Chitosan as a Coagulant to Remove Cyanobacteria Can Cause Microcystin Release




Abstract
:1. Introduction
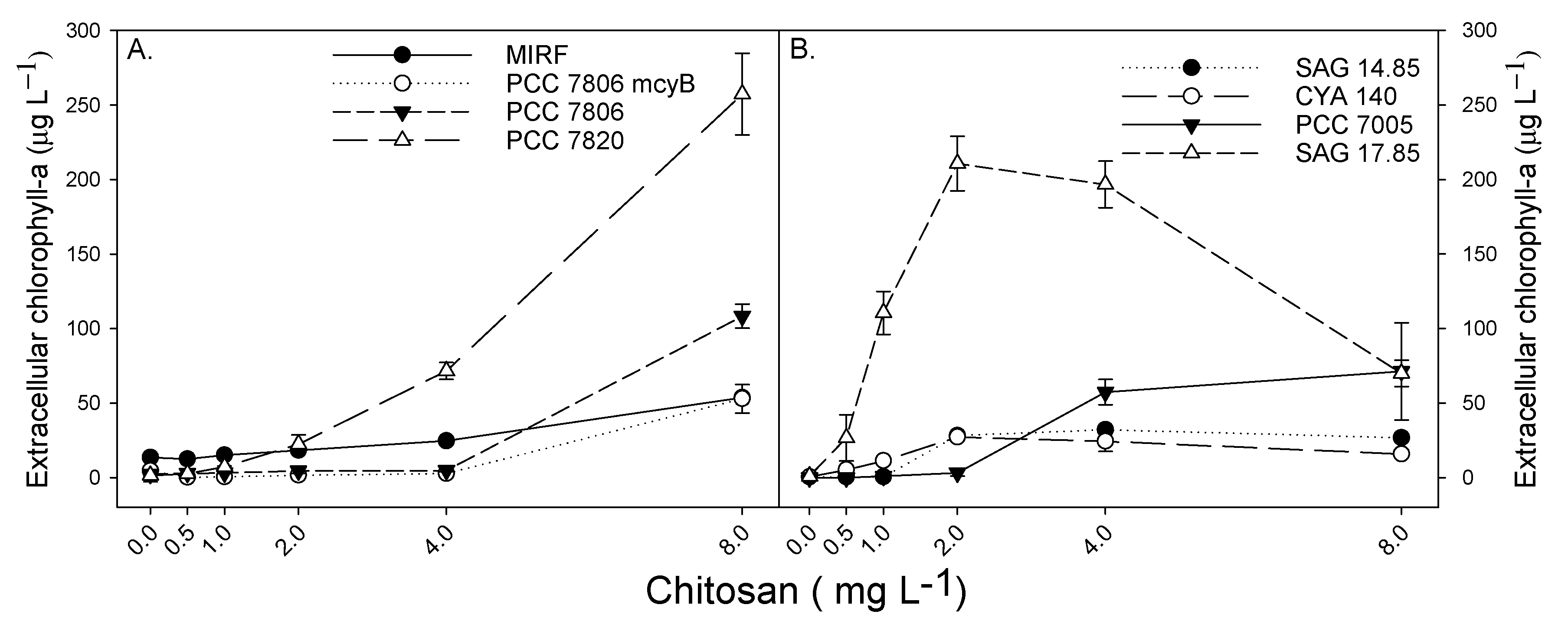
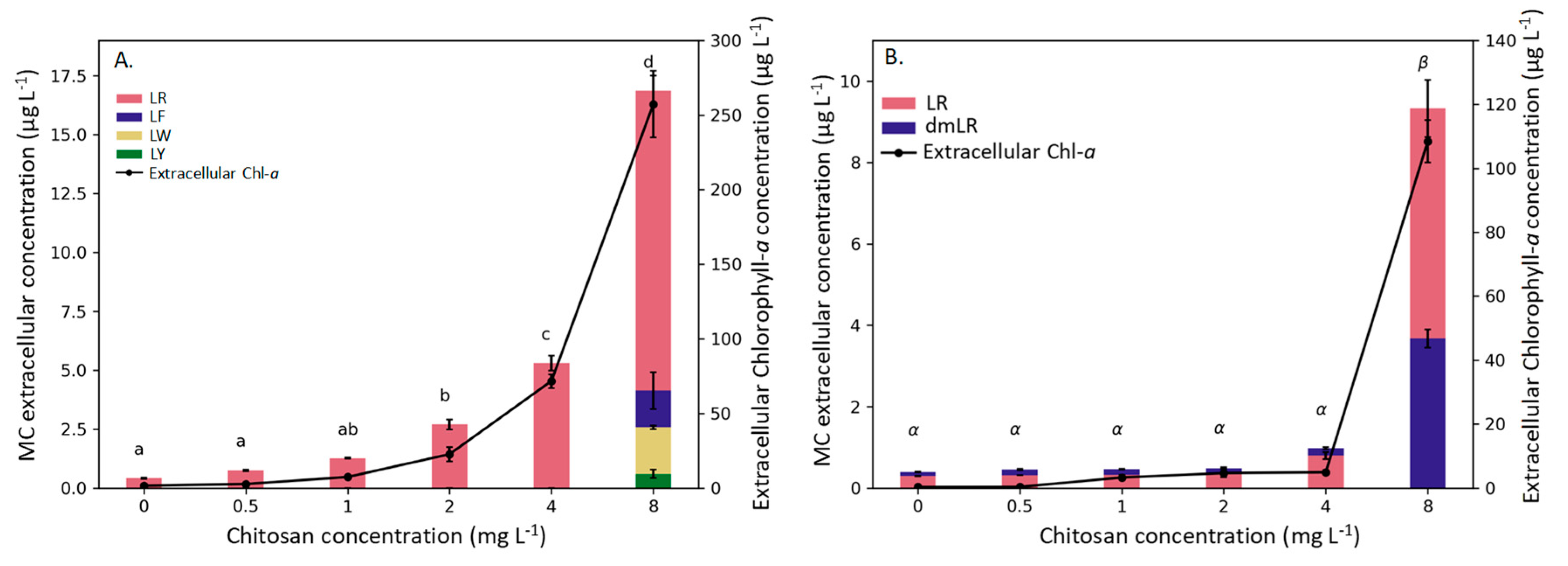
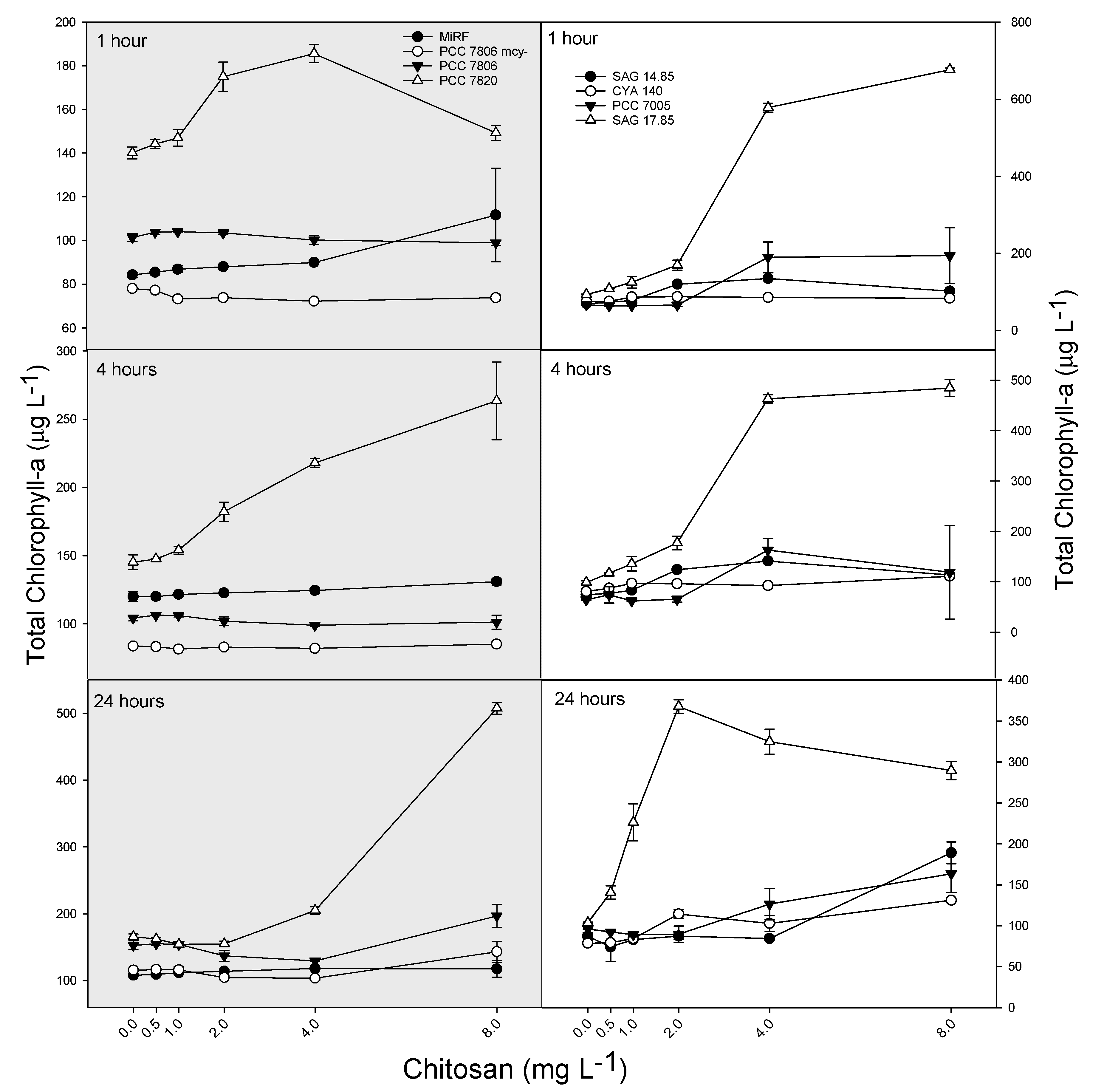
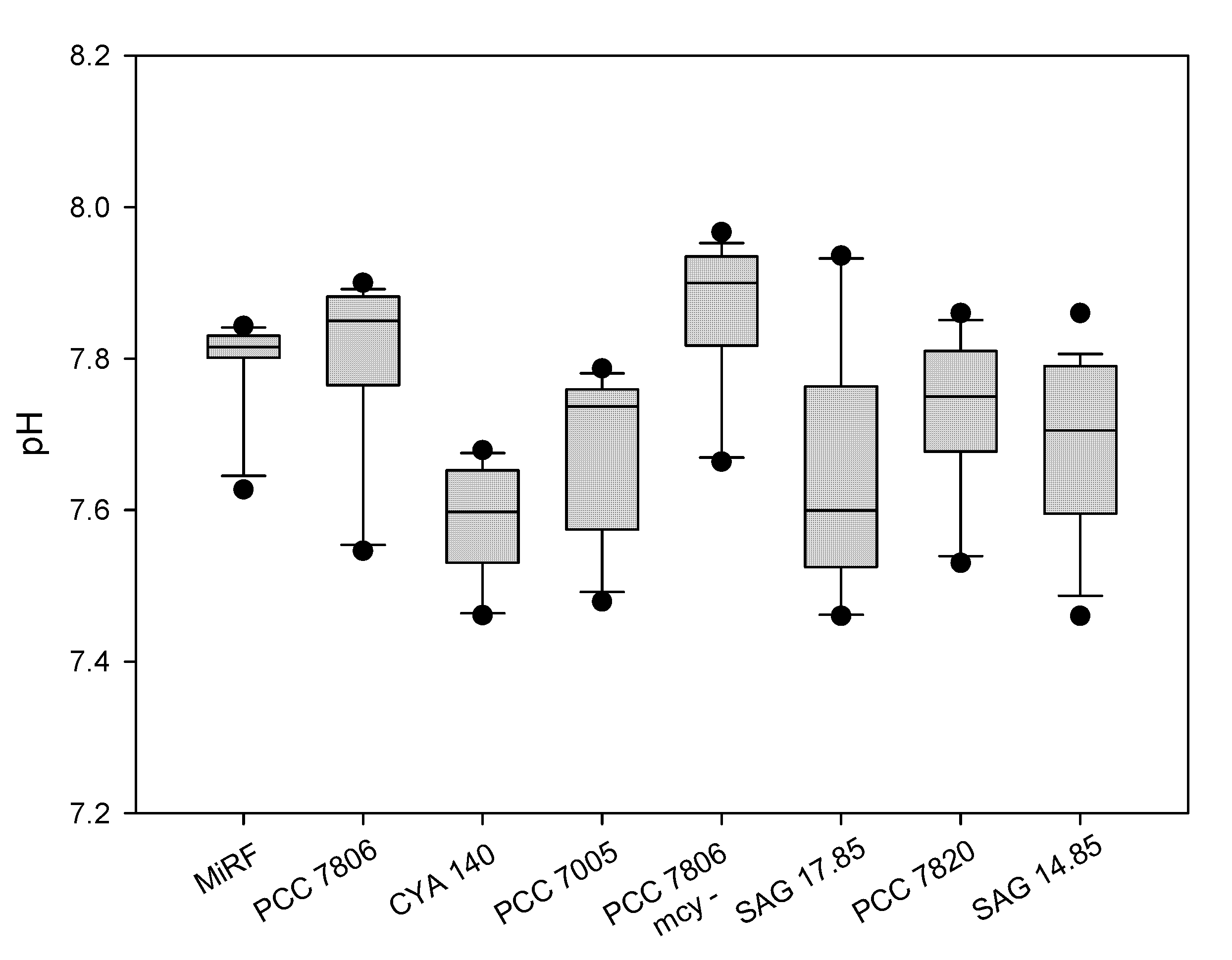
2. Results
3. Discussion
4. Materials and Methods
4.1. Microcystis aeruginosa Cultures
4.2. Chitosan
4.3. Experiment Design
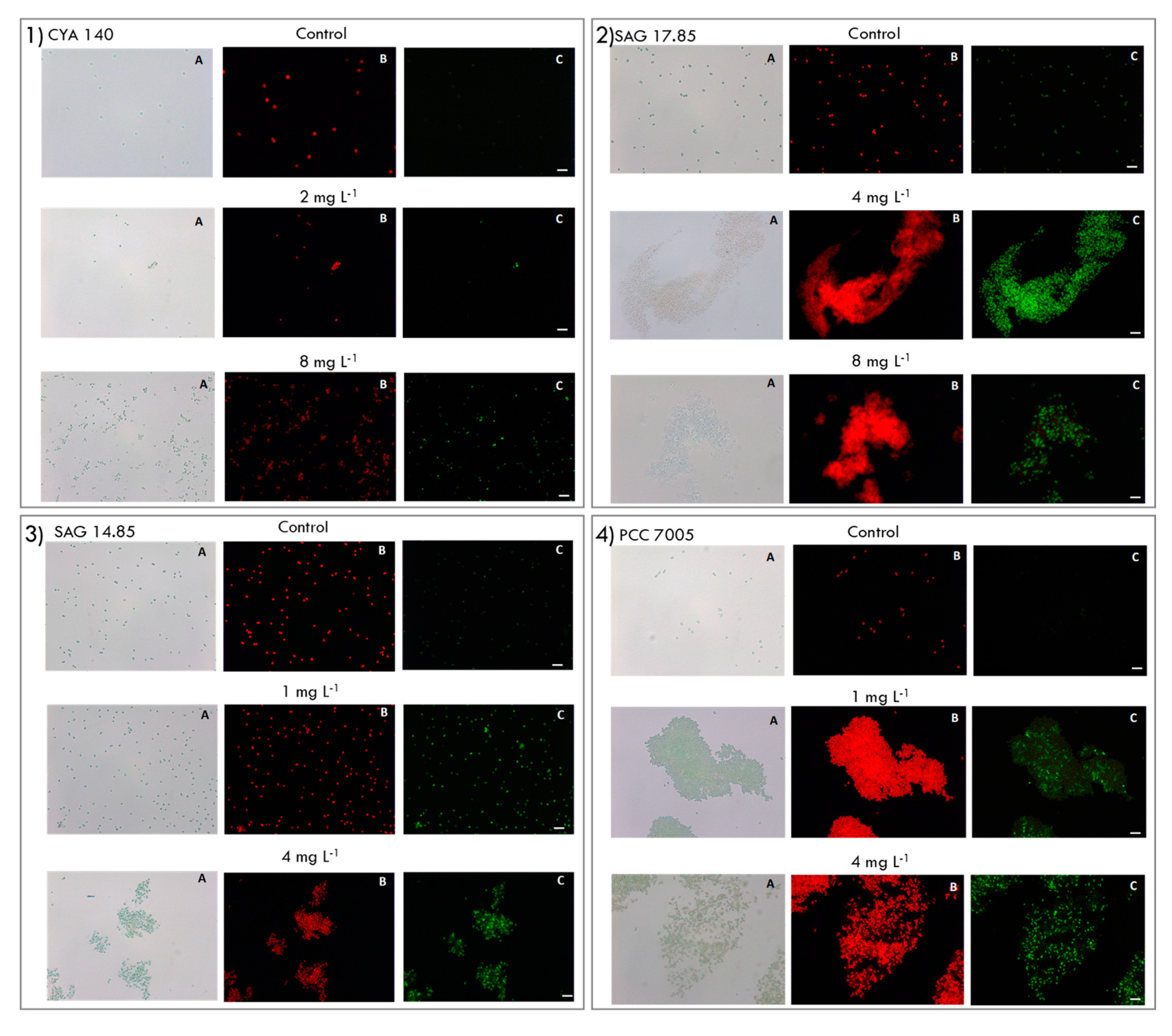
4.4. Cell Membrane Permeability
4.5. Matrix Effect on MC Analysis
4.6. Data Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| M. aeruginosa Strain | Exposure Duration | ||
|---|---|---|---|
| 1 h | 4 h | 24 h | |
| MiRF-1 | H5 = 9.93; p = 0.077 | F5,12 = 4.65; p = 0.014 * | F5,12 = 13.87; p < 0.001 * |
| PCC 7806 ΔmcyB | H5 = 16.70; p = 0.005 * | F5,12 = 66.34; p < 0.001 * | H5 = 12.70; p = 0.026 * |
| PCC 7806 | H5 = 12.24; p = 0.032 * | H5 = 11.28; p = 0.046 * | H5 = 15.81; p = 0.007 * |
| PCC 7820 | F5,12 = 89.62; p < 0.001 * | F5,12 = 47.06; p < 0.001 * | F5,12 = 1079.0 p < 0.001 * |
| SAG 14.85 | H5 = 16.317; p = 0.006 * | F5,12 = 876.16; p < 0.001 * | H5 = 16.1; p = 0.007 * |
| CYA 140 | F5,12 = 149.94; p < 0.001 * | F5,12 = 144.96; p < 0.001 * | F5,12 = 338.1; p < 0.001 * |
| PCC 7005 | H5 = 14.13; p = 0.015 * | F5,12 = 187.49; p < 0.001 * | H5 = 15.16; p = 0.01 * |
| SAG 17.85 | H5 = 16.74; p = 0.005 * | H5 = 16.74; p = 0.005 * | H5 = 16.56; p = 0.005 * |






| Literature Available | Endpoints | Effect |
|---|---|---|
| [46] | PSII efficiency and Membrane permeability (MP) | − |
| [47] | K+ release and dissolved toxins | − |
| [78] | Phycocyanin and allophycocyanin release | − |
| [48] | K+/M2+ release and dissolved toxins | − |
| This study | PSII efficiency, dissolved toxins, MP and FChl-a | − |
| [40] | PSII efficiency | 0 |
| [79] | PSII efficiency | 0 |
| [35] | Dissolved toxins | 0 |
| [21] | Cell viability and recovery | 0 |
| [55] | PSII efficiency | 0 |
| [66] | K+ release | 0 |
| [80] | Growth | 0 |
| [45] | PSII efficiency | 0 |
| [42] | PSII efficiency | 0 |
| [41] | PSII efficiency and dissolved toxins | 0 and − |
| [43] | Dissolved toxins | 0 and − |
| [81] | NR | NR |
| [82] | NR | NR |
| [83] | NR | NR |
| [84] | NR | NR |
| [35] | NR | NR |
| [85] | NR | NR |
| [86] | NR | NR |
| [38] | NR | NR |
| [87] | NR | NR |
| [88] | NR | NR |
| [37] | NR | NR |
| [36] | NR | NR |
| [25] | NR | NR |
| [26] | NR | NR |
| [89] | NR | NR |
References
- Gadd, G.M.; Raven, J.A. Geomicrobiology of Eukaryotic Microorganisms. Geomicrobiol. J. 2010, 27, 491–519. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Smith, V.H.; Schindler, D.W. Eutrophication science: Where do we go from here? Trends Ecol. Evol. 2009, 24, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, S.M.F.; Carmichael, W.W.; Jochimsen, E.M.; Rinehart, K.L.; Lau, S.; Shaw, G.R.; Eaglesham, G.K. Human intoxication by microcystins during renal dialysis treatment in Caruaru—Brazil. Toxicology 2002, 181–182, 441–446. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Azevedo, S.M.F.O.; An, J.S.; Molica, R.J.R.; Jochimsen, E.M.; Lau, S.; Rinehart, K.L.; Shaw, G.R.; Eaglesham, G.K. Human Fatalities from Cyanobacteria: Chemical and Biological Evidence for Cyanotoxins. Environ. Health Perspect. 2001, 109, 663. [Google Scholar] [CrossRef] [PubMed]
