Toxicological Characterization and Phospholipase D Activity of the Venom of the Spider Sicarius thomisoides

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Description of the Captured S. thomisoides and Determination of the Venom Protein Concentration
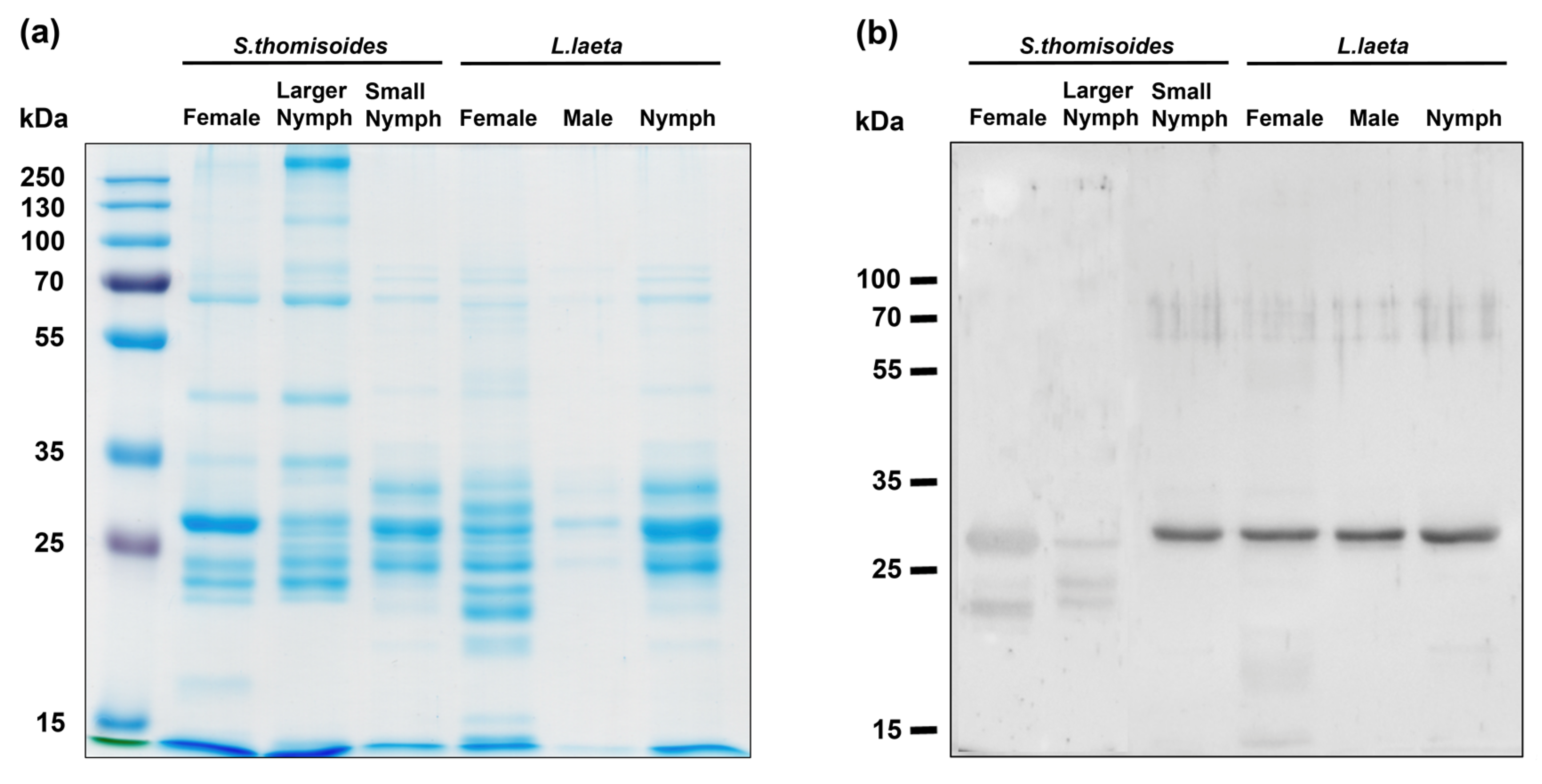
2.2. Electrophoretic Characterization of Venom from the S. thomisoides Spider and Evaluation of Phospholipase D activity
2.3. The Venom of S. thomisoides Spiders Induces Hemolysis, Cytotoxicity, and Dermonecrosis
3. Discussion
4. Materials and Methods
4.1. Spiders and Venoms
4.2. Electrophoresis and Immunoblotting
4.3. Phospholipase D Activity
4.4. Complement-Dependent Hemolysis Assay
4.5. Viability Assay
4.6. Dermonecrosis Assay
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- WSC. World spider catalog. Version 21.0. Natural History Museum Bern. 2020. Available online: http://wsc.nmbe.ch (accessed on 7 July 2020).
