Distribution of Toxinogenic Methicillin-Resistant and Methicillin-Susceptible Staphylococcus aureus from Different Ecological Niches in Algeria

 , and
, and 
Abstract
:1. Introduction
2. Results
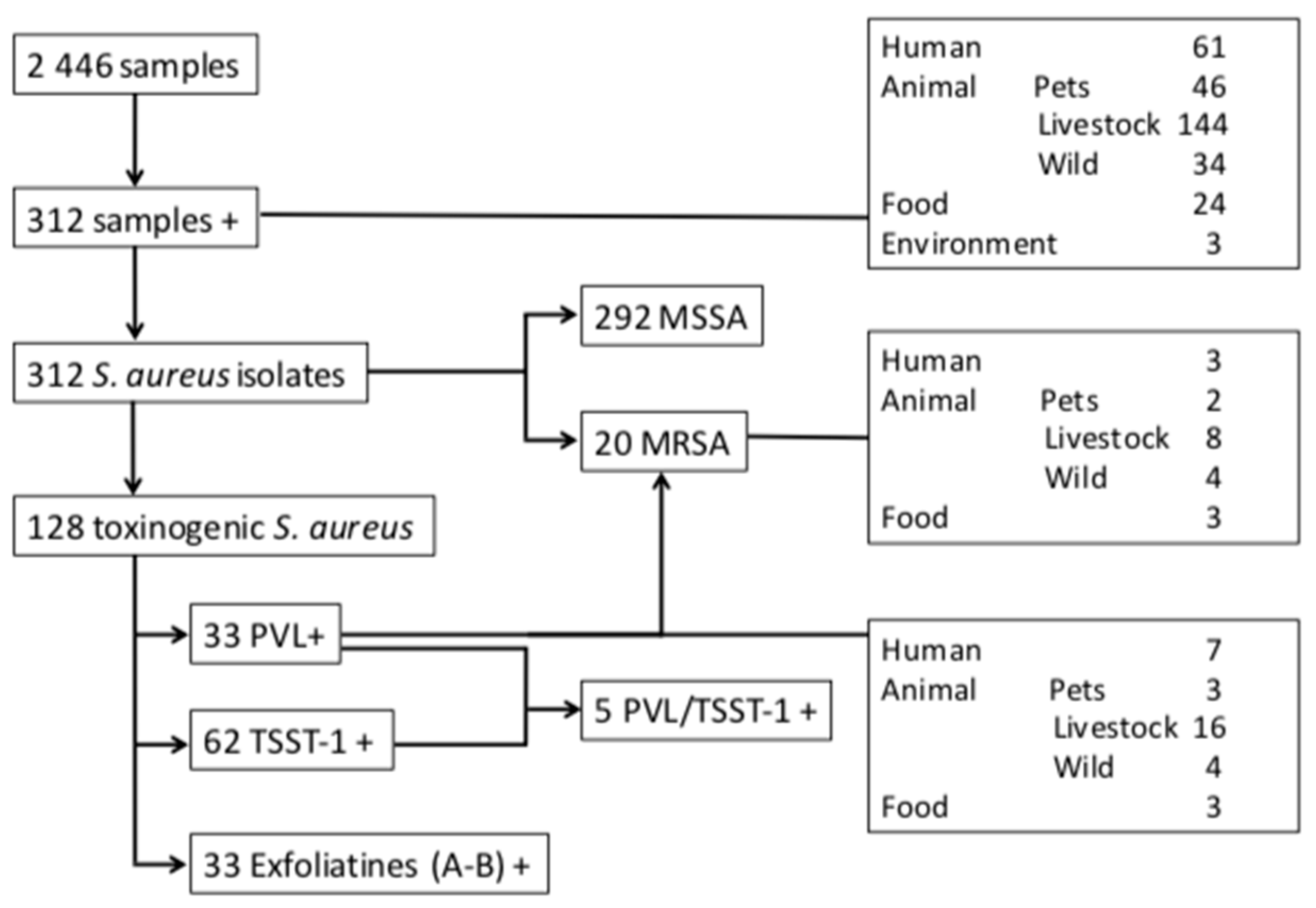
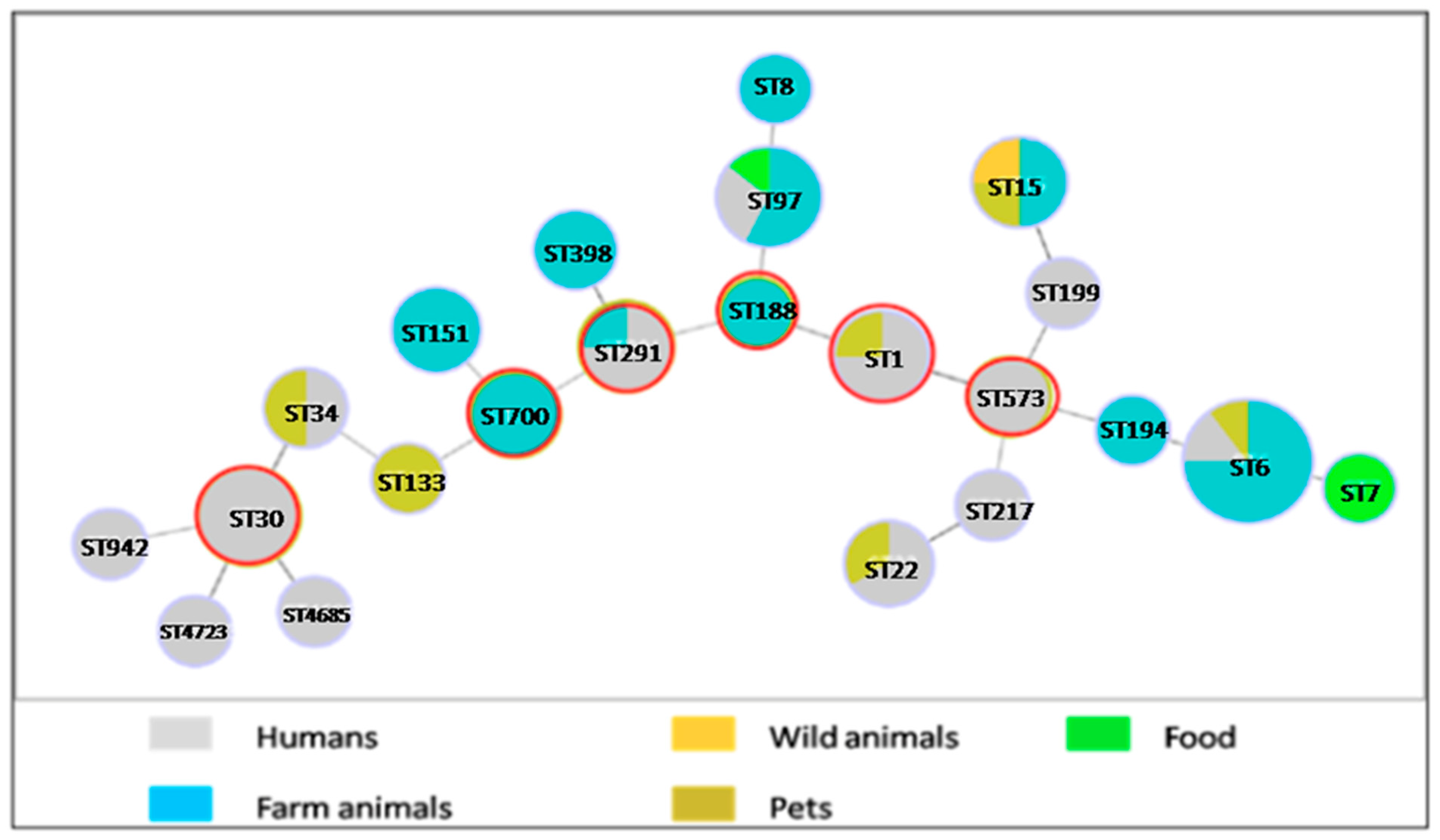
2.1. Population of the Isolates
2.2. Antimicrobial Susceptibility Profiles
2.3. Importance of Toxinogenic S. Aureus
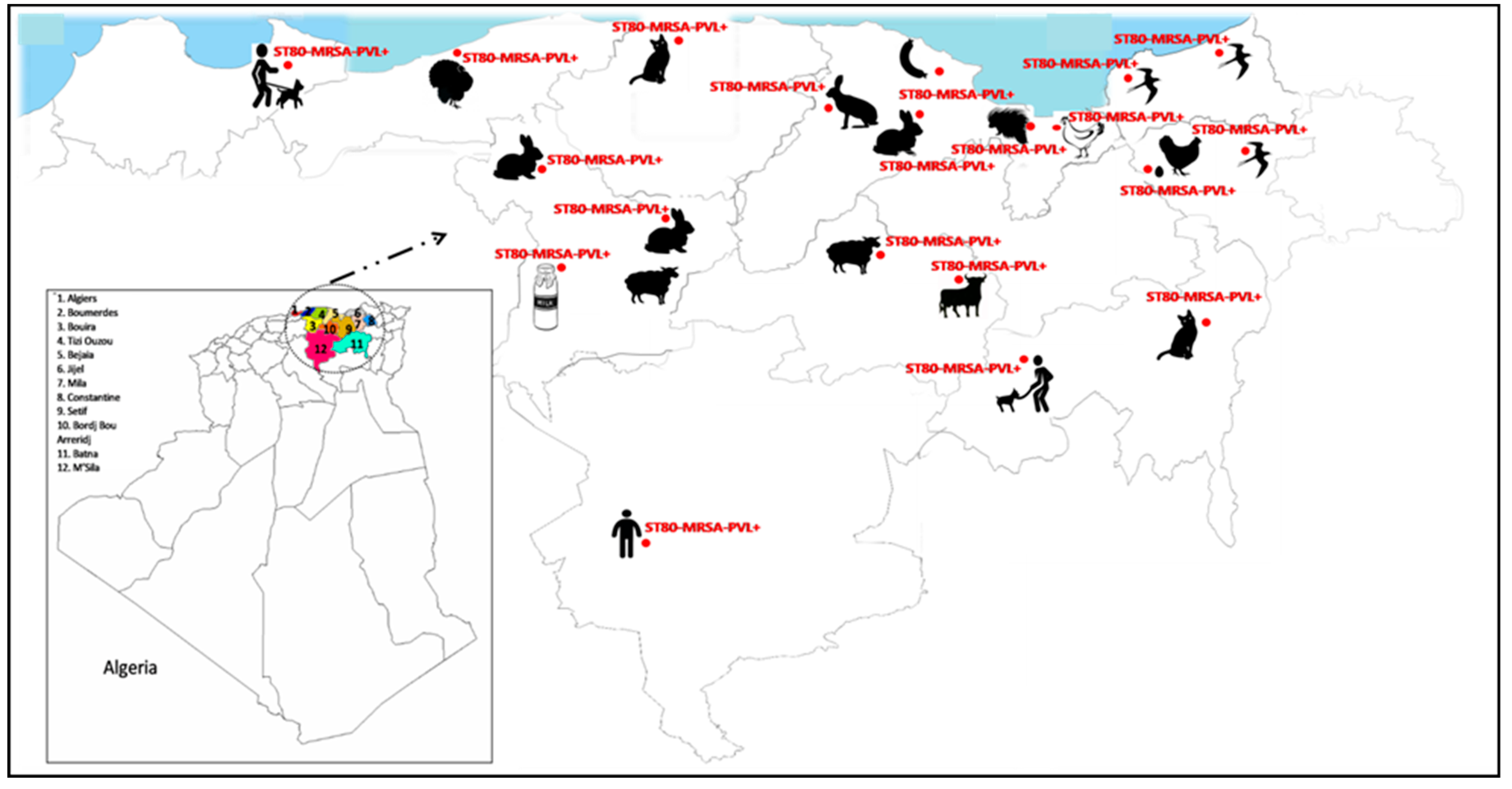
2.4. Characteristics of Panton–Valentine Leukocidin (PVL) + S. aureus
2.5. Characteristics of the Other Toxinogenic S. aureus
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Sampling and Microbiological Procedures
5.2. Antibiotic Susceptibility Testing
5.3. DNA Arrays Procedures
5.4. MultiLocus Sequence Typing (MLST)
5.5. goeBURST Analysis of Methicillin-Susceptible S. Aureus (MSSA) Distribution
5.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G., Jr. Methicillin-resistant Staphylococcus aureus: An overview of basic and clinical research. Lancet Infect. Dis. 2019, 17, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Lakhundi, S.; Zhang, K. Methicillin-resistant Staphylococcus aureus: Molecular characterization, evolution, and epidemiology. Clin. Microbiol. Rev. 2018, 31, e00020-18. [Google Scholar] [CrossRef] [PubMed]
- Newsom, S.W. Ogston’s coccus. J. Hosp. Infect. 2008, 70, 369–372. [Google Scholar] [CrossRef] [PubMed]
- Grundmann, H.; Aires-de-Sousa, M.; Boyce, J.; Tiemersma, E. Emergence and resurgence of methicillin-resistant Staphylococcus aureus as a public-health threat. Lancet 2006, 368, 874–885. [Google Scholar] [CrossRef]
- Ayeni, F.A.; Ruppitsch, W.; Allerberger, F. Molecular characterization of clonal lineage and staphylococcal toxin genes from S. aureus in Southern Nigeria. Peer J. 2018, 6, e5204. [Google Scholar] [CrossRef] [PubMed]
- Planet, P.J.; Narechania, A.; Chen, L.; Mathema, B.; Boundy, S.; Archer, G.; Kreiswirth, B. Architecture of a species phylogenomics of Staphylococcus aureus. Trends Microbiol. 2017, 25, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Uhlemann, A.C.; Dordel, J.; Knox, J.R.; Raven, K.E.; Parkhill, J.; Holden, M.T.; Peacock, S.J.; Lowy, F.D. Molecular tracing of the emergence, diversification, and transmission of S. aureus sequence type 8 in a New York community. Proc. Natl. Acad. Sci. USA 2014, 111, 6738–6743. [Google Scholar] [CrossRef] [PubMed]
- Saeed, K.; Gould, I.; Esposito, S.; Ahmad-Saeed, N.; Ahmed, S.S.; Alp, E.; Bal, A.M.; Bassetti, M.; Bonnet, E.; Chan, M.; et al. Panton-Valentine leukocidin-positive Staphylococcus aureus: A position statement from the International Society of Chemotherapy. Int. J. Antimicrob. Agents 2018, 51, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Shallcross, L.J.; Fragaszy, E.; Johnson, A.M.; Hayward, A.C. The role of the Panton-Valentine leucocidin toxin in staphylococcal disease: A systematic review and meta-analysis. Lancet Infect. Dis. 2013, 13, 43–54. [Google Scholar] [CrossRef]