- Lürling, M.; Faassen, E.J. Dog poisonings associated with a Microcystis aeruginosa bloom in The Netherlands. Toxins 2013, 5, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Dodds, W.K.; Bouska, W.W.; Eitzmann, J.L.; Pilger, T.J.; Pitts, K.L.; Riley, A.J.; Schloesser, J.T.; Thornbrugh, D.J. Eutrophication of U.S. Freshwaters: Analysis of Potential Economic Damages. Environ. Sci. Technol. 2009, 43, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, D.P.; Wood, S.A.; Dietrich, D.R.; Puddick, J. Costs of harmful blooms of freshwater cyanobacteria. In Cyanobacteria; John Wiley & Sons, Ltd.: Chichester, UK, 2013; pp. 245–256. ISBN 9781118402238. [Google Scholar]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Cooke, G.D.; Welch, E.B.; Peterson, S.A.; Nichols, S. Restoration and Management of Lakes and Reservoirs, 3rd ed.; CRC Press-Taylor & Francis Group: Boca Raton, FL, USA, 2005; ISBN 1420032100. [Google Scholar]
- Hilt, S.; Gross, E.M.; Hupfer, M.; Morscheid, H.; Mählmann, J.; Melzer, A.; Poltz, J.; Sandrock, S.; Scharf, E.-M.; Schneider, S.; et al. Restoration of submerged vegetation in shallow eutrophic lakes—A guideline and state of the art in Germany. Limnol. Ecol. Manag. Inl. Waters 2006, 36, 155–171. [Google Scholar] [CrossRef] [Green Version]
- Paerl, H.W.; Barnard, M.A. Mitigating the global expansion of harmful cyanobacterial blooms: Moving targets in a human- and climatically-altered world. Harmful Algae 2020, 96, 101845. [Google Scholar] [CrossRef]
- Huser, B.J.; Futter, M.; Lee, J.T.; Perniel, M. In-lake measures for phosphorus control: The most feasible and cost-effective solution for long-term management of water quality in urban lakes. Water Res. 2016, 97, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Lürling, M.; Van Oosterhout, F. Controlling eutrophication by combined bloom precipitation and sediment phosphorus inactivation. Water Res. 2013, 47, 6527–6537. [Google Scholar] [CrossRef] [PubMed]
- Waajen, G.; van Oosterhout, F.; Douglas, G.; Lürling, M. Management of eutrophication in Lake De Kuil (The Netherlands) using combined flocculant—Lanthanum modified bentonite treatment. Water Res. 2016, 97, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, S.R. Eutrophication of aquatic ecosystems: Bistability and soil phosphorus. PNAS 2005, 102, 10002–10005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fastner, J.; Abella, S.; Litt, A.; Morabito, G.; Vörös, L.; Pálffy, K.; Straile, D.; Kümmerlin, R.; Matthews, D.; Phillips, M.G.; et al. Combating cyanobacterial proliferation by avoiding or treating inflows with high P load—experiences from eight case studies. Aquat. Ecol. 2016, 50, 367–383. [Google Scholar] [CrossRef] [Green Version]
- Lürling, M.; Mucci, M. Mitigating eutrophication nuisance: In-lake measures are becoming inevitable in eutrophic waters in The Netherlands. Hydrobiologia 2020. [Google Scholar] [CrossRef]
