Impact of the Escherichia coli Heat-Stable Enterotoxin b (STb) on Gut Health and Function




Abstract
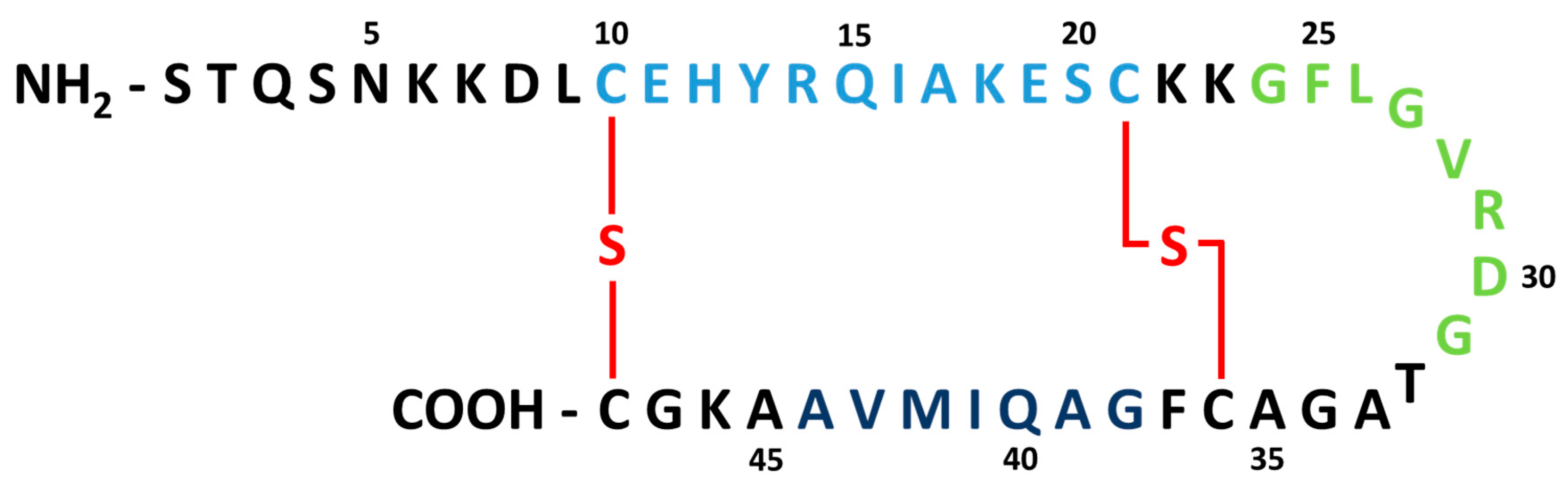
1. Introduction
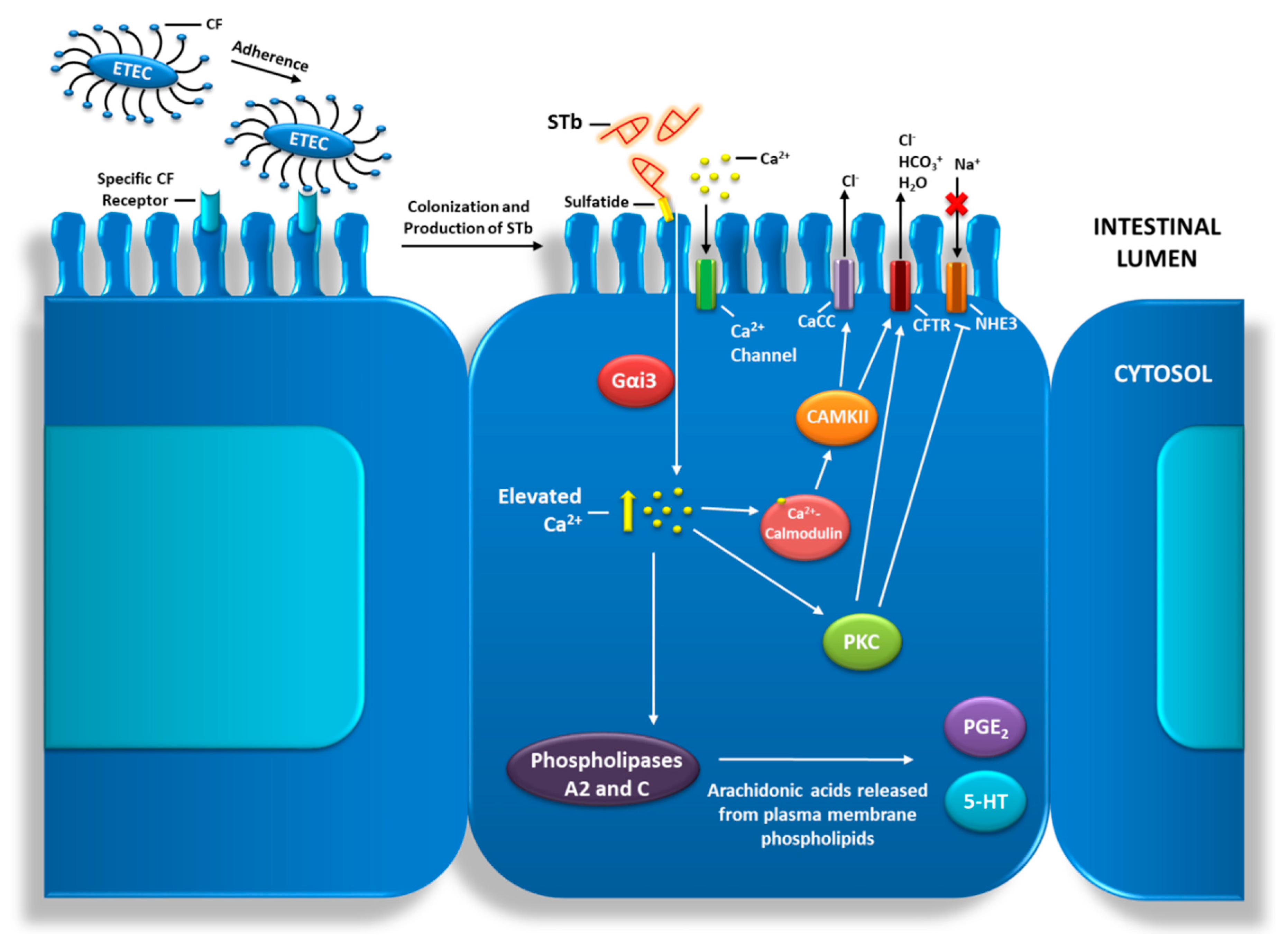
2. Impact of ETEC STb on Host Gut Health and Function
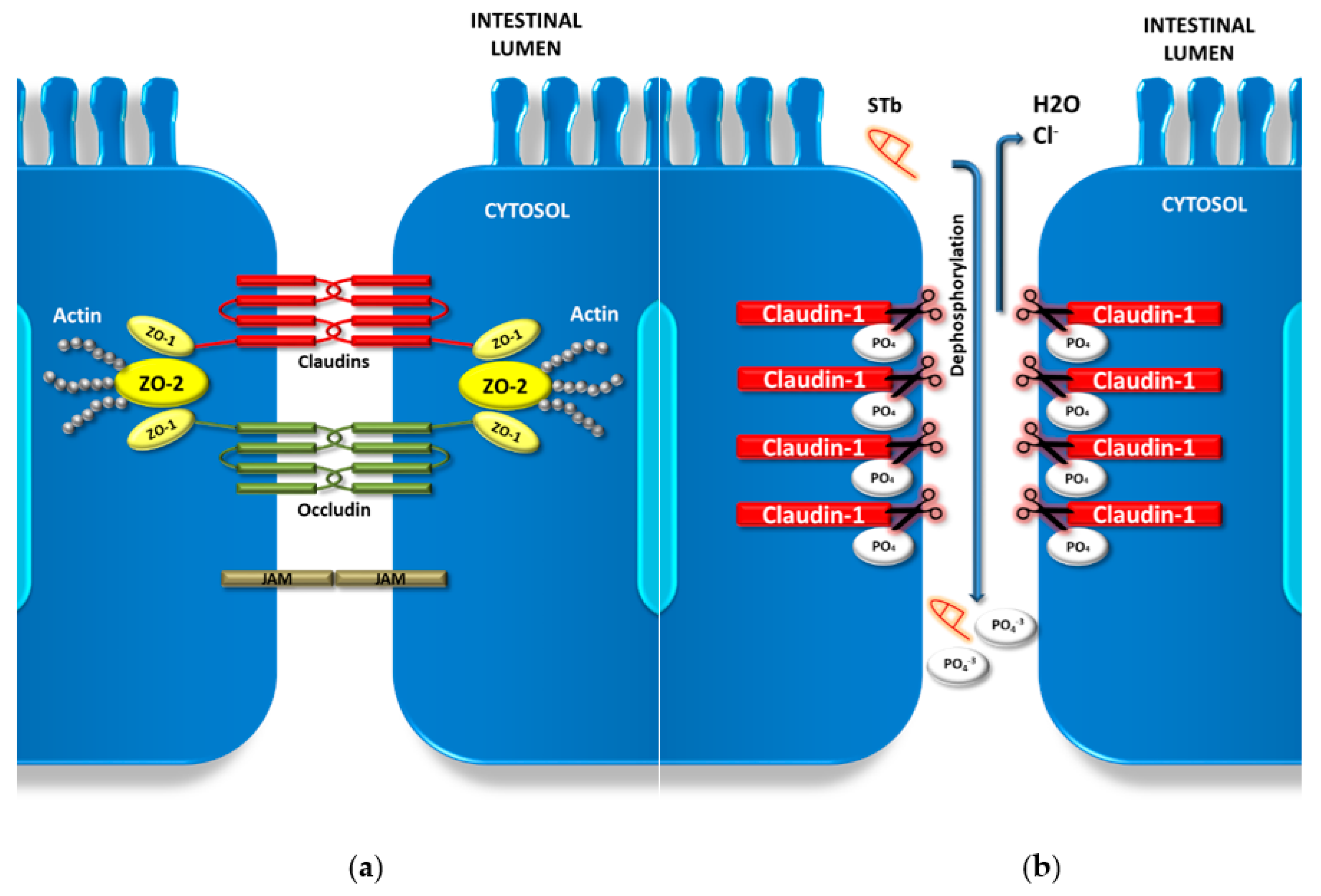
2.1. STb Alters Barrier Function through Tight Junction Manipulation
2.2. STb Pore Formation and Internalization by Enterocytes
2.3. STb Triggers Enterocyte Programmed Cell Death
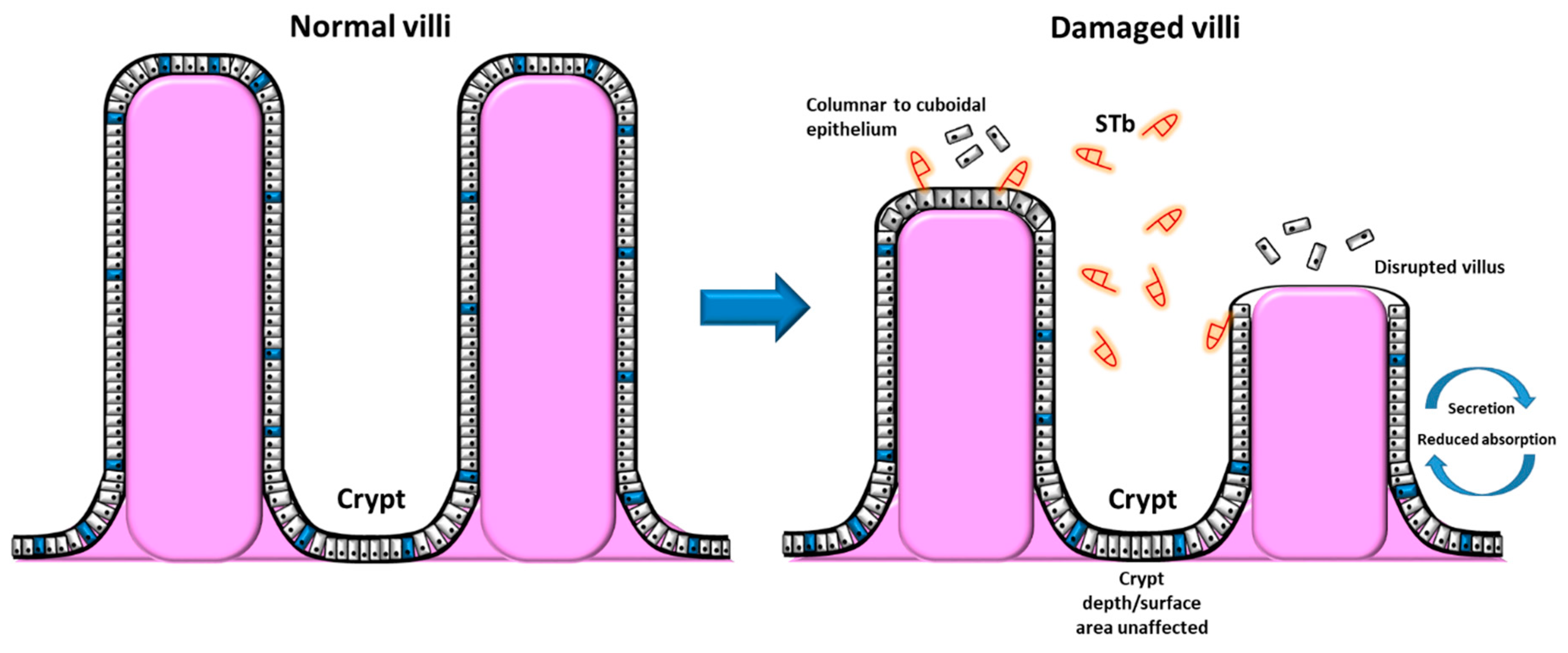
2.4. STb Causes Deterioration of the Intestinal Absorptive Mucosa
2.5. Fate of Enteroendocrine Cells and Gut Hormone Regulation
3. Conclusions
Funding
Conflicts of Interest
References
- Qadri, F.; Svennerholm, A.-M.; Faruque, A.S.G.; Sack, R.B. Enterotoxigenic Escherichia coli in Developing Countries: Epidemiology, Microbiology, Clinical Features, Treatment, and Prevention. Clin. Microbiol. Rev. 2005, 18, 465–483. [Google Scholar] [CrossRef]
- Dubreuil, J.D.; Isaacson, R.E.; Schifferli, D.M. Animal Enterotoxigenic Escherichia coli. EcoSal Plus 2016, 7. [Google Scholar] [CrossRef]