- Alioua, M.A.; Labid, A.; Amoura, K.; Bertine, M.; Gacemi-Kirane, D.; Dekhil, M. Emergence of the European ST80 clone of community-associated methicillin-resistant Staphylococcus aureus as a cause of healthcare-associated infections in Eastern Algeria. Med. Mal. Infect. 2014, 44, 180–183. [Google Scholar] [CrossRef]
- Djoudi, F.; Bonura, C.; Benallaoua, S.; Touati, A.; Touati, D.; Aleo, A.; Cala, C.; Fasciana, T.; Mammina, C. Panton-Valentine leukocidin positive sequence type 80 methicillin-resistant Staphylococcus aureus carrying a staphylococcal cassette chromosome mec type IVc is dominant in neonates and children in an Algiers hospital. New Microbiol. 2013, 36, 49–55. [Google Scholar] [PubMed]
- Irek, E.O.; Amupitan, A.A.; Obadare, T.O.; Aboderin, A.O. A systemic review of healthcare-associated infections in Africa: An antimicrobial resistance perspective. Afr. J. Lab. Med. 2018, 7, 796. [Google Scholar] [CrossRef] [PubMed]
- Dweba, C.C.; Zishiri, O.T.; El Zowalaty, M.E. Methicillin-resistant Staphylococcus aureus: Livestock-associated, antimicrobial, and heavy metal resistance. Infect. Drug Resist. 2018, 11, 2497–2509. [Google Scholar] [CrossRef] [PubMed]
- Bekkhoucha, S.N.; Cady, A.; Gautier, P.; Itim, F.; Donnio, P.Y. A portrait of Staphylococcus aureus from the other side of the Mediterranean Sea: Molecular characteristics of isolates from Western Algeria. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 553–555. [Google Scholar] [CrossRef] [PubMed]
- Antri, K.; Rouzic, N.; Dauwalder, O.; Boubekri, I.; Bes, M.; Lina, G.; Vandenesch, F.; Tazir, M.; Ramdani-Bouguessa, N.; Etienne, J. High prevalence of methicillin-resistant Staphylococcus aureus clone ST80-IV in hospital and community settings in Algiers. Clin. Microbiol. Infect. 2011, 17, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Basset, P.; Amhis, W.; Blanc, D.S. Changing molecular epidemiology of methicillin-resistant Staphylococcus aureus in an Algerian hospital. J. Infect. Dev. Ctries. 2015, 9, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Djoudi, F.; Benallaoua, S.; Aleo, A.; Touati, A.; Challal, M.; Bonura, C.; Mammina, C. Descriptive epidemiology of nasal carriage of Staphylococcus aureus and methicillin-resistant Staphylococcus aureus among patients admitted to two healthcare facilities in Algeria. Microb. Drug Resist. 2015, 21, 218–223. [Google Scholar] [CrossRef]
- Agabou, A.; Ouchenane, Z.; Ngba Essebe, C.; Khemissi, S.; Chehboub, M.T.E.; Chehboub, I.B.; Sotto, A.; Dunyach-Remy, C.; Lavigne, J.P. Emergence of nasal carriage of ST80 and ST152 PVL + Staphylococcus aureus isolates from livestock in Algeria. Toxins (Basel) 2017, 9, 303. [Google Scholar] [CrossRef]
- One Health. Available online: https://www.cdc.gov/onehealth/index.html (accessed on 31 July 2019).