- Lürling, M.; Kang, L.; Mucci, M.; van Oosterhout, F.; Noyma, N.P.; Miranda, M.; Huszar, V.L.M.; Waajen, G.; Manzi, M. Coagulation and precipitation of cyanobacterial blooms. Ecol. Eng. 2020, 158. [Google Scholar] [CrossRef]
- Mucci, M.; Waajen, G.; van Oosterhout, F.; Yasseri, S.; Lürling, M. Whole lake application PAC-Phoslock treatment to manage eutrophication and cyanobacterial bloom. Inl. Waters 2020. under review. [Google Scholar]
- Li, L.; Pan, G. A Universal Method for Flocculating Harmful Algal Blooms in Marine and Fresh Waters Using Modified Sand. Environ. Sci. Technol. 2013, 47, 4555–4562. [Google Scholar] [CrossRef]
- Yang, R.; Li, H.; Huang, M.; Yang, H.; Li, A. A review on chitosan-based flocculants and their applications in water treatment. Water Res. 2016, 95, 59–89. [Google Scholar] [CrossRef]
- Chen, G.; Zhao, L.; Qi, Y.; Cui, Y. Chitosan and Its Derivatives Applied in Harvesting Microalgae for Biodiesel Production: An Outlook. J. Nanomater. 2014, 2014. [Google Scholar] [CrossRef] [Green Version]
- Renault, F.; Sancey, B.; Badot, P.-M.; Crini, G. Chitosan for coagulation/flocculation processes—An eco-friendly approach. Eur. Polym. J. 2009, 45, 1337–1348. [Google Scholar] [CrossRef]
- Pan, G.; Chen, J.; Anderson, D.M. Modified local sands for the mitigation of harmful algal blooms. Harmful Algae 2011, 10, 381–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, G.; Zou, H.; Chen, H.; Yuan, X. Removal of harmful cyanobacterial blooms in Taihu Lake using local soils III. Factors affecting the removal efficiency and an in situ field experiment using chitosan-modified local soils. Environ. Pollut. 2006, 141, 206–212. [Google Scholar] [CrossRef]
- Allan, C.R.; Hadwigei, L.A. The Fungicidal Effect of Chitosan on Fungi of Varying Cell Wall Composition. Exp. Mycol. 1979, 3, 285–287. [Google Scholar] [CrossRef]
- Kendra, D.F.; Hadwiger~, L.A.; Kendra, D.E. Characterization of the smallest chitosan oligomer that is maximally antifungal to fusarium solani and elicits pisatin formation in Pisum sativum. Exp. Mycol. 1984, 8, 276–281. [Google Scholar] [CrossRef]
- Kong, M.; Chen, X.G.; Xing, K.; Park, H.J. Antimicrobial properties of chitosan and mode of action: A state of the art review. Int. J. Food Microbiol. 2010, 144, 51–63. [Google Scholar] [CrossRef] [PubMed]
- No, H.K.; Young Park, N.; Ho Lee, S.; Meyers, S.P. Antibacterial activity of chitosans and chitosan oligomers with different molecular weights. Int. J. Food Microbiol. 2002, 74, 65–72. [Google Scholar] [CrossRef]
- Sudarshan, N.R.; Hoover, D.G.; Knorr, D. Antibacterial action of chitosan. Food Biotechnol. 1992, 6, 257–272. [Google Scholar] [CrossRef]
- Jeon, Y.-J.; Kamil, J.Y.V.A.; Shahidi, F. Chitosan as an Edible Invisible Film for Quality Preservation of Herring and Atlantic Cod. J. Agric. food Chem. 2002, 50, 5167–5178. [Google Scholar] [CrossRef]
- Cao, Z.; Sun, Y. Chitosan-based rechargeable long-term antimicrobial and biofilm-controlling systems. J. Biomed. Mater. Res. Part A 2009, 89A, 960–967. [Google Scholar] [CrossRef]