- Binford, G.J.; Bodner, M.R.; Cordes, M.H.; Baldwin, K.L.; Rynerson, M.R.; Burns, S.N.; Zobel-Thropp, P.A. Molecular evolution, functional variation, and proposed nomenclature of the gene family that includes sphingomyelinase D in Sicariid spider venoms. Mol. Biol. Evol. 2009, 26, 547–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gremski, L.H.; Trevisan-Silva, D.; Ferrer, V.P.; Matsubara, F.H.; Meissner, G.O.; Wille, A.C.; Vuitika, L.; Dias-Lopes, C.; Ullah, A.; de Moraes, F.R.; et al. Recent advances in the understanding of brown spider venoms: From the biology of spiders to the molecular mechanisms of toxins. Toxicon 2014, 83, 91–120. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, P.H.; da Silveira, R.B.; Appel, M.H.; Mangili, O.C.; Gremski, W.; Veiga, S.S. Brown spiders and loxoscelism. Toxicon 2004, 44, 693–709. [Google Scholar] [CrossRef]
- Swanson, D.L.; Vetter, R.S. Loxoscelism. Clin. Dermatol. 2006, 24, 213–221. [Google Scholar] [CrossRef]
- Chaim, O.M.; Trevisan-Silva, D.; Chaves-Moreira, D.; Wille, A.C.; Ferrer, V.P.; Matsubara, F.H.; Mangili, O.C.; da Silveira, R.B.; Gremski, L.H.; Gremski, W.; et al. Brown spider (Loxosceles genus) venom toxins: Tools for biological purposes. Toxins 2011, 3, 309–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walckenaer, C.A. Dernier supplément. In Histoire Naturelle des Insectes: Aptères; Walckenaer, C.A., Gervais, P., Eds.; Librairie Encyclopédique de Roret: Paris, France, 1847; pp. 365–564. [Google Scholar]
- Magalhaes, I.L.F.; Brescovit, A.D.; Santos, A.J. Phylogeny of Sicariidae spiders (Araneae: Haplogynae), with a monograph on neotropical Sicarius. Zool. J. Linn. Soc. 2017, 179, 767–864. [Google Scholar]
- Taucare-Rios, A.; Veloso, C.; Bustamante, R.O. Microhabitat selection in the sand recluse spider (Sicarius thomisoides): The effect of rock size and temperature. J. Nat. Hist. 2017, 51, 2199–2210. [Google Scholar] [CrossRef]
- Taucare-Rios, A.; Piel, W.H. Predation on the gecko Phyllodactylus gerrhopygus (Wiegmann) (Squamata: Gekkonidae) by the six-eyed sand spider Sicarius thomisoides (Walckenaer) (Araneae: Sicariidae). Rev. Soc. Entomol. Argent. 2020, 79, 48–51. [Google Scholar] [CrossRef]
- Binford, G.J.; Wells, M.A. The phylogenetic distribution of sphingomyelinase D activity in venoms of Haplogyne spiders. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2003, 135, 25–33. [Google Scholar] [CrossRef]
- Zobel-Thropp, P.A.; Bodner, M.R.; Binford, G.J. Comparative analyses of venoms from American and African Sicarius spiders that differ in sphingomyelinase D activity. Toxicon 2010, 55, 1274–1282. [Google Scholar] [CrossRef]