- Coll, F.; Harrison, E.M.; Toleman, M.S.; Reuter, S.; Raven, K.E.; Blane, B.; Palmer, B.; Kappeler, A.R.M.; Brown, N.M.; Török, M.E.; et al. Longitudinal genomic surveillance of MRSA in the UK reveals transmission patterns in hospitals and the community. Sci. Transl. Med. 2017, 9, eaak9745. [Google Scholar] [CrossRef]
- Rolo, J.; Miragaia, M.; Turlej-Rogacka, A.; Empel, J.; Bouchami, O.; Faria, N.A.; Tavares, A.; Hryniewicz, W.; Fluit, A.C.; de Lencastre, H.; et al. High genetic diversity among community-associated Staphylococcus aureus in Europe: Results from a multicenter study. PLoS ONE 2012, 7, e34768. [Google Scholar] [CrossRef]
- Gharsa, H.; Ben Slama, K.; Gómez-Sanz, E.; Lozano, C.; Zarazaga, M.; Messadi, L.; Boudabous, A.; Torres, C. Molecular characterization of Staphylococcus aureus from nasal samples of healthy farm animals and pets in Tunisia. Vector Borne Zoonotic Dis. 2015, 15, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Orchein, R.C.; Hundstad, D.A.; Fritz, S.A.; Loughman, J.A.; Mitchell, K.; Storch, E.K.; Gaudreault, M.; Sellenriek, P.L.; Armstrong, J.R.; Mardis, E.R.; et al. Contribution of gentetically restricted, mthcillin-susceptible strains to the ongoing epidemic of community-acquired Staphylococcus aureus infections. Clin. Infect. Dis. 2009, 49, 536–542. [Google Scholar]
- Oosthuysen, W.F.; Orth, H.; Lombard, C.J.; Sinha, B.; Wasserman, E. Population structure analyses of Staphylococcus aureus at Tygerberg hospital, South Africa, reveals a diverse population, a high prevalence of Panton-Valentine leukocidin genes, and unitque local methicillin-resistant S. aureus clone. Clin. Microbiol. Infect. 2014, 20, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Wu, S.; Luo, W.; Su, Y.; Luan, Y.; Wang, X. Staphylococcus aureus ST6-t701 isolates from food-posoning outbreaks (2006–2013) in Xi’an, China. Foodborne Pathog. Dis. 2015, 12, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Santosaningsih, D.; Santoso, S.; Setijowati, N.; Rasyid, H.A.; Budayanti, N.S.; Suata, K.; Widhyatmoko, D.B.; Purwono, P.B.; Kuntaman, K.; Damayanti, D.; et al. Prevalence and characterisation of Staphylococcus aureus causing community-acquired skin and soft tissue infections on Java and Bali, Indonesia. Trop. Med. Int. Health 2018, 23, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Pirolo, M.; Gioffrè, A.; Visaggio, D.; Gherardi, M.; Pavia, G.; Samele, P.; Ciambrone, L.; Di Natale, R.; Spatari, G.; Casalinuovo, F.; et al. Prevalence, molecular epidemiology, and antimicrobial resistance of methicillin-resistant Staphylococcus aureus from swine in southern Italy. BMC Microbiol. 2019, 19, 51. [Google Scholar] [CrossRef]
- Cormican, P.; Keane, O.M. Complete genome sequences of Sequence Type 71 (ST71) and ST97 Staphylococcus aureus isolates from bovine milk. Microbiol. Resour. Announc. 2018, 7, e00954-18. [Google Scholar] [CrossRef]