- WHO Future directions for research on enterotoxigenic Escherichia coli vaccines for developing countries. Wkly. Epidemiol. Rec. 2006, 81, 97–104.
- Ren, W.; Yin, J.; Duan, J.; Liu, G.; Zhu, X.; Chen, S.; Li, T.; Wang, S.; Tang, Y.; Hardwidge, P.R. Mouse intestinal innate immune responses altered by enterotoxigenic Escherichia coli (ETEC) infection. Microbes Infect. 2014, 16, 954–961. [Google Scholar] [CrossRef]
- Dubreuil, J.D. Escherichia coli STb toxin and colibacillosis: Knowing is half the battle. FEMS Microbiol. Lett. 2008, 278, 137–145. [Google Scholar] [CrossRef]
- Vipin Madhavan, T.P.; Sakellaris, H. Chapter Five—Colonization Factors of Enterotoxigenic Escherichia coli. In Advances in Applied Microbiology; Sariaslani, S., Gadd, G.M., Eds.; Academic Press: Cambridge, MA, USA, 2015; Volume 90, pp. 155–197. [Google Scholar]
- Ngendahayo Mukiza, C.; Dubreuil, J.D. Escherichia coli Heat-Stable Toxin b Impairs Intestinal Epithelial Barrier Function by Altering Tight Junction Proteins. Infect. Immun. 2013, 81, 2819–2827. [Google Scholar] [CrossRef]