- Campaniello, D.; Bevilacqua, A.; Sinigaglia, M.; Corbo, M.R. Chitosan: Antimicrobial activity and potential applications for preserving minimally processed strawberries. Food Microbiol. 2008, 25, 992–1000. [Google Scholar] [CrossRef]
- Li, H.; Pan, G. Simultaneous removal of harmful algal blooms and microcystins using microorganism- and chitosan-modified local soil. Environ. Sci. Technol. 2015, 49, 6249–6256. [Google Scholar] [CrossRef] [Green Version]
- Pan, G.; Dai, L.; Li, L.; He, L.; Li, H.; Bi, L.; Gulati, R.D. Reducing the Recruitment of Sedimented Algae and Nutrient Release into the Overlying Water Using Modified Soil/Sand Flocculation- Capping in Eutrophic Lakes. Environ. Sci. Technol. 2012, 46, 5077–5084. [Google Scholar] [CrossRef]
- Zou, H.; Pan, G.; Chen, H.; Yuan, X. Removal of cyanobacterial blooms in Taihu Lake using local soils. II. Effective removal of Microcystis aeruginosa using local soils and sediments modified by chitosan. Environ. Pollut. 2006, 141, 201–205. [Google Scholar] [CrossRef]
- Wang, L.; Pan, G.; Shi, W.; Wang, Z.; Zhang, H. Manipulating nutrient limitation using modified local soils: A case study at Lake Taihu (China). Water Res. 2016, 101, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.; Lei, L.; Xiao, L.; Han, B. Cyanobacterial removal by a red soil-based flocculant and its effect on zooplankton: An experiment with deep enclosures in a tropical reservoir in China. Environ. Sci. Pollut. Res. 2019, 26, 30663–30674. [Google Scholar] [CrossRef] [Green Version]
- de Magalhães, L.; Noyma, N.P.; Furtado, L.L.; Mucci, M.; van Oosterhout, F.; Huszar, V.L.M.; Marinho, M.M.; Lürling, M. Efficacy of Coagulants and Ballast Compounds in Removal of Cyanobacteria (Microcystis) from Water of the Tropical Lagoon Jacarepaguá (Rio de Janeiro, Brazil). Estuaries Coasts 2016, 1–13. [Google Scholar] [CrossRef]
- Miranda, M.; Noyma, N.; Pacheco, F.S.; de Magalhaes, L.; Pinto, E.; Santos, S.; Soares, M.F.A.; Huszar, V.L.; Lürling, M.; Marinho, M.M. The efficiency of combined coagulant and ballast to remove harmful cyanobacterial blooms in a tropical shallow system. Harmful Algae 2017, 65, 27–39. [Google Scholar] [CrossRef]
- Noyma, N.P.; De Magalhães, L.; Miranda, M.; Mucci, M.; Van Oosterhout, F.; Huszar, V.L.M.; Marinho, M.M.; Lima, E.R.A.; Lürling, M. Coagulant plus ballast technique provides a rapid mitigation of cyanobacterial nuisance. PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- de Lucena-Silva, D.; Molozzi, J.; dos Santos Severiano, J.; Becker, V.; de Lucena Barbosa, J.E. Removal efficiency of phosphorus, cyanobacteria and cyanotoxins by the “flock & sink” mitigation technique in semi-arid eutrophic waters. Water Res. 2019, 159, 262–273. [Google Scholar] [CrossRef]
- Park, Y.H.; Kim, S.; Kim, H.S.; Park, C.; Choi, Y.-E. Adsorption Strategy for Removal of Harmful Cyanobacterial Species Microcystis aeruginosa Using Chitosan Fiber. Sustainability 2020, 12, 4587. [Google Scholar] [CrossRef]
