholar] [CrossRef] [PubMed]
- Cherniack, E.P.; Govorushko, S. To bee or not to bee: The potential efficacy and safety of bee venom acupuncture in humans. Toxicon 2018, 154, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, Y.; Ye, Y.; Wang, X.R.; Lin, L.T.; Xiao, L.Y.; Zhou, P.; Shi, G.X.; Liu, C.Z. Bee venom therapy: Potential mechanisms and therapeutic applications. Toxicon 2018, 148, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Guan, S.M.; Sun, W.; Fu, H. Melittin, the Major Pain-Producing Substance of Bee Venom. Neuroscience 2016, 32, 265–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sah, P.; Faber, E.L. Channels underlying neuronal calcium-activated potassium currents. Prog. Neurobiol. 2002, 66, 345–353. [Google Scholar] [CrossRef]
- Alvarez-Fischer, D.; Noelker, C.; Vulinović, F.; Grünewald, A.; Chevarin, C.; Klein, C.; Oertel, W.H.; Hirsch, E.C.; Michel, P.P.; Hartmann, A. Bee Venom and Its Component Apamin as Neuroprotective Agents in a Parkinson Disease Mouse Model. PLoS ONE 2013, 8, e61700. [Google Scholar] [CrossRef]
- Bae, G.L.H. Bee Venom Phospholipase A2: Yesterday’s Enemy Becomes Today’s Friend. Toxins 2016, 8, 48. [Google Scholar] [Green Version]
- Pak, S.C. An Introduction to the Toxins Special Issue on “Bee and Wasp Venoms: Biological Characteristics and Therapeutic Application”. Toxins 2016, 8, 315. [Google Scholar] [CrossRef]
- Zaitoun, S.T.; Al-Ghzawi, A.M.; Shannag, H.K. Population dynamics of the Syrian Honeybee, Apis mellifera syriaca, under semi-arid Mediterranean conditions. Zool. Middle East 2000, 21, 129–132. [Google Scholar] [CrossRef]
- Haddad, N.; Mahmud Batainh, A.; Suleiman Migdadi, O.; Saini, D.; Krishnamurthy, V.; Parameswaran, S.; Alhamuri, Z. Next generation sequencing of Apis mellifera syriacaidentifies genes for Varroaresistance and beneficial beekeeping traits. Insect Sci. 2016, 23, 579–590. [Google Scholar] [CrossRef]
- Zakour, M.K.; Ehrhardt, K.; Bienefeld, K. First estimate of genetic parameters for the Syrian honeybeeApis mellifera syriaca. Apidologie 2012, 43, 600–607. [Google Scholar] [CrossRef]
- Chen, J.; Luo, C.; Li, H.L.; Chen, H.S. Primary hyperalgesia to mechanical and heat stimuli following subcutaneous bee venom injection into the plantar surface of hindpaw in the conscious rat: A comparative study with the formalin test. Pain 1999, 83, 67–76. [Google Scholar] [CrossRef]
- Teixeira, C.F.P.; Landucci, E.C.T.; Antunes, E.; Chacur, M.; Cury, Y. Inflammatory effects of snake venom myotoxic phospholipases A2. Toxicon 2003, 42, 947–962. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.L.; DeWitt, D.L.; Garavito, R.M. Cyclooxygenases: Structural, cellular, and molecular biology. Annu. Rev. Biochem. 2000, 69, 145–182. [Google Scholar] [CrossRef]
- Rybak-Chmielewska, H.; Szczêsna, T. HPLC study of chemical composition of honeybee (Apis mellifera L.) venom. J. Apicult. Sci. 2004, 48, 103–109. [Google Scholar]
- Giralt, M.M.A.E. Three valuable peptides from bee and wasp venoms for therapeutic and biotechnological use: Melittin, apamin and mastoparan. Toxins 2015, 7, 1126–1150. [Google Scholar]
- Loo, J.A. Studying non covalent protein complexes by electrospray ionization mass spectrometry. Mass Spectrom. Rev. 1997, 16, 1–23. [Google Scholar] [CrossRef]
- Camel, V.R. Solid phase extraction of trace elements. Spectrochim. Acta Part B 2003, 58, 1177–1233. [Google Scholar] [CrossRef]
- Leandro, L.F.; Mendes, C.A.; Casemiro, L.A.; Vinholis, A.H.; Cunha, W.R.; Almeida, R.D.; Martins, C.H. Antimicrobial activity of apitoxin, melittin and phospholipase A2 of honey bee (Apis mellifera) venom against oral pathogens. Anais Academia Brasileira Ciencias 2015, 87, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Fratini, F.; Cilia, G.; Turchi, B.; Felicioli, A. Insects, arachnids and centipedes venom: A powerful weapon against bacteria. Toxicon 2017, 130, 91–103. [Google Scholar] [CrossRef]