- Spangler, B.D. Structure and function of cholera toxin and the related Escherichia coli heat-labile enterotoxin. Microbiol. Mol. Biol. Rev. 1992, 56, 622–647. [Google Scholar] [CrossRef]
- Syed, H.C.; Dubreuil, J.D. Escherichia coli STb toxin induces apoptosis in intestinal epithelial cell lines. Microb. Pathog. 2012, 53, 147–153. [Google Scholar] [CrossRef]
- Kupersztoch, Y.M.; Tachias, K.; Moomaw, C.R.; Dreyfus, L.A.; Urban, R.; Slaughter, C.; Whipp, S. Secretion of methanol-insoluble heat-stable enterotoxin (STB): Energy- and secA-dependent conversion of pre-STB to an intermediate indistinguishable from the extracellular toxin. J. Bacteriol. 1990, 172, 2427–2432. [Google Scholar] [CrossRef]
- Hitotsubashi, S.; Fujii, Y.; Yamanaka, H.; Okamoto, K. Some properties of purified Escherichia coli heat-stable enterotoxin II. Infect. Immun. 1992, 60, 4468–4474. [Google Scholar] [CrossRef]
- Dubreuil, J.D. Escherichia coli STb Enterotoxin: A Multifaceted Molecule. In Microbial Toxins; Stiles, B., Alape-Girón, A., Dubreuil, J.D., Mandal, M., Eds.; Toxinology; Springer: Dordrecht, The Netherlands, 2018; pp. 205–222. ISBN 978-94-007-6449-1. [Google Scholar]