- Panahi, M.; Saei, H.D. Genetic diversity and methicillin resistance of Staphylococcus aureus originating from buffaloes with mastitis in Iran. Comp. Immunol. Microbiol. Infect. Dis. 2019, 62, 19–24. [Google Scholar] [CrossRef]
- Donkor, E.S.; Jamrozy, D.; Mills, R.O.; Dankwah, T.; Amoo, P.K.; Egyir, B.; Badoe, E.V.; Twasam, J.; Bentley, S.D. A genomic infection control study for Staphylococcus aureus in two Ghanaian hospitals. Infect. Drug Resist. 2018, 11, 1757–1765. [Google Scholar] [CrossRef]
- Grundmann, H.; Aanensen, D.M.; van den Wijngaard, C.C.; Spratt, B.G.; Harmsen, D.; Friedrich, A.W. Geographic distribution of Staphylococcus aureus causing invasive infections in Europe: A molecular-epidemiological analysis. PLoS Med. 2010, 7, e1000215. [Google Scholar] [CrossRef]
- Lozano, C.; Marí, A.; Aspiroz, C.; Gómez-Sanz, E.; Ceballos, S.; Fortuño, B.; Barcenilla, F.; Jover-Sáenz, A.; Torres, C. Nasal carriage of coagulase positive staphylococci in patients of a primary-healthcare-center: Genetic lineages and resistance and virulence genes. Enferm. Infecc. Microbiol. Clin. 2015, 33, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Suhaili, Z.; Lean, S.S.; Mohamad, N.M.; Rachman, A.R.; Desa, M.N.; Yeo, C.C. Draft genome sequence of Staphylococcus aureus KT/312045, an ST1-MSSA PVL positive isolated from pus sample in East Coast Malaysia. Genom. Data 2016, 9, 111–112. [Google Scholar] [CrossRef] [PubMed]
- Valentin-Domelier, A.S.; Girard, M.; Bertrand, X.; Violette, J.; François, P.; Donnio, P.Y.; Talon, D.; Quentin, R.; Schrenzel, J.; van der Mee-Marquet, N.; et al. Methicillin-susceptible ST398 Staphylococcus aureus responsible for bloodstream infections: An emerging human-adapted subclone? PLoS ONE 2011, 6, e28369. [Google Scholar] [CrossRef] [PubMed]
- Ramdani-Bouguessa, N.; Bes, M.; Meugnier, H.; Forey, F.; Reverdy, M.E.; Lina, G.; Vandenesch, F.; Tazir, M.; Etienne, J. Detection of methicillin-resistant Staphylococcus aureus strains resistant to multiple antibiotics and carrying the Panton-Valentine Leukocidin genes in an Algiers hospital. Antimicrob. Agents Chemother. 2006, 50, 1083–1085. [Google Scholar] [CrossRef] [PubMed]
- Chaalal, W.; Chaalal, N.; Bourafa, N.; Kihal, M.; Diene, S.M.; Rolain, J.M. Characterization of Staphylococcus aureus isolated from food products in Western Algeria. Foodborne Pathog. Dis. 2018, 15, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Malachowa, N.; DeLeo, F.R. Mobile genetic elements of Staphylococcus aureus. Cell. Mol. Life Sci. 2010, 67, 3057–3071. [Google Scholar] [CrossRef] [PubMed]
- Monecke, S.; Coombs, G.; Shore, A.C.; Coleman, D.C.; Akpaka, P.; Borg, M.; Chow, H.; Ip, M.; Jatzwauk, L.; Jonas, D.; et al. A field guide to pandemic, epidemic and sporadic clones of methicillin-resistant Staphylococcus aureus. PLoS ONE 2011, 6, e17936. [Google Scholar] [CrossRef]