- Tosteson, M.T.; Holmes, S.J.; Razin, M.; Tosteson, D.C. Melittin Lysis of Red Cells. J. Membr. Biol. 1985, 87, 35–44. [Google Scholar] [CrossRef]
- Vetter, R.S.; Visscher, P.K.; Camazine, S. Mass Envenomations by Honey Bees and Wasps. West J. Med. 1999, 170, 223. [Google Scholar]
- CezaryWatala, J.K.K. Hemolytic potency and phospholipase activity of some bee and wasp venoms. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1990, 97, 187–194. [Google Scholar]
- Somwongin, S.; Chantawannakul, P.; Chaiyana, W. Antioxidant activity and irritation property of venoms from Apis species. Toxicon 2018, 145, 32–39. [Google Scholar] [CrossRef]
- Murakami, M.; Kudo, I. Phospholipase A2. J. Biochem. 2002, 131, 285–292. [Google Scholar] [CrossRef]
- Tusiimire, J.; Wallace, J.; Woods, N.; Dufton, M.; Parkinson, J.; Abbott, G.; Clements, C.; Young, L.; Park, J.; Jeon, J. Effect of bee venom and its fractions on the release of pro-inflammatory cytokines in PMA-differentiated U937 cells co-stimulated with LPS. Vaccines 2016, 4, 11. [Google Scholar] [CrossRef]
- Oršolić, N. Bee venom in cancer therapy. Cancer Metastasis Rev. 2012, 31, 173–194. [Google Scholar] [CrossRef] [PubMed]
- Premratanachai, P.; Chanchao, C. Review of the anticancer activities of bee products. Afr. J. Microbiol. Res. 2014, 4, 337–344. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, F.; Tan, J.; Peng, X.; Sun, L.; Wang, P.; Jia, S.; Yu, Q.; Huo, H.; Zhao, H. Melittin inhibits the invasion of MCF-7 cells by downregulating CD147 and MMP-9 expression. Oncol. Lett. 2017, 13, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Pence, R.J. Methods for producing and bio-assaying intact honeybee venom for medical use. Am. Bee J. 1981, 121, 726–731. [Google Scholar]
- Bauer, A.W.; Kirby, W.M.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Surendra, N.S.; Jayaram, G.N.; Reddy, M.S. Antimicrobial activity of crude venom extracts in honeybees (Apis cerana, Apis dorsata, Apis florea) tested against selected pathogens. Afr. J. Microbiol. Res. 2011, 5, 2765–2772. [Google Scholar]
- Accary, C.; Rima, M.; Kouzayha, A.; Hleihel, W.; Sadek, R.; Desfontis, J.C.; Fajloun, Z.; Hraoui-Bloquet, S. Effect of the Montivipera bornmuelleri snake venom on human blood: Coagulation disorders and hemolytic activities. Open J. Hematol. 2014, 5. [Google Scholar] [CrossRef]
- Accary, C.; Hraoui-Bloquet, S.; Hamze, M.; Sadek, R.; Hleihel, W.; Desfontis, J.-C.; Fajloun, Z. Preliminary proteomic analysis and biological characterization of the crude venom of Montivipera bornmuelleri; a viper from Lebanon. Recent Adv. Biomed. Chem. Eng. Mater. Sci. 2014, 1, 167–173. [Google Scholar]
- Amini, E.; Baharara, J.; Nikdel, N.; Salek Abdollahi, F. Cytotoxic and Pro-Apoptotic Effects of Honey Bee Venom and Chrysin on Human Ovarian Cancer Cells. Asia Pac. J. Med. Toxicol. 2015, 4, 68–73. [Google Scholar]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frangieh, J.; Salma, Y.; Haddad, K.; Mattei, C.; Legros, C.; Fajloun, Z.; El Obeid, D. First Characterization of The Venom from Apis mellifera syriaca, A Honeybee from The Middle East Region. Toxins 2019, 11, 191. https://doi.org/10.3390/toxins11040191
Frangieh J, Salma Y, Haddad K, Mattei C, Legros C, Fajloun Z, El Obeid D. First Characterization of The Venom from Apis mellifera syriaca, A Honeybee from The Middle East Region. Toxins. 2019; 11(4):191. https://doi.org/10.3390/toxins11040191
Chicago/Turabian StyleFrangieh, Jacinthe, Yahya Salma, Katia Haddad, Cesar Mattei, Christian Legros, Ziad Fajloun, and Dany El Obeid. 2019. "First Characterization of The Venom from Apis mellifera syriaca, A Honeybee from The Middle East Region" Toxins 11, no. 4: 191. https://doi.org/10.3390/toxins11040191