αD-Conotoxins in Species of the Eastern Pacific: The Case of Conus princeps from Mexico


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
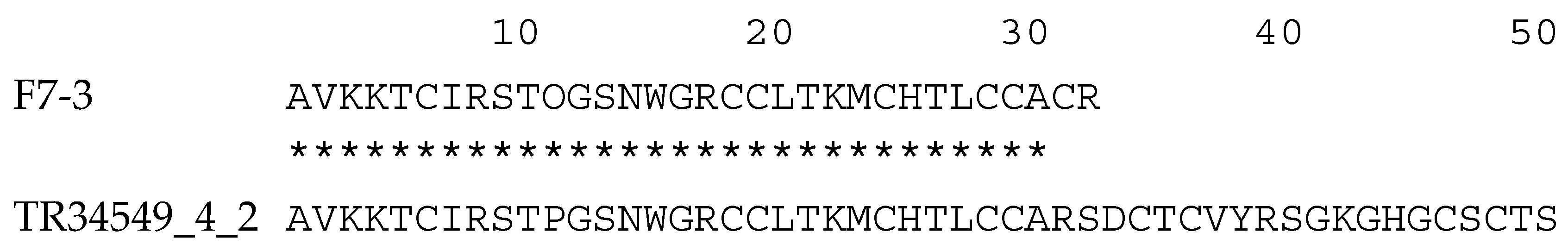{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
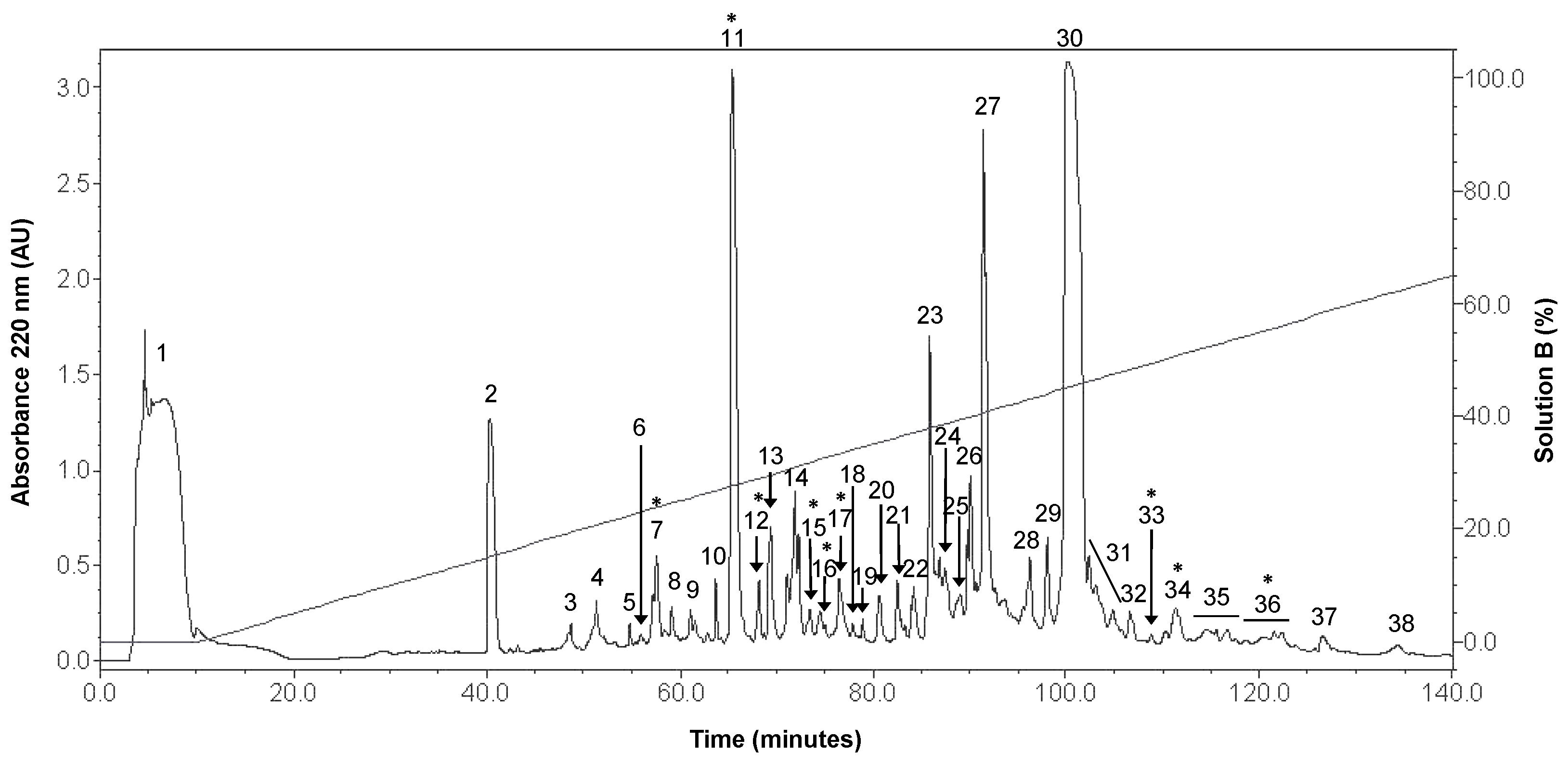
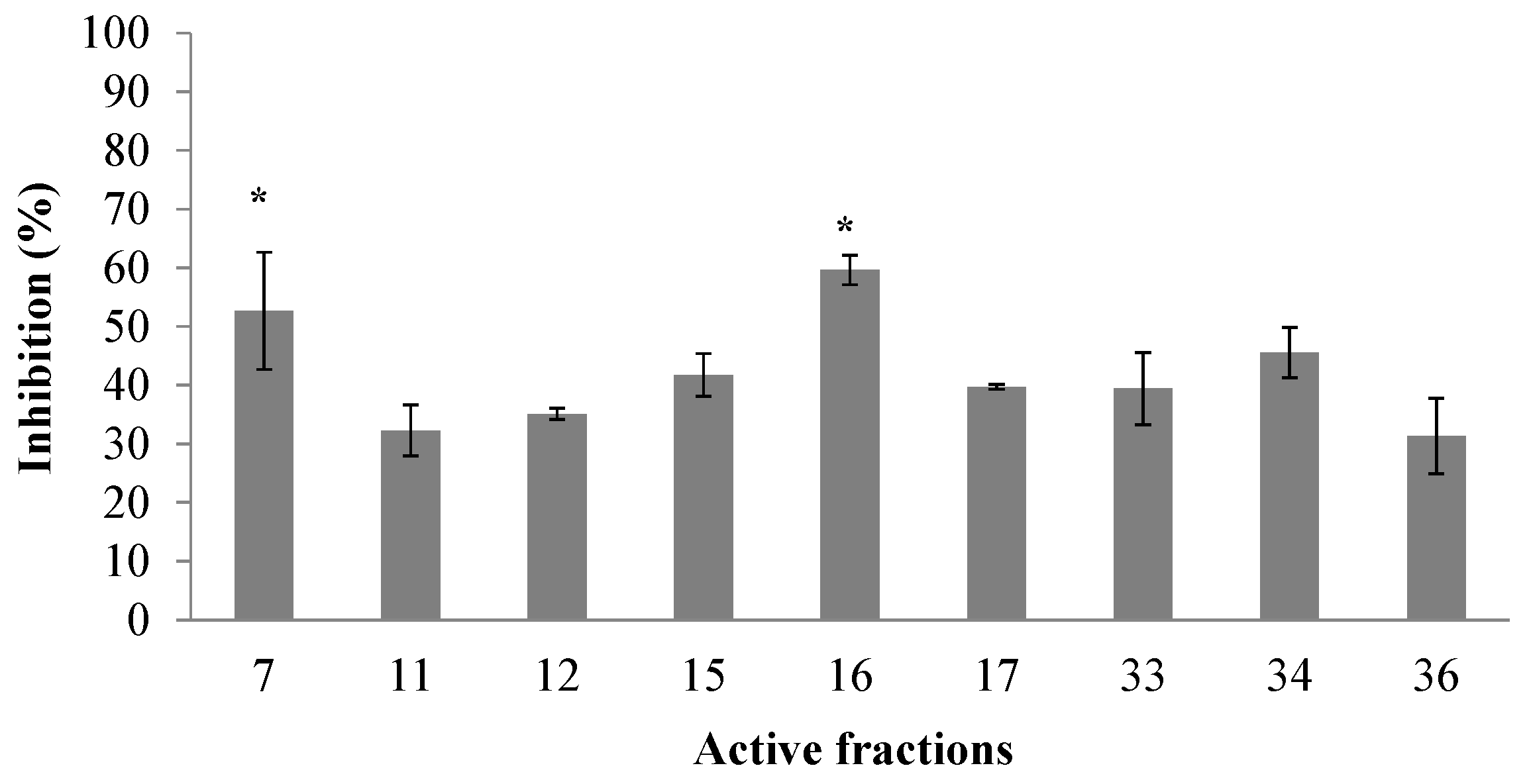
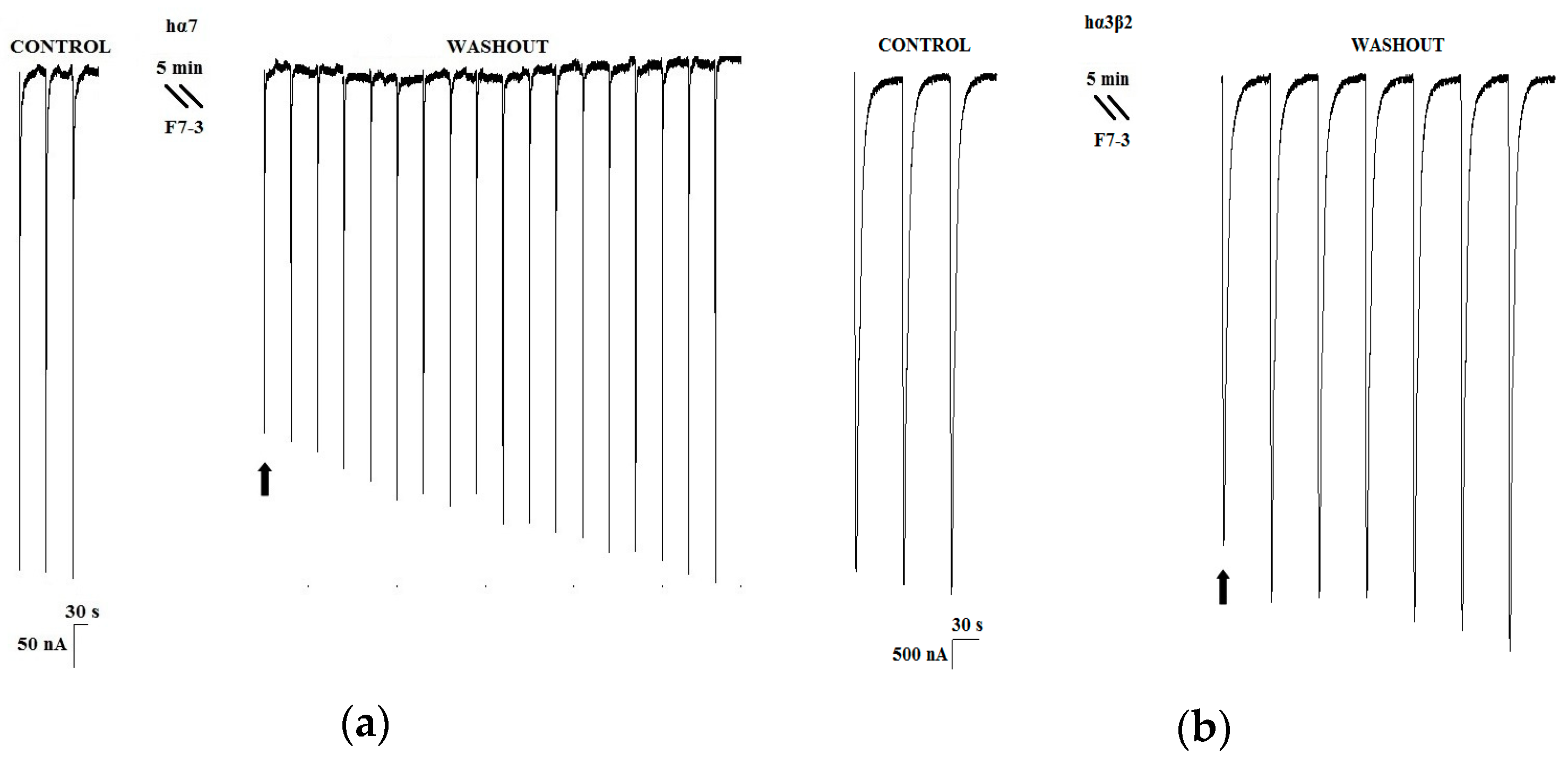
2.1. Peptidic Fractions of the C. princeps Venom and Determination of Activity on nAChRs
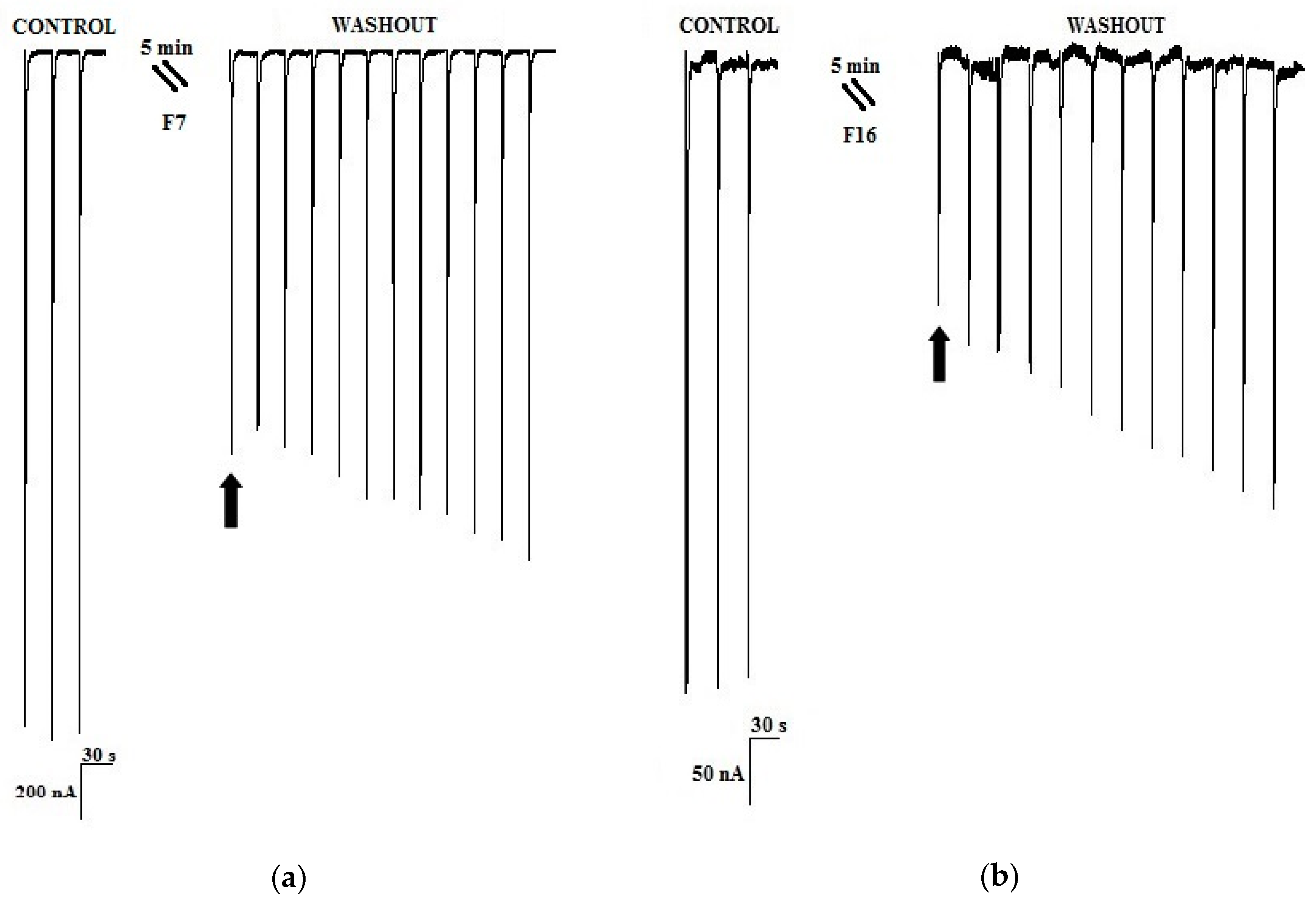
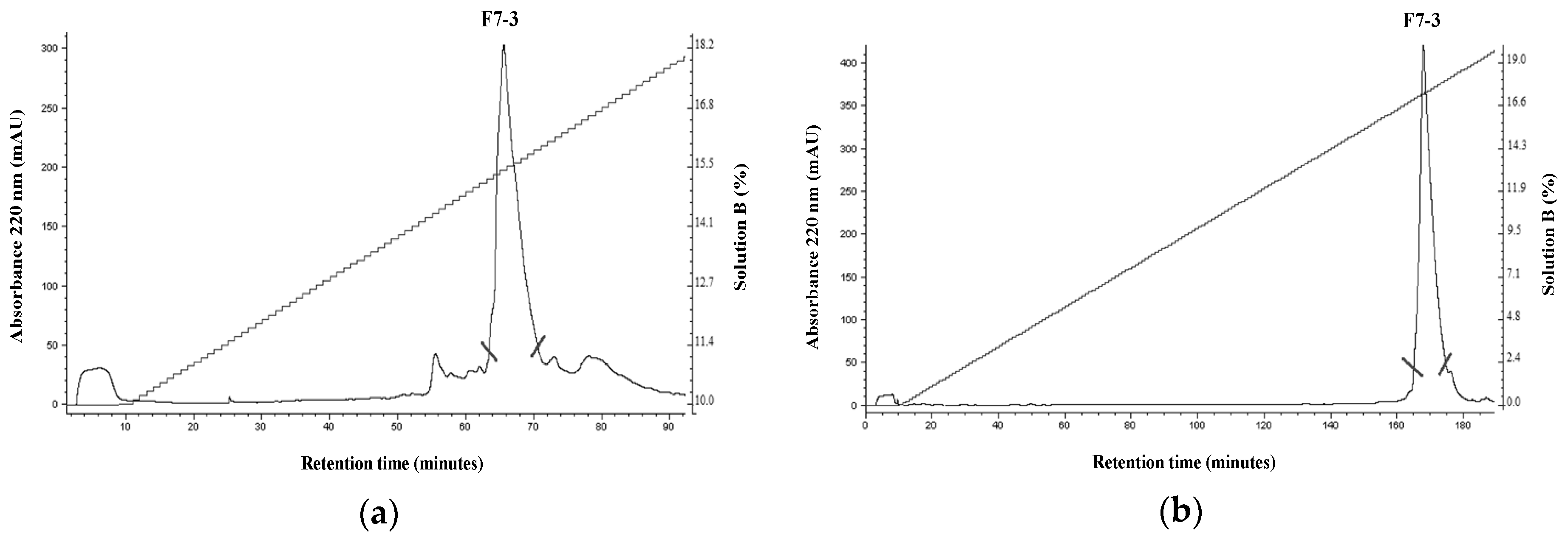
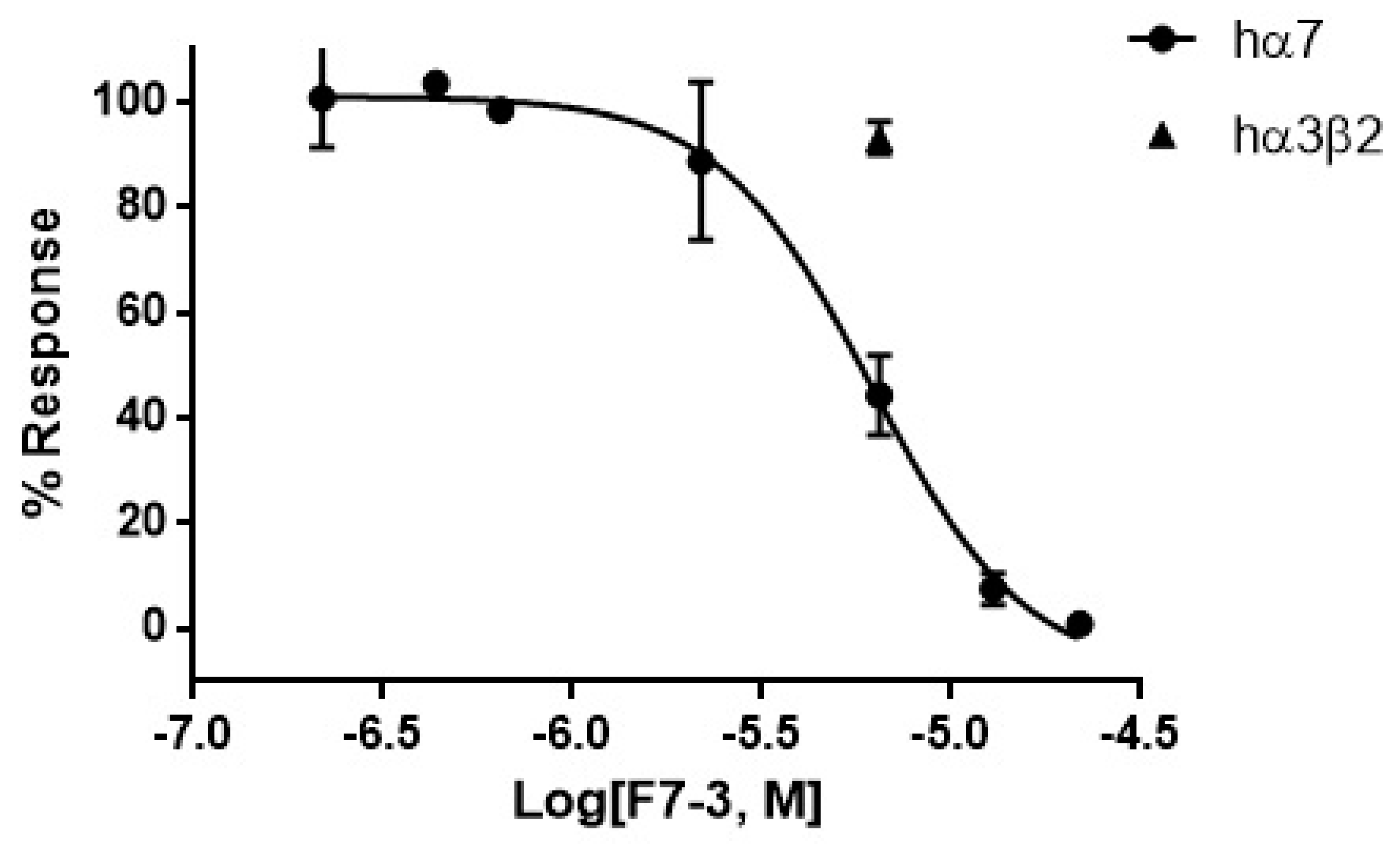
2.2. Purification of an Active Peptide of C. princeps Venom and Determination of Activity on nAChRs
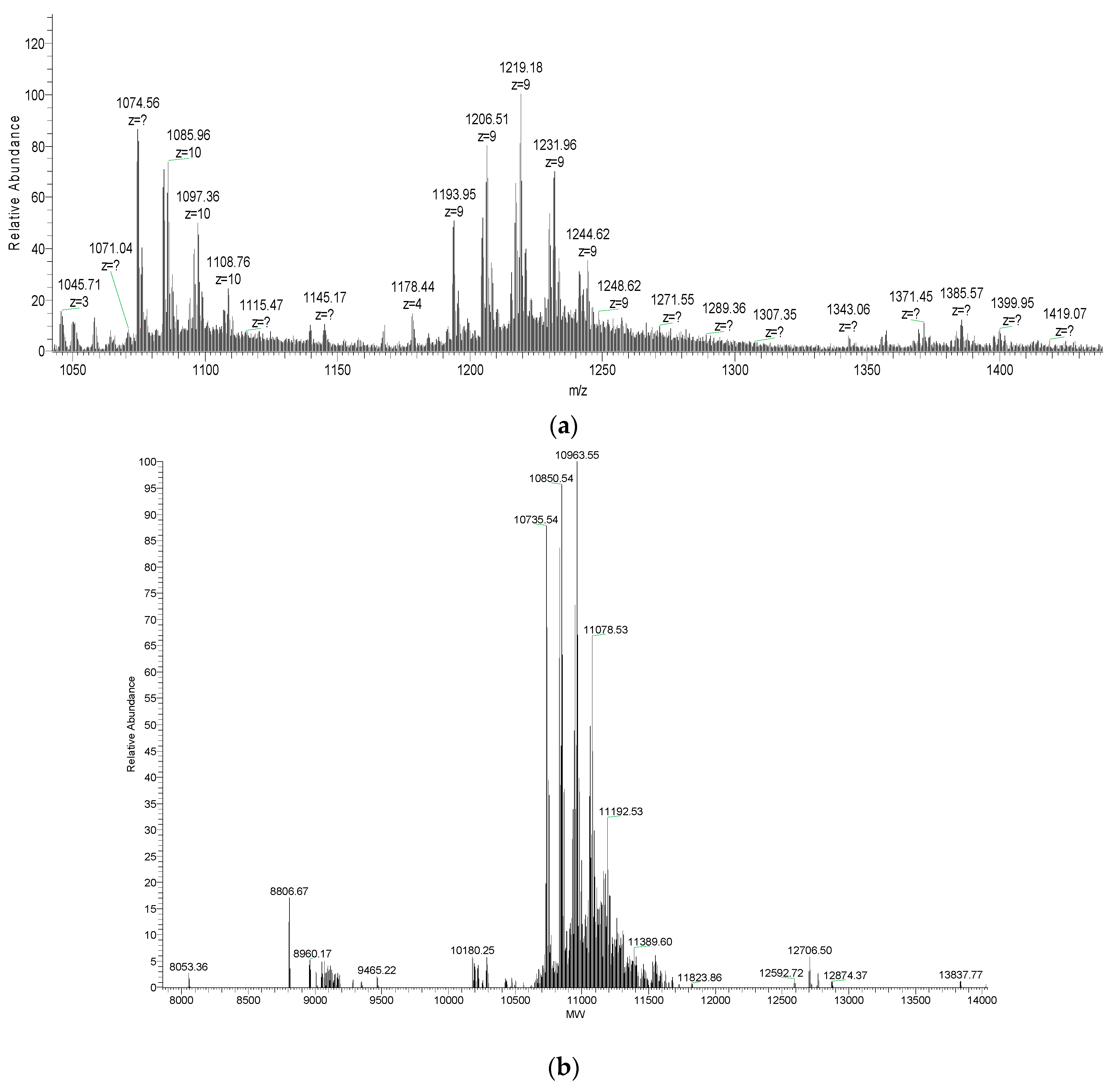
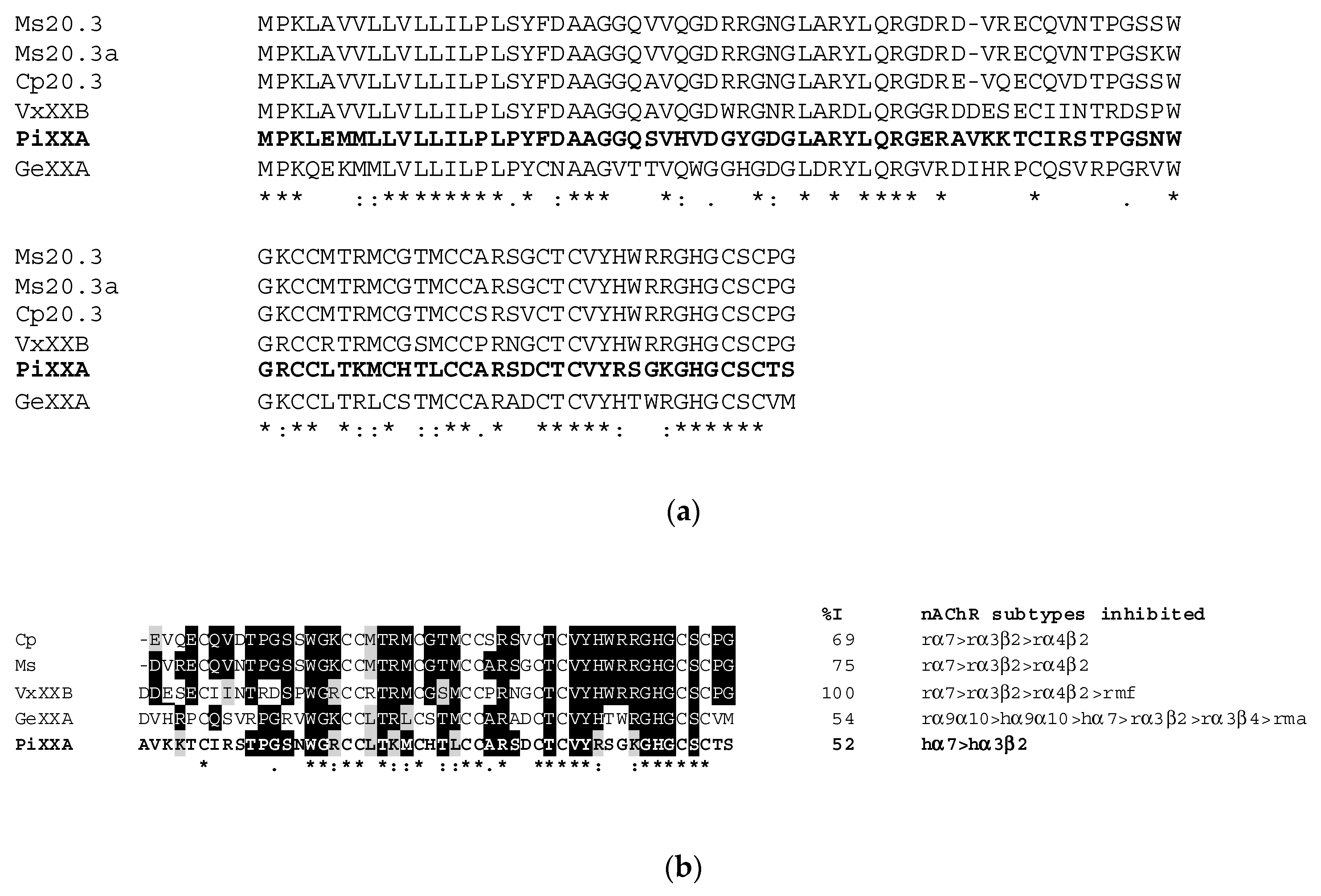
2.3. Molecular Mass and Amino Acid Sequence of the Purified Peptide
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Obtainment of Crude Venom Extract
4.3. Fractionation of the Crude Venom Extract
4.4. Purification of the Active Peptide
4.5. Determination of Electrophysiological Activity on nAChRs
4.6. Determination of Molecular Mass and Amino Acid Sequence of the Purified Peptide