- Bierowiec, K.; Ploneczka-Janeczko, K.; Rypula, K. Prevalence and risk factors of colonisation with Staphylococcus aureus in healthy pet cats kept in the city households. Biomed. Res. Int. 2016, 2016, 3070524. [Google Scholar] [CrossRef]
- Papadimitriou-Olivgeris, M.; Drougka, E.; Fligou, F.; Dodou, V.; Kolonitsiou, F.; Filos, K.S.; Anastassiou, E.D.; Petinaki, E.; Marangos, M.; Spiliopoulou, I. Spread of Tst-positive Staphylococcus aureus strains belonging to ST30 clone among patients and healthcare workers in two intensive care units. Toxins (Basel) 2017, 9, 270. [Google Scholar] [CrossRef]
- Sung, J.M.; Lloyd, D.H.; Lindsay, J.A. Staphylococcus aureus host specificity: Comparative genomics of human versus animal isolates by multi-strain microarray. Microbiology 2008, 154, 1949–1959. [Google Scholar] [CrossRef]
- Asanin, J.; Misic, D.; Aksentijevic, K.; Tambur, Z.; Rakonjac, B.; Kovacevic, I.; Spergser, J.; Loncaric, I. Genetic profiling and comparison of human and animal methicillin-resistant Staphylococcus aureus (MRSA) isolates from Serbia. Antibiotics (Basel) 2019, 16, 26. [Google Scholar] [CrossRef] [PubMed]
- Jarraud, S.; Mougel, C.; Thioulouse, J.; Lina, G.; Meugnier, H.; Forey, F.; Nesme, X.; Etienne, J.; Vandenesch, F. Relationships between Staphylococcus aureus genetic background, virulence factors, agr groups (alleles), and human disease. Infect. Immun. 2002, 70, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Jamrozy, D.; Coll, F.; Matther, A.E.; Harris, S.R.; Harrison, E.M.; MacGowan, A.; Karas, A.; Elston, T.; Estée Török, M.; Parkhill, J.; et al. Evolution of mobile genetic element composition in an epidemic methicillin-resistant Staphylococcus aureus: Temporal changes correlated with frequent loss and gain events. BMC Genom. 2017, 18, 684. [Google Scholar] [CrossRef] [PubMed]
- Karakonstantis, S.; Kalemaki, D. Antimicrobial overuse and misuse in the community in Greece and link to antimicrobial resistance using methicillin-resistant S. aureus as an example. J. Infect. Public Health 2019, 12, 460–464. [Google Scholar] [CrossRef] [PubMed]
- European Committee on Antimicrobial Susceptibility Testing. Available online: https://www.sfm-microbiologie.org/wp-content/uploads/2018/12/CASFMV2_SEPTEMBRE2018.pdf (accessed on 31 July 2019).
- Sahebnasagh, R.; Saderi, H.; Owlia, P. The prevalence of resistance to methicillin in Staphylococcus aureus strains isolated from patients by PCR method for detection of mecA and nuc genes. Iran. J. Public Health 2014, 43, 84–92. [Google Scholar] [PubMed]
- Monecke, S.; Jatzwauk, L.; Weber, S.; Slickers, P.; Ehricht, R. DNA microarray-based genotyping of methicillin-resistant Staphylococcus aureus strains from Eastern Saxony. Clin. Microbiol. Infect. 2008, 14, 534–545. [Google Scholar] [CrossRef] [PubMed]