- Gremski, L.H.; da Justa, H.C.; da Silva, T.P.; Polli, N.L.C.; Antunes, B.C.; Minozzo, J.C.; Wille, A.C.M.; Senff-Ribeiro, A.; Arni, R.K.; Veiga, S.S. Forty years of the description of brown spider venom phospholipases-D. Toxins 2020, 12, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, P.H.; Bertani, R.; Goncalves-de-Andrade, R.M.; Nagahama, R.H.; van den Berg, C.W.; Tambourgi, D.V. Venom of the brazilian spider Sicarius ornatus (Araneae, Sicariidae) contains active sphingomyelinase D: Potential for toxicity after envenomation. PLoS Negl. Trop. Dis. 2013, 7, e2394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schenone, H. Toxic pictures produced spiders bites in Chile: Latrodectism and loxoscelism. Rev. Med. Chil. 2003, 131, 437–444. [Google Scholar] [PubMed]
- Taucare-Rios, A. Synantropic dangerous spiders from Chile. Rev. Med. Chil. 2012, 140, 1228–1229. [Google Scholar] [PubMed] [Green Version]
- De Oliveira, C.K.; Goncalves de Andrade, R.M.; Giusti, A.L.; Dias da Silva, W.; Tambourgi, D.V. Sex-linked variation of Loxosceles intermedia spider venoms. Toxicon 1999, 37, 217–221. [Google Scholar] [CrossRef]
- De Oliveira, K.C.; Goncalves de Andrade, R.M.; Piazza, R.M.; Ferreira, J.M., Jr.; van den Berg, C.W.; Tambourgi, D.V. Variations in Loxosceles spider venom composition and toxicity contribute to the severity of envenomation. Toxicon 2005, 45, 421–429. [Google Scholar] [CrossRef]
- Pekar, S.; Bocanek, O.; Michalek, O.; Petrakova, L.; Haddad, C.R.; Sedo, O.; Zdrahal, Z. Venom gland size and venom complexity-essential trophic adaptations of venomous predators: A case study using spiders. Mol. Ecol. 2018, 27, 4257–4269. [Google Scholar] [CrossRef]
- Binford, G.J.; Callahan, M.S.; Bodner, M.R.; Rynerson, M.R.; Nunez, P.B.; Ellison, C.E.; Duncan, R.P. Phylogenetic relationships of Loxosceles and Sicarius spiders are consistent with Western Gondwanan vicariance. Mol. Phylogenet. Evol. 2008, 49, 538–553. [Google Scholar] [CrossRef]
- Aran-Sekul, T.; Rojas, J.M.; Subiabre, M.; Cruz, V.; Cortes, W.; Osorio, L.; Gonzalez, J.; Araya, J.E.; Catalan, A. Heterophilic antibodies in sera from individuals without loxoscelism cross-react with phospholipase D from the venom of Loxosceles and Sicarius spiders. J. Venom Anim. Toxins Incl. Trop. Dis. 2018, 24, 18. [Google Scholar] [CrossRef]
- Kurpiewski, G.; Forrester, L.J.; Barrett, J.T.; Campbell, B.J. Platelet aggregation and sphingomyelinase D activity of a purified toxin from the venom of Loxosceles reclusa. Biochim. Biophys. Acta 1981, 678, 467–476. [Google Scholar] [CrossRef]
- Tambourgi, D.V.; Magnoli, F.C.; van den Berg, C.W.; Morgan, B.P.; de Araujo, P.S.; Alves, E.W.; Da Silva, W.D. Sphingomyelinases in the venom of the spider Loxosceles intermedia are responsible for both dermonecrosis and complement-dependent hemolysis. Biochem. Biophys. Res. Commun. 1998, 251, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Van Meeteren, L.A.; Frederiks, F.; Giepmans, B.N.; Pedrosa, M.F.; Billington, S.J.; Jost, B.H.; Tambourgi, D.V.; Moolenaar, W.H. Spider and bacterial sphingomyelinases D target cellular lysophosphatidic acid receptors by hydrolyzing lysophosphatidylcholine. J. Biol. Chem. 2004, 279, 10833–10836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Lynch, K.R. Brown recluse spider (Loxosceles reclusa) venom phospholipase D (PLD) generates lysophosphatidic acid (LPA). Biochem. J. 2005, 391, 317–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, L.F.; Laugesen, S.; Botelho, E.D.; Ricart, C.A.; Fontes, W.; Barbaro, K.C.; Roepstorff, P.; Sousa, M.V. Proteome analysis of brown spider venom: Identification of loxnecrogin isoforms in Loxosceles gaucho venom. Proteomics 2005, 5, 2167–2176. [Google Scholar] [CrossRef]