4.7. Similarity Search and Sequence Alignment
4.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hopping, G.; Wang, C.I.A.; Hogg, R.C.; Nevin, S.T.; Lewis, R.J.; Adams, D.J.; Alewood, P.F. Hydrophobic residues at position 10 of α-conotoxin PnIA influence subtype selectivity between α7 and α3β2 neuronal nicotinic acetylcholine receptors. Biochem. Pharmacol. 2014, 91, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Zhang, T.; Kompella, S.N.; Yan, M.; Lu, A.; Wang, Y.; Shao, X.; Chi, C.; Adams, D.J.; Ding, J.; et al. Conotoxin αD-GeXXA utilizes a novel strategy to antagonize nicotinic acetylcholine receptors. Sci. Rep. 2015, 5, 14261. [Google Scholar] [CrossRef] [PubMed]
- Giribaldi, J.; Dutertre, S. α-Conotoxins to explore the molecular, physiological and pathophysiological functions of neuronal acetylcholine receptors. Neurosci. Lett. 2017, 679, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Abraham, N.; Lewis, R.J. Review—Neuronal nicotinic acetylcholine receptor modulators from cone snails. Mar. Drugs 2018, 16, 208. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhao, C.; Liu, Z.; Wang, X.; Liu, N.; Du, W.; Dai, Q. Structural and functional characterization of a novel α-conotoxin Mr1.7 from Conus marmoreus targeting neuronal nAChR α3β2, α9α10 and α6/α3β2β3 subtypes. Mar. Drugs 2015, 13, 3259–3275. [Google Scholar] [CrossRef] [PubMed]
- Prashanth, J.R.S.; Dutertre, S.; Lewis, R.J. Pharmacology of predatory and defensive venom peptides in cone snails. Mol. Biosyst. 2017, 13, 2453–2465. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, S.; Jin, A.H.; Vetter, I.; Hamilton, B.; Sunagar, K.; Lavergne, V.; Dutertre, V.; Fry, B.G.; Antunes, A.; Venter, D.J.; et al. Evolution of separate predation- and defence-evoked venoms in carnivorous cone snails. Nat. Commun. 2014, 5, 3521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, P.S.; Kumar, D.S.; Umamaheswari, S. A perspective on toxicology of Conus venom peptides. Asian Pac. J. Trop. Med. 2015, 8, 337–351. [Google Scholar] [CrossRef]
- Akondi, K.B.; Muttenthaler, M.; Dutertre, S.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Discovery, synthesis, and structure-activity relationships of conotoxins. Chem. Rev. 2014, 114, 5815–5847. [Google Scholar] [CrossRef]