- Monecke, S.; Slickers, P.; Ehricht, R. Assignment of Staphylococcus aureus isolates to clonal complexes based on microarray analysis and pattern recognition. FEMS Immunol. Med. Microbiol. 2008, 53, 237–251. [Google Scholar] [CrossRef] [PubMed]
- Enright, M.C.; Day, N.P.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar]
- Staphylococcus Aureus MLST Databases. Available online: http://www.mlst.net/ (accessed on 31 July 2019).
- Francisco, A.P.; Bugalho, M.; Ramirez, M.; Carriço, J.A. Global optimal eBURST analysis of multilocus typing data using a graphic matroid approach. BMC Bioinform. 2009, 10, 152. [Google Scholar] [CrossRef]
- Francisco, A.P.; Vaz, C.; Monteiro, P.T.; Melo-Cristino, J.; Ramirez, M.; Carriço, J.A. PHYLOViZ: Phylogenetic inference and data visualization for sequence based typing methods. BMC Bioinform. 2012, 13, 87. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Antibiotics | Origin of Isolates | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Human | Bovine | Ovine | Goats | Rabbits | Poultry | Pets | Wild Animals | Food Products | Aquatic Environment | Total Samples | |
| Total | 61 | 18 | 34 | 8 | 15 | 69 | 46 | 34 | 24 | 3 | 312 |
| Penicillin G | 51 (83.6) | 11 (61.1) | 19 (55.8) | 4 (50) | 8 (53.3) | 51 (73.9) | 28 (60.8) | 25 (73.5) | 11 (45.8) | 1 (33.3) | 209 (66.9) |
| Cefoxitin | 3 (4.9) | 1 (5.5) | 1 (2.9) | 0 (0) | 3 (20) | 3 (4.3) | 2 (4.3) | 5 (14.7) | 2 (8.3) | 0 (0) | 20 (6.4) |
| Erythromycin | 5 (8.1) | 0 (0) | 1 (2.9) | 0 (0) | 0 (0) | 6 (8.6) | 4 (8.6) | 2 (5.8) | 6 (25) | 0 (0) | 24 (7.6) |
| Ceftobiprole | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| Clindamycin | 1 (1.6) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (0.3) |
| Quinupristin/Dalfopristin | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| Kanamycin | 4 (6.5) | 3 (16.6) | 4 (11.7) | 0 (0) | 3 (20) | 4 (5.7) | 2 (4.3) | 5 (14.7) | 4 (16.6) | 1 (33.3) | 30 (9.6) |
| Tobramycin | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 2 (8.3) | 0 (0) | 2 (0.6) |
| Gentamicin | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| Minocycline | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (1.4) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (0.3) |
| Ofloxacin | 2 (3.2) | 0 (0) | 0 (0) | 0 (0) | 2 (13.3) | 1 (1.4) | 1 (2.1) | 3 (8.8) | 2 (8.3) | 0 (0) | 11 (3.5) |
| Fusidic acid | 1 (1.6) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (1.4) | 0 (0) | 5 (14.7) | 1 (4.1) | 0 (0) | 8 (2.5) |
| Fosfomycin | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| Rifampicin | 3 (4.9) | 1 (5.5) | 1 (2.9) | 0 (0) | 3 (20) | 3 (4.3) | 2 (4.3) | 7 (20.5) | 2 (8.3) | 0 (0) | 22 (7.0) |
| Cotrimoxazole | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| Virulence and Resistance Genes | MRSA-PVL + no. (%) | MSSA-PVL + no. (%) | p Value |
|---|---|---|---|
| Virulence Genotyping | |||
| Enterotoxins | |||