- Kalapothakis, E.; Chatzaki, M.; Goncalves-Dornelas, H.; de Castro, C.S.; Silvestre, F.G.; Laborne, F.V.; de Moura, J.F.; Veiga, S.S.; Chavez-Olortegui, C.; Granier, C.; et al. The loxtox protein family in Loxosceles intermedia (Mello-Leitao) venom. Toxicon 2007, 50, 938–946. [Google Scholar] [CrossRef]
- Murakami, M.T.; Fernandes-Pedrosa, M.F.; de Andrade, S.A.; Gabdoulkhakov, A.; Betzel, C.; Tambourgi, D.V.; Arni, R.K. Structural insights into the catalytic mechanism of sphingomyelinases D and evolutionary relationship to glycerophosphodiester phosphodiesterases. Biochem. Biophys. Res. Commun. 2006, 342, 323–329. [Google Scholar] [CrossRef]
- De Giuseppe, P.O.; Ullah, A.; Silva, D.T.; Gremski, L.H.; Wille, A.C.; Chaves Moreira, D.; Ribeiro, A.S.; Chaim, O.M.; Murakami, M.T.; Veiga, S.S.; et al. Structure of a novel class II phospholipase D: Catalytic cleft is modified by a disulphide bridge. Biochem. Biophys. Res. Commun. 2011, 409, 622–627. [Google Scholar] [CrossRef] [Green Version]
- Paixao-Cavalcante, D.; van den Berg, C.W.; de Freitas Fernandes-Pedrosa, M.; Goncalves de Andrade, R.M.; Tambourgi, D.V. Role of matrix metalloproteinases in HaCaT keratinocytes apoptosis induced by Loxosceles venom sphingomyelinase D. J. Invest. Dermatol. 2006, 126, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Tambourgi, D.V.; Morgan, B.P.; de Andrade, R.M.; Magnoli, F.C.; van Den Berg, C.W. Loxosceles intermedia spider envenomation induces activation of an endogenous metalloproteinase, resulting in cleavage of glycophorins from the erythrocyte surface and facilitating complement-mediated lysis. Blood 2000, 95, 683–691. [Google Scholar] [CrossRef]
- Ponec, M.; Havekes, L.; Kempenaar, J.; Vermeer, B.J. Cultured human skin fibroblasts and keratinocytes: Differences in the regulation of cholesterol synthesis. J. Invest. Dermatol. 1983, 81, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Van Aswegen, G.; Van Rooyen, J.M.; Van der Nest, D.G.; Veldman, F.J.; De Villiers, T.H.; Oberholzer, G. Venom of a six-eyed crab spider, Sicarius testaceus (Purcell, 1908), causes necrotic and haemorrhagic lesions in the rabbit. Toxicon 1997, 35, 1149–1152. [Google Scholar] [CrossRef]
- Lotz, L. An update on the spider genus Hexophthalma (Araneae: Sicariidae) in the afrotropical region, with descriptions of new species. Eur. J. Taxon. 2018, 424, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Dos-Santos, M.C.; Cardoso, J.L.C. Lesao dermonecrótica por Sicarius tropicus, simulando loxoscelismo cutaneo. Rev. Soc. Bras. Med. Trop. 1992, 25, 115–123. [Google Scholar]