- Noyma, N.P.; de Magalhães, L.; Furtado, L.L.; Mucci, M.; van Oosterhout, F.; Huszar, V.L.M.; Marinho, M.M.; Lürling, M. Controlling cyanobacterial blooms through effective flocculation and sedimentation with combined use of flocculants and phosphorus adsorbing natural soil and modified clay. Water Res. 2016, 1–13. [Google Scholar] [CrossRef]
- Mucci, M.; Noyma, N.P.; de Magalhães, L.; Miranda, M.; van Oosterhout, F.; Guedes, I.A.; Huszar, V.L.M.; Marinho, M.M.; Lürling, M. Chitosan as coagulant on cyanobacteria in lake restoration management may cause rapid cell lysis. Water Res. 2017, 118, 121–130. [Google Scholar] [CrossRef]
- Pei, H.-Y.; Ma, C.-X.; Hu, W.-R.; Sun, F. The behaviors of Microcystis aeruginosa cells and extracellular microcystins during chitosan flocculation and flocs storage processes. Bioresour. Technol. 2014, 151, 314–322. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, C.; Wang, P.; Qian, J.; Hou, J.; Ao, Y.; Wu, B. The performance of chitosan/montmorillonite nanocomposite during the flocculation and floc storage processes of Microcystis aeruginosa cells. Env. Sci Pollut Res 2015, 22, 11148–11161. [Google Scholar] [CrossRef]
- Pan, G.; Yang, B.; Wang, D.; Chen, H.; Tian, B.; Zhang, M.; Yuan, X.; Chen, J. In-lake algal bloom removal and submerged vegetation restoration using modified local soils. Ecol. Eng. 2011, 37, 302–308. [Google Scholar] [CrossRef]
- Lürling, M.; Mackay, E.; Reitzel, K.; Spears, B. Editorial—A critical perspective on geo-engineering for eutrophication management in lakes. Water Res. 2016, 97, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baurès, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef]
- Harke, M.J.; Steffen, M.M.; Otten, T.G.; Wilhelm, S.W.; Wood, S.A.; Paerl, H.W. A review of the global ecology, genomics, and biogeography of the toxic cyanobacterium, Microcystis spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef] [Green Version]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Srivastava, A.; Singh, S.; Ahn, C.-Y.; Oh, H.-M.; Asthana, R.K. Monitoring Approaches for a Toxic Cyanobacterial Bloom. Environ. Sci. Technol. 2013, 47, 8999–9013. [Google Scholar] [CrossRef]
- Lürling, M.; Noyma, N.P.; de Magalhães, L.; Miranda, M.; Mucci, M.; van Oosterhout, F.; Huszar, V.L.M.; Marinho, M.M. Critical assessment of chitosan as coagulant to remove cyanobacteria. Harmful Algae 2017, 66, 1–12. [Google Scholar] [CrossRef]
- Liu, H.; Du, Y.; Wang, X.; Sun, L. Chitosan kills bacteria through cell membrane damage. Int. J. Food Microbiol. 2004, 95, 147–155. [Google Scholar] [CrossRef]
- Witt, H.T. Primary Reactions of Oxygenic Photosynthesis. Ber. Bunsenges. Phys. Chem 1996, 100, 1923–1942. [Google Scholar] [CrossRef]
- Forni, C.; Telo’, F.R.; Caiola, M.G. Comparative analysis of the polysaccharides produced by different species of Microcystis (Chroococcales, Cyanophyta). Phycologia 1997, 36, 181–185. [Google Scholar] [CrossRef]
- Xu, H.; Cai, H.; Yu, G.; Jiang, H. Insights into extracellular polymeric substances of cyanobacterium Microcystis aeruginosa using fractionation procedure and parallel factor analysis. Water Res. 2013, 47, 2005–2014. [Google Scholar] [CrossRef]