- Dubreuil, J.D. Escherichia coli STb enterotoxin. Microbiology 1997, 143, 1783–1795. [Google Scholar] [CrossRef]
- Rousset, E.; Harel, J.; Dubreuil, D.J. Binding characteristics ofEscherichia colienterotoxin b (STb) to the pig jejunum and partial characterization of the molecule involved. Microb. Pathog. 1998, 24, 277–288. [Google Scholar] [CrossRef]
- Dreyfus, L.A.; Harville, B.; Howard, D.E.; Shaban, R.; Beatty, D.M.; Morris, S.J. Calcium influx mediated by the Escherichia coli heat-stable enterotoxin B (STB). Proc. Natl. Acad. Sci. USA 1993, 90, 3202–3206. [Google Scholar] [CrossRef]
- Fujii, Y.; Nomura, T.; Yamanaka, H.; Okamoto, K. Involvement of Ca2+ -Calmodulin-Dependent Protein Kinase II in the Intestinal Secretory Action of Escherichia coli Heat-Stable Enterotoxin II. Microbiol. Immunol. 1997, 41, 633–636. [Google Scholar] [CrossRef]
- Harville, B.A.; Dreyfus, L.A. Involvement of 5-hydroxytryptamine and prostaglandin E2 in the intestinal secretory action of Escherichia coli heat-stable enterotoxin B. Infect. Immun. 1995, 63, 745–750. [Google Scholar] [CrossRef]
- Berkes, J.; Viswanathan, V.K.; Savkovic, S.D.; Hecht, G. Intestinal epithelial responses to enteric pathogens: Effects on the tight junction barrier, ion transport, and inflammation. Gut 2003, 52, 439–451. [Google Scholar] [CrossRef]
- Odenwald, M.A.; Turner, J.R. The intestinal epithelial barrier: A therapeutic target? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 9–21. [Google Scholar] [CrossRef]
- Groschwitz, K.R.; Hogan, S.P. Intestinal barrier function: Molecular regulation and disease pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–20. [Google Scholar] [CrossRef]
- Laukoetter, M.G.; Bruewer, M.; Nusrat, A. Regulation of the intestinal epithelial barrier by the apical junctional complex. Curr. Opin. Gastroenterol. 2006, 22, 85–89. [Google Scholar] [CrossRef]
- Bauer, H.; Zweimueller-Mayer, J.; Steinbacher, P.; Lametschwandtner, A.; Bauer, H.C. The Dual Role of Zonula Occludens (ZO) Proteins. Available online: https://www.hindawi.com/journals/bmri/2010/402593/ (accessed on 28 May 2020).