| sea | 0 (0) | 8 (61.5) | < 0.001 |
| seb | 0 (0) | 2 (15.3) | NS |
| seg | 0 (0) | 2 (15.3) | NS |
| seh | 0 (0) | 3 (23) | 0.05 |
| sei | 0 (0) | 5 (38.4) | 0.005 |
| sek | 0 (0) | 1 (7.6) | NS |
| seq | 0 (0) | 1 (7.6) | NS |
| egc clustera | 0 (0) | 0 (0) | NS |
| Other Toxins | |||
| tst | 0 (0) | 5 (38.4) | 0.005 |
| etA | 0 (0) | 0 (0) | NS |
| etB | 0 (0) | 0 (0) | NS |
| etD | 20 (100) | 5 (38.4) | 0.005 |
| edinA | 0 (0) | 1 (7.6) | NS |
| edinB | 20 (100) | 5 (38.4) | 0.005 |
| Leukocidins | |||
| lukD | 20 (100) | 12 (92.3) | NS |
| lukE | 19 (95) | 12 (92.3) | NS |
| lukX | 7 (35) | 13 (100) | < 0.001 |
| lukY | 15 (75) | 13 (100) | NS |
| Hemolysins | |||
| hla | 10 (50) | 12 (92.3) | 0.02 |
| hlb | 15 (75) | 6 (46.1) | NS |
| hld | 20 (100) | 13 (100) | NS |
| hlgA | 20 (100) | 13 (100) | NS |
| hlgv | 20 (100) | 13 (100) | NS |
| Microbial Surface Components Recognizing Adhesive Matrix Molecules (MSRAMMs) | |||
| bbp | 20 (100) | 11 (84.6) | NS |
| clfA | 20 (100) | 13 (100) | NS |
| clfB | 20 (100) | 13 (100) | NS |
| cna | 0 (0) | 5 (38.4) | 0.005 |
| ebpS | 20 (100) | 13 (100) | NS |
| fib | 20 (100) | 12 (92.3) | NS |
| fnbA | 20 (100) | 12 (92.3) | NS |
| fnbB | 20 (100) | 13 (100) | NS |
| Capsule Components | |||
| cap5 | 0 (0) | 3 (23) | 0.05 |
| cap8 | 20 (100) | 10 (76.9) | NS |
| Intracellular Adhesion Polysaccharide | |||
| icaA | 20 (100) | 13 (100) | NS |
| icaC | 20 (100) | 13 (100) | NS |
| icaD | 20 (100) | 13 (100) | NS |
| Immune Evasion Cluster and Other | |||
| sak | 20 (100) | 10 (76.9) | NS |
| chp | 0 (0) | 3 (23) | 0.05 |
| scn | 17 (85) | 11 (84.6) | NS |
| ACME clusterb | 0 (0) | 0 (0) | NS |
| Accessory Gene Regulators | |||
| agr1 | 0 (0) | 9 (69.2) | < 0.001 |
| agr2 | 0 (0) | 1 (7.6) | NS |
| agr3 | 20 (100) | 3 (23) | < 0.001 |
| agr4 | 0 (0) | 0 (0) | NS |
| Resistance Genotyping | |||
| mecA | 20 (100) | 0 (0) | < 0.001 |
| mecC | 0 (0) | 0 (0) | NS |
| blaZ | 2 (10) | 10 (76.9) | < 0.001 |
| ermA | 0 (0) | 0 (0) | NS |
| ermC | 3 (15) | 0 (0) | NS |
| aphA3 | 20 (100) | 1 (7.6) | < 0.001 |
| sat | 20 (100) | 1 (7.6) | < 0.001 |
| fosB | 1 (5) | 3 (23) | NS |
| Virulence and Resistance Genes | TSST-1 + no. (%) | Et + no. (%) | Non Toxinogenic MSSA no. (%) | p valuea |
|---|---|---|---|---|
| Isolates | n = 62 | n = 33 | n = 203 | |
| Enterotoxins | ||||
| sea | 36 (58.0) | 26 (78.7) | 115 (56.6) | NS |
| seb | 7 (11.2) | 2 (6.0) | 14 (6.8) | NS |
| seg | 43 (69.3) | 30 (90.9) | 38 (18.7) | < 0.001 |
| seh | 18 (29.0) | 0 (0) | 16 (7.8) | 0.001 |
| sei | 62 (100) | 32 (96.9) | 179 (88.1) | NS |
| sek | 13 (20.9) | 2 (6.0) | 20 (9.8) | NS |
| seq | 13 (20.9) | 2 (6.0) | 16 (7.8) | NS |
| Other Toxins | ||||
| tst | 62 (100) | 6 (18.1) | 0 (0) | < 0.001 |
| etA | 2 (3.2) | 8 (24.2) | 0 (0) | < 0.001 |
| etB | 4 (6.4) | 25 (75.7) | 0 (0) | < 0.001 |
| etD | 0 (0) | 0 (0) | 0 (0) | NS |