- Gao, L.; Pan, X.; Zhang, D.; Mu, S.; Lee, D.-J.; Halik, U.; Zhang, D. Extracellular polymeric substances buffer against the biocidal effect of H2O2 on the bloom-forming cyanobacterium Microcystis aeruginosa. Water Res. 2015, 69, 51–58. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, J.; Pan, X. Cadmium sorption by EPSs produced by anaerobic sludge under sulfate-reducing conditions. J. Hazard. Mater. 2006, 138, 589–593. [Google Scholar] [CrossRef]
- Ozturk, S.; Aslim, B. Relationship between chromium(VI) resistance and extracellular polymeric substances (EPS) concentration by some cyanobacterial isolates. Env. Sci. Pollut. Res. 2008, 15, 478–480. [Google Scholar] [CrossRef]
- Bai, L.; Xu, H.; Wang, C.; Deng, J.; Jiang, H. Extracellular polymeric substances facilitate the biosorption of phenanthrene on cyanobacteria Microcystis aeruginosa. Chemosphere 2016, 162, 172–180. [Google Scholar] [CrossRef]
- De Philippis, R.; Colica, G.; Micheletti, E. Exopolysaccharide-producing cyanobacteria in heavy metal removal from water: Molecular basis and practical applicability of the biosorption process. Appl. Microbiol. Biotechnol. 2011, 92, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.; Su, Y.; Chen, C.; Jia, G.; Wang, H.; Wu, J.C.G.; Lin, J. Relationship between antibacterial activity of chitosan and surface characteristics of cell wall. Acta Pharmacol. Sin. 2004, 25, 932–936. [Google Scholar] [PubMed]
- Ma, C.; Hu, W.; Pei, H.; Xu, H.; Pei, R. Enhancing integrated removal of Microcystis aeruginosa and adsorption of microcystins using chitosan-aluminum chloride combined coagulants: Effect of chemical dosing orders and coagulation mechanisms. Colloids Surfaces A Physicochem. Eng. Asp. 2016, 490, 258–267. [Google Scholar] [CrossRef]
- Lee, J.; Walker, H.W. Adsorption of microcystin-Lr onto iron oxide nanoparticles. Colloids Surfaces A Physicochem. Eng. Asp. 2011, 373, 94–100. [Google Scholar] [CrossRef]
- Shimizu, Y.; Hsu, C.-P.; Genenah, A. Structure of Saxitoxin in Solutions and Stereochemistry of Dihydrosaxitoxins1. J. Am. Chem. Soc 1981, 103, 605–609. [Google Scholar] [CrossRef]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Chorus, I., Bartram, J., Eds.; E & FN Spon: London, UK, 1999; ISBN 0419239308. [Google Scholar]
- Lürling, M.; Mucci, M.; Waajen, G. Removal of Positively Buoyant Planktothrix rubescens in Lake Restoration. Toxins 2020, 12, 700. [Google Scholar] [CrossRef]
- Tashyreva, D.; Elster, J.; Billi, D. A Novel Staining Protocol for Multiparameter Assessment of Cell Heterogeneity in Phormidium Populations (Cyanobacteria) Employing Fluorescent Dyes. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Geng, L.; Qin, B.; Yang, Z. Unicellular, Microcystis aeruginosa cannot revert back to colonial form after short-term exposure to natural conditions. Biochem. Syst. Ecol. 2013, 51, 104–108. [Google Scholar] [CrossRef]
- Lürling, M.; Beekman, W. Palmelloids formation in Chlamydomonas reinhardtii: Defence against rotifer predators? Ann. Limnol. Int. J. Limnol. 2006, 42, 65–72. [Google Scholar] [CrossRef]