- Lebbe, E.K.; Peigneur, S.; Wijesekara, I.; Tytgat, J. Conotoxins targeting nicotinic acetylcholine receptors: An overview. Mar. Drugs 2014, 12, 2970–3004. [Google Scholar] [CrossRef]
- Olivera, B.M.; Quik, M.; Vincler, M.; McIntosh, J.M. Subtype-selective conopeptides targeted to nicotinic receptors: Concerted discovery and biomedical applications. Channels 2008, 2, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, S.; Nicke, A.; Tsetlin, V.I. Nicotinic acetylcholine receptor inhibitors derived from snake and snail venoms. Neuropharmacology 2017, 127, 196–223. [Google Scholar] [CrossRef] [PubMed]
- Loughnan, M.; Nicke, A.; Jones, A.; Schroeder, C.I.; Nevin, S.T.; Adams, D.J.; Alewood, P.F.; Lewis, R.J. Identification of a novel class of nicotinic receptor antagonists: Dimeric conotoxins VxXIIA, VxXIIB, and VxXIIC from Conus vexillum. J. Biol. Chem. 2006, 281, 24745–24755. [Google Scholar] [CrossRef] [PubMed]
- Morales-González, D.; Flores-Martínez, E.; Zamora-Bustillos, R.; Rivera-Reyes, R.; Michel-Morfín, J.E.; Landa-Jaime, V.; Falcón, A.; Aguilar, M.B. Diversity of A-conotoxins of three worm-hunting cone snails (Conus brunneus, Conus nux, and Conus princeps) from the Mexican Pacific coast. Peptides 2015, 68, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Mass Spectrometry and Biotechnology Resource of IonSourcesm Mass Spectrometry Educational Resource Website. Available online: http://www.ionsource.com/programs/PEPCALC.HTM (accessed on 21 May 2019).
- Loughnan, M.L.; Nicke, A.; Lawrence, N.; Lewis, R.J. Novel αD-conopeptides and their precursors identified by cDNA cloning define the D-conotoxin superfamily. Biochemistry 2009, 48, 3717–3729. [Google Scholar] [CrossRef] [PubMed]
- Kauferstein, S.; Kendel, Y.; Nicke, A.; Coronas, F.I.V.; Possani, L.D.; Favreau, P.; Krizaj, I.; Wunder, C.; Kauert, G.; Mebs, D. New conopeptides of the D-superfamily selectively inhibiting neuronal nicotinic acetylcholine receptors. Toxicon 2009, 54, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Jeziorski, M.C.; Michel-Morfín, J.E.; Landa-Jaime, V.; Falcón, A.; Varela-Echavarría, A.; Ayala-Sumuano, J.T.; Aguilar, M.B. Transcriptomic