| edinA | 0 (0) | 0 (0) | 0 (0) | NS |
| edinB | 24 (38.7) | 1 (3.0) | 18 (8.8) | < 0.001 |
| Leukocidins | ||||
| lukD | 62 (100) | 33 (100) | 203 (100) | NS |
| lukE | 62 (100) | 33 (100) | 203 (100) | NS |
| Hemolysins | ||||
| hla | 55 (88.7) | 33 (100) | 191 (94.0) | NS |
| hlb | 24 (38.7) | 1 (3.0) | 43 (21.1) | NS |
| hld | 62 (100) | 33 (100) | 203 (100) | NS |
| hlgA | 49 (79.0) | 33 (100) | 188 (92.6) | NS |
| hlgv | 60 (96.7) | 32 (96.9) | 197 (97.0) | NS |
| MSCRAMMs | ||||
| bbp | 62 (100) | 33 (100) | 203 (100) | NS |
| clfA | 62 (100) | 33 (100) | 203 (100) | NS |
| clfB | 62 (100) | 33 (100) | 203 (100) | NS |
| cna | 0 (0) | 0 (0) | 14 (6.8) | 0.006 |
| ebpS | 62 (100) | 33 (100) | 203 (100) | NS |
| fib | 55 (88.7) | 32 (96.9) | 187 (92.1) | NS |
| fnbA | 10 (16.1) | 7 (21.2) | 26 (12.8) | NS |
| fnbB | 28 (45.1) | 6 (18.1) | 62 (30.5) | NS |
| Capsule Components | ||||
| cap5 | 35 (56.5) | 20 (60.6) | 138 (68.0) | NS |
| cap8 | 27 (44.5) | 13 (39.4) | 65 (32.0) | NS |
| Intracellular Adhesion Polysaccharide | ||||
| icaA | 62 (100) | 33 (100) | 203 (100) | NS |
| icaC | 62 (100) | 33 (100) | 203 (100) | NS |
| icaD | 62 (100) | 33 (100) | 203 (100) | NS |
| Other | ||||
| ACME clusterb | 0 (0) | 0 (0) | 0 (0) | NS |
| Accessory Gene Regulators | ||||
| agr1 | 42 (67.7) | 25 (75.7) | 153 (75.3) | NS |
| agr2 | 6 (9.6) | 3 (9.0) | 31 (15.2) | NS |
| agr3 | 14 (22.5) | 2 (6.0) | 19 (9.3) | NS |
| agr4 | 0 (0) | 3 (9.0) | 0 (0) | NS |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mairi, A.; Touati, A.; Pantel, A.; Zenati, K.; Martinez, A.Y.; Dunyach-Remy, C.; Sotto, A.; Lavigne, J.-P. Distribution of Toxinogenic Methicillin-Resistant and Methicillin-Susceptible Staphylococcus aureus from Different Ecological Niches in Algeria. Toxins 2019, 11, 500. https://doi.org/10.3390/toxins11090500
Mairi A, Touati A, Pantel A, Zenati K, Martinez AY, Dunyach-Remy C, Sotto A, Lavigne J-P. Distribution of Toxinogenic Methicillin-Resistant and Methicillin-Susceptible Staphylococcus aureus from Different Ecological Niches in Algeria. Toxins. 2019; 11(9):500. https://doi.org/10.3390/toxins11090500
Chicago/Turabian StyleMairi, Assia, Abdelaziz Touati, Alix Pantel, Karima Zenati, Alex Yahiaoui Martinez, Catherine Dunyach-Remy, Albert Sotto, and Jean-Philippe Lavigne. 2019. "Distribution of Toxinogenic Methicillin-Resistant and Methicillin-Susceptible Staphylococcus aureus from Different Ecological Niches in Algeria" Toxins 11, no. 9: 500. https://doi.org/10.3390/toxins11090500
APA StyleMairi, A., Touati, A., Pantel, A., Zenati, K., Martinez, A. Y., Dunyach-Remy, C., Sotto, A., & Lavigne, J.-P. (2019). Distribution of Toxinogenic Methicillin-Resistant and Methicillin-Susceptible Staphylococcus aureus from Different Ecological Niches in Algeria. Toxins, 11(9), 500. https://doi.org/10.3390/toxins11090500