- Marinho, M.M.; Souza, M.B.G.; Lürling, M. Light and phosphate competition between Cylindrospermopsis raciborskii and Microcystis aeruginosa is strain dependent. Microb. Ecol. 2013, 66, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Dittmann, E.; Neilan, B.A.; Erhard, M.; von Döhren, H.; Börner, T. Insertional mutagenesis of a peptide synthetase gene that is responsible for hepatotoxin production in the cyanobacterium Microcystis aeruginosa PCC 7806. Mol. Microbiol. 1997, 26, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Ger, K.A.; Faassen, E.J.; Pennino, M.G.; Lürling, M. Effect of the toxin (microcystin) content of Microcystis on copepod grazing. Harmful Algae 2016, 52, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Lürling, M.; Meng, D.; Faassen, E.J. Effects of hydrogen peroxide and ultrasound on biomass reduction and toxin release in the cyanobacterium, Microcystis aeruginosa. Toxins 2014, 6, 3260–3280. [Google Scholar] [CrossRef] [Green Version]
- Shao, J.; Wang, Z.; Liu, Y.; Liu, H.; Peng, L.; Wei, X.; Lei, M.; Li, R. Physiological responses of Microcystis aeruginosa NIES-843 (cyanobacterium) under the stress of chitosan modified kaolinite (CMK) loading. Ecotoxicology 2012, 21, 698–704. [Google Scholar] [CrossRef]
- Guo, P.; Liu, Y.; Liu, C. Effects of chitosan, gallic acid, and algicide on the physiological and biochemical properties of Microcystis flos-aquae. Environ. Sci. Pollut. Res. 2015, 22, 13514–13524. [Google Scholar] [CrossRef]
- Rojsitthisak, P.; Burut-Archanai, S.; Pothipongsa, A.; Powtongsook, S. Repeated phosphate removal from recirculating aquaculture system using cyanobacterium remediation and chitosan flocculation. Water Environ. J. 2017, 31, 598–602. [Google Scholar] [CrossRef]
- Capelete, B.C.; Brandão, C.C.S. Evaluation of trihalomethane formation in treatment of. Water Sci. Technol. Water Supply 2013, 13.4, 1167–1173. [Google Scholar] [CrossRef]
- Huang, Y.; Xu, L.; Han, R.; Wang, G.; Wang, J.; Jia, J.; Zhang, P.; Pang, Y. Using chitosan-modified clays to control black-bloom-induced black suspended matter in Taihu Lake: Deposition and resuspension of black matter/clay flocs. Harmful Algae 2015, 45, 33–39. [Google Scholar] [CrossRef]
- Lama, S.; Muylaert, K.; Karki, T.B.; Foubert, I.; Henderson, R.K.; Vandamme, D. Flocculation properties of several microalgae and a cyanobacterium species during ferric chloride, chitosan and alkaline flocculation. Bioresour. Technol. 2016, 220, 464–470. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Pan, G. Cyanobacterial bloom mitigation using proteins with high isoelectric point and chitosan-modified soil. J. Appl. Phycol. 2016, 28, 357–363. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Pei, H.; Hu, W.; Cheng, J.; Xu, H.; Jin, Y. Significantly enhanced dewatering performance of drinking water sludge from a coagulation process using a novel chitosan–aluminum chloride composite coagulant in the treatment of cyanobacteria-laden source water. RSC Adv. 2016, 6, 61047–61056. [Google Scholar] [CrossRef]