analysis of the venom gland of the cone snail Conus princeps from the Mexican Pacific. In Proceedings of the 12th Congress of the Pan-American Section of the International Society on Toxinology, Miami Beach, FL, USA, 18–23 September 2016. [Google Scholar]
- Bernáldez, J.; Jiménez, S.; González, L.J.; Noda, F.J.; Soto, E.; Salceda, E.; Chávez, D.; Aguilar, M.B.; Licea-Navarro, A. A new member of gamma-conotoxin family isolated from Conus princeps displays a novel molecular target. Toxins 2016, 8, 39. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Sámano, A.C.; Falcón, A.; Batista, C.V.F.; Zamudio, F.; Michel-Morfín, J.E.; Landa-Jaime, V.; López-Vera, E.; Martínez-Romero, G.M.; Aguilar, M.B. γ-Conotoxin PiVIIA from Conus princeps venom potentiates hα3β2 nicotinic acetylcholine receptors. In Proceedings of the Invertebrate Neuropeptide Conference 2019, Campeche, Mexico, 17–21 February 2019. [Google Scholar]
- Zamora-Bustillos, R.; Rivera-Reyes, R.; Aguilar, M.B.; Michel-Morfín, E.; Landa-Jaime, V.; Falcón, A.; Heimer, E.P. Identification, by RT-PCR, of eight novel I2-conotoxins from the worm-hunting cone snails Conus brunneus, Conus nux, and Conus princeps from the eastern Pacific (Mexico). Peptides 2014, 53, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, D.; Lee, C.H.; Flood, D.; Marger, F.; Donnelly-Roberts, D. Therapeutic potential of α7 nicotinic acetylcholine receptors. Pharmacol. Rev. 2015, 67, 1025–1073. [Google Scholar] [CrossRef] [PubMed]
- Armishaw, C.; Jensen, A.A.; Balle, T.; Clark, R.J.; Harpsøe, K.; Skonberg, C.; Liljefors, T.; Strømgaard, K. Rational design of α-conotoxin analogues targeting α7 nicotinic acetylcholine receptors. J. Biol. Chem. 2009, 284, 9498–9512. [Google Scholar] [CrossRef] [PubMed]
- Daly, N.L.; Callaghan, B.; Clark, R.J.; Nevin, S.T.; Adams, D.J.; Craik, D.J. Structure and activity of α-conotoxin PeIA at nicotinic acetylcholine receptor subtypes and GABAB receptor-coupled N-type calcium channels. J. Biol. Chem. 2011, 286, 10233–10237. [Google Scholar] [CrossRef] [PubMed]
- Conoserver Website. Available online: http://www.conoserver.org/ (accessed on 12 May 2019).