- Monteiro, C.L.; Rubel, R.; Cogo, L.L.; Mangili, O.C.; Gremski, W.; Veiga, S.S. Isolation and identification of Clostridium perfringens in the venom and fangs of Loxosceles intermedia (brown spider): Enhancement of the dermonecrotic lesion in loxoscelism. Toxicon 2002, 40, 409–418. [Google Scholar] [CrossRef]
- Ospedal, K.Z.; Appel, M.H.; Fillus Neto, J.; Mangili, O.C.; Sanches Veiga, S.; Gremski, W. Histopathological findings in rabbits after experimental acute exposure to the Loxosceles intermedia (brown spider) venom. Int. J. Exp. Pathol. 2002, 83, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Gertsch, W.J. The spider genus Loxosceles in South America (Araneae Scytodidae). Bull. Am. Mus. Nat. Hist. 1967, 136, 117–174. [Google Scholar]
- Catalan, A.; Espoz, M.C.; Cortes, W.; Sagua, H.; Gonzalez, J.; Araya, J.E. Tetracycline and penicillin resistant Clostridium perfringens isolated from the fangs and venom glands of Loxosceles laeta: Its implications in loxoscelism treatment. Toxicon 2010, 56, 890–896. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Catalan, A.; Cortes, W.; Sagua, H.; Gonzalez, J.; Araya, J.E. Two new phospholipase D isoforms of Loxosceles laeta: Cloning, heterologous expression, functional characterization, and potential biotechnological application. J. Biochem. Mol. Toxicol. 2011, 25, 393–403. [Google Scholar] [CrossRef]
- Rojas, J.M.; Aran-Sekul, T.; Cortes, E.; Jaldin, R.; Ordenes, K.; Orrego, P.R.; Gonzalez, J.; Araya, J.E.; Catalan, A. Phospholipase D from Loxosceles laeta spider venom induces IL-6, IL-8, CXCL1/GRO-alpha, and CCL2/MCP-1 production in human skin fibroblasts and stimulates monocytes migration. Toxins 2017, 9, 125. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. Nih image to imagej: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arán-Sekul, T.; Perčić-Sarmiento, I.; Valencia, V.; Olivero, N.; Rojas, J.M.; Araya, J.E.; Taucare-Ríos, A.; Catalán, A. Toxicological Characterization and Phospholipase D Activity of the Venom of the Spider Sicarius thomisoides. Toxins 2020, 12, 702. https://doi.org/10.3390/toxins12110702
Arán-Sekul T, Perčić-Sarmiento I, Valencia V, Olivero N, Rojas JM, Araya JE, Taucare-Ríos A, Catalán A. Toxicological Characterization and Phospholipase D Activity of the Venom of the Spider Sicarius thomisoides. Toxins. 2020; 12(11):702. https://doi.org/10.3390/toxins12110702
Chicago/Turabian StyleArán-Sekul, Tomás, Ivanka Perčić-Sarmiento, Verónica Valencia, Nelly Olivero, José M. Rojas, Jorge E. Araya, Andrés Taucare-Ríos, and Alejandro Catalán. 2020. "Toxicological Characterization and Phospholipase D Activity of the Venom of the Spider Sicarius thomisoides" Toxins 12, no. 11: 702. https://doi.org/10.3390/toxins12110702
APA StyleArán-Sekul, T., Perčić-Sarmiento, I., Valencia, V., Olivero, N., Rojas, J. M., Araya, J. E., Taucare-Ríos, A., & Catalán, A. (2020). Toxicological Characterization and Phospholipase D Activity of the Venom of the Spider Sicarius thomisoides. Toxins, 12(11), 702. https://doi.org/10.3390/toxins12110702