- Rakesh, S.; Saxena, S.; Dhar, D.W.; Prasanna, R.; Saxena, A.K. Comparative evaluation of inorganic and organic amendments for their flocculation efficiency of selected microalgae. J. Appl. Phycol. 2014, 26, 399–406. [Google Scholar] [CrossRef]
- Yan, Q.; Yu, Y.; Feng, W.; Pan, G.; Chen, H.; Chen, J.; Yang, B.; Li, X.; Zhang, X. Plankton Community Succession in Artificial Systems Subjected to Cyanobacterial Blooms Removal using Chitosan-Modified Soils. Microb. Ecol. 2009, 58, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Zhang, H.; Pan, G. Flocculation of cyanobacterial cells using coal fly ash modified chitosan. Water Res. 2016, 97, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Pandhal, J.; Choon, W.; Kapoore, R.; Russo, D.; Hanotu, J.; Wilson, I.; Desai, P.; Bailey, M.; Zimmerman, W.; Ferguson, A. Harvesting Environmental Microalgal Blooms for Remediation and Resource Recovery: A Laboratory Scale Investigation with Economic and Microbial Community Impact Assessment. Biology 2017, 7, 4. [Google Scholar] [CrossRef] [Green Version]




| M. aeruginosa Strain | EC50-24 h (mg L−1) |
|---|---|
| MiRF-1 | >8 |
| PCC 7806 ΔmcyB | >8 |
| PCC 7806 | >8 |
| PCC 7820 | 4.51 (0.37) p < 0.0001A |
| PCC 7005 | 3.44 (0.42) p < 0.0001B |
| SAG 14.85 | 1.71 (0.08) p < 0.0001C |
| CYA 140 | 1.06 (0.04) p < 0.0001C |
| SAG 17.85 | 0.47 (0.05) p < 0.0001C |
| Strain ID | Acquired from | Microcystins (MCs) Produced |
|---|---|---|
| MiRF-1 | Laboratory of Ecophysiology and Toxicology of Cyanobacteria (Brazil) | dm-MC-LR, MC-LR, MC-LY, MC- LW, MC-LF [74] |
| PCC 7806 ΔmcyB | Pasteur Culture Collection (France) | None [75] |
| PCC 7806 | Pasteur Culture Collection (France) | dm-MC-LR, MC-LR [76] |
| PCC 7005 | Pasteur Culture Collection (France) | None detected (this study) |
| PCC 7820 | Pasteur Culture Collection (France) | dm-MC-LR, MC-LR, MC-LY, MC-LW, MC-LF [77] |
| SAG 14.85 | Sammlung von Algenkulturen der Universität Göttingen (Germany) | dm-MC-LR, MC-LR (unpublished data) |
| SAG 17.85 | Sammlung von Algenkulturen der Universität Göttingen (Germany) | dm-MC-LR, MC-LR, MC-YR [76] |
| CYA 140 | Norwegian Institute for Water Research (Norway) | dm-MC-LR, MC-LR [76] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mucci, M.; Guedes, I.A.; Faassen, E.J.; Lürling, M. Chitosan as a Coagulant to Remove Cyanobacteria Can Cause Microcystin Release. Toxins 2020, 12, 711. https://doi.org/10.3390/toxins12110711
Mucci M, Guedes IA, Faassen EJ, Lürling M. Chitosan as a Coagulant to Remove Cyanobacteria Can Cause Microcystin Release. Toxins. 2020; 12(11):711. https://doi.org/10.3390/toxins12110711
Chicago/Turabian StyleMucci, Maíra, Iame A. Guedes, Elisabeth J. Faassen, and Miquel Lürling. 2020. "Chitosan as a Coagulant to Remove Cyanobacteria Can Cause Microcystin Release" Toxins 12, no. 11: 711. https://doi.org/10.3390/toxins12110711
APA StyleMucci, M., Guedes, I. A., Faassen, E. J., & Lürling, M. (2020). Chitosan as a Coagulant to Remove Cyanobacteria Can Cause Microcystin Release. Toxins, 12(11), 711. https://doi.org/10.3390/toxins12110711