- Ellison, M.; Gao, F.; Wang, H.L.; Sine, S.M.; Olivera, B.M. α-conotoxins ImI and ImII target distinct regions of the human alpha7 nicotinic acetylcholine receptor and distinguish human nicotinic receptor subtypes. Biochemistry 2004, 43, 16019–16026. [Google Scholar] [CrossRef] [PubMed]
- Franco, A.; Pisarewicz, K.; Moller, C.; Mora, D.; Fields, G.B.; Mari, F. Hyperhydroxylation: A new strategy for neuronal targeting by venomous marine molluscs. Prog. Mol. Subcell. Biol. 2006, 43, 83–103. [Google Scholar] [PubMed]
- Inserra, M.C.; Kompella, S.N.; Vetter, I.; Brust, A.; Daly, N.L.; Cuny, H.; Craik, D.J.; Alewood, P.F.; Adams, D.J.; Lewis, R.J. Isolation and characterization of α-conotoxin LsIA with potent activity at nicotinic acetylcholine receptors. Biochem. Pharmacol. 2013, 86, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhu, X.; Zhang, L.; Kudryavtsev, D.; Kasheverov, I.; Lei, Y.; Zhangsun, D.; Tsetlin, V.; Luo, S. Species specificity of rat and human α7 nicotinic acetylcholine receptors towards different classes of peptide and protein antagonists. Neuropharmacology 2018, 139, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Azam, L.; McIntosh, J.M. Molecular basis for the differential sensitivity of rat and human α9α10 nAChRs to α-conotoxin RgIA. J. Neurochem. 2012, 122, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Kompella, S.N.; Adams, D.J.; Craik, D.J.; Kaas, Q. Determination of the α-conotoxin Vc1.1 binding site on the α9α10 nicotinic acetylcholine receptor. J. Med. Chem. 2013, 56, 3557–3567. [Google Scholar] [CrossRef] [PubMed]
- Kompella, S.N.; Cuny, H.; Hung, A.; Adams, D.J. Molecular basis for differential sensitivity of α-conotoxin RegIIA at rat and human neuronal nicotinic acetylcholine receptors. Mol. Pharmacol. 2015, 88, 993–1001. [Google Scholar] [CrossRef]
- Puillandre, N.; Bouchet, P.; Duda, T.F., Jr.; Kauferstein, S.; Kohn, A.J.; Olivera, B.M.; Watkins, M.; Meyer, C. Molecular phylogeny and evolution of the cone snails (Gastropoda, Conoidea). Mol. Phylogenet. Evol. 2014, 78, 290–303. [Google Scholar] [CrossRef] [Green Version]
- Mebs, D.; Kordis, D.; Kendel, Y.; Kauferstein, S. The evolution of αD-conopeptides targeting neuronal nicotinic acetylcholine receptors. Acta Chim. Slov. 2011, 58, 730–734. [Google Scholar]
- Prashanth, J.R.; Dutertre, S.; Jin, A.H.; Lavergne, V.; Hamilton, B.; Cardoso, F.C.; Griffin, J.; Venter, D.J.; Alewood, P.F.; Lewis, R.J. The role of defensive ecological interactions in the evolution of conotoxins. Mol. Ecol. 2016, 25, 598–615. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Cartier, G.E.; Yoshikami, D.; Gray, W.R.; Luo, S.; Olivera, B.M.; McIntosh, J.M. A new α-conotoxin which targets α3β2 nicotinic acetylcholine receptors. J. Biol. Chem. 1996, 271, 7522–7528. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Wootton, J.C.; Gertz, E.M.; Agarwala, R.; Morgulis, A.; Schäffer, A.A.; Yu, Y.K. Protein database searches using compositionally adjusted substitution matrices. FEBS J. 2005, 272, 5101–5109. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GenomeNet of the Kyoto University Bioinformatics Center. Available online: https://www.genome.jp/tools-bin/clustalw (accessed on 12 May 2019).









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-Sámano, A.C.; Falcón, A.; Zamudio, F.; Batista, C.V.F.; Michel-Morfín, J.E.; Landa-Jaime, V.; López-Vera, E.; Jeziorski, M.C.; Aguilar, M.B. αD-Conotoxins in Species of the Eastern Pacific: The Case of Conus princeps from Mexico. Toxins 2019, 11, 405. https://doi.org/10.3390/toxins11070405
Hernández-Sámano AC, Falcón A, Zamudio F, Batista CVF, Michel-Morfín JE, Landa-Jaime V, López-Vera E, Jeziorski MC, Aguilar MB. αD-Conotoxins in Species of the Eastern Pacific: The Case of Conus princeps from Mexico. Toxins. 2019; 11(7):405. https://doi.org/10.3390/toxins11070405
Chicago/Turabian StyleHernández-Sámano, Arisaí C., Andrés Falcón, Fernando Zamudio, César V.F. Batista, Jesús Emilio Michel-Morfín, Víctor Landa-Jaime, Estuardo López-Vera, Michael C. Jeziorski, and Manuel B. Aguilar. 2019. "αD-Conotoxins in Species of the Eastern Pacific: The Case of Conus princeps from Mexico" Toxins 11, no. 7: 405. https://doi.org/10.3390/toxins11070405
APA StyleHernández-Sámano, A. C., Falcón, A., Zamudio, F., Batista, C. V. F., Michel-Morfín, J. E., Landa-Jaime, V., López-Vera, E., Jeziorski, M. C., & Aguilar, M. B. (2019). αD-Conotoxins in Species of the Eastern Pacific: The Case of Conus princeps from Mexico. Toxins, 11(7), 405. https://doi.org/10.3390/toxins11070405