- Balda, M.S.; Whitney, J.A.; Flores, C.; González, S.; Cereijido, M.; Matter, K. Functional dissociation of paracellular permeability and transepithelial electrical resistance and disruption of the apical-basolateral intramembrane diffusion barrier by expression of a mutant tight junction membrane protein. J. Cell Biol. 1996, 134, 1031–1049. [Google Scholar] [CrossRef]
- Nassour, H.; Dubreuil, J.D. Escherichia coli STb Enterotoxin Dislodges Claudin-1 from Epithelial Tight Junctions. PLoS ONE 2014, 9, e113273. [Google Scholar] [CrossRef]
- Pope, J.L.; Bhat, A.A.; Sharma, A.; Ahmad, R.; Krishnan, M.; Washington, M.K.; Beauchamp, R.D.; Singh, A.B.; Dhawan, P. Claudin-1 regulates intestinal epithelial homeostasis through the modulation of Notch-signalling. Gut 2014, 63, 622–634. [Google Scholar] [CrossRef]
- Raju, P.; Shashikanth, N.; Tsai, P.-Y.; Pongkorpsakol, P.; Chanez-Paredes, S.; Steinhagen, P.R.; Kuo, W.-T.; Singh, G.; Tsukita, S.; Turner, J.R. Inactivation of paracellular cation-selective claudin-2 channels attenuates immune-mediated experimental colitis in mice. J. Clin. Investig. 2020, 130, 5197–5208. [Google Scholar] [CrossRef]
- Nattramilarasu, P.K.; Bücker, R.; Lobo de Sá, F.D.; Fromm, A.; Nagel, O.; Lee, I.-F.M.; Butkevych, E.; Mousavi, S.; Genger, C.; Kløve, S.; et al. Campylobacter concisus Impairs Sodium Absorption in Colonic Epithelium via ENaC Dysfunction and Claudin-8 Disruption. Int. J. Mol. Sci. 2020, 21, 373. [Google Scholar] [CrossRef]
- Cameron, H.L.; Perdue, M.H. Muscarinic acetylcholine receptor activation increases transcellular transport of macromolecules across mouse and human intestinal epithelium in vitro. Neurogastroenterol. Motil. 2007, 19, 47–56. [Google Scholar] [CrossRef]
- Neal, M.D.; Leaphart, C.; Levy, R.; Prince, J.; Billiar, T.R.; Watkins, S.; Li, J.; Cetin, S.; Ford, H.; Schreiber, A.; et al. Enterocyte TLR4 Mediates Phagocytosis and Translocation of Bacteria Across the Intestinal Barrier. J. Immunol. 2006, 176, 3070–3079. [Google Scholar] [CrossRef]
- Van der Goot, F.G.; Lakey, J.; Pattus, F.; Kay, C.M.; Sorokine, O.; Van Dorsselaer, A.; Buckley, J.T. Spectroscopic study of the activation and oligomerization of the channel-forming toxin aerolysin: Identification of the site of proteolytic activation. Biochemistry 1992, 31, 8566–8570. [Google Scholar] [CrossRef]
- Labrie, V.; Harel, J.; Dubreuil, J.D. Oligomerization of Escherichia coli enterotoxin b through its C-terminal hydrophobic α-helix. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2001, 1535, 128–133. [Google Scholar] [CrossRef]
- Gonçalves, C.; Vachon, V.; Schwartz, J.-L.; Dubreuil, J.D. The Escherichia coli Enterotoxin STb Permeabilizes Piglet Jejunal Brush Border Membrane Vesicles. Infect. Immun. 2007, 75, 2208–2213. [Google Scholar] [CrossRef]
- Moniatte, M.; van der Goot, F.G.; Buckley, J.T.; Pattus, F.; van Dorsselaer, A. Characterisation of the heptameric pore-forming complex of the Aeromonas toxin aerolysin using MALDI-TOF mass spectrometry. FEBS Lett. 1996, 384, 269–272. [Google Scholar] [CrossRef]
- Olofsson, A.; Hebert, H.; Thelestam, M. The structure of Staphylococcus aureus α-toxin: Effects of trypsin treatment. J. Struct. Biol. 1991, 106, 199–204. [Google Scholar] [CrossRef]
- Labrie, V.; Harel, J.; Dubreuil, J.D. Escherichia coli heat-stable enterotoxin b (STb) in vivo internalization within rat intestinal epithelial cells. Vet. Res. 2002, 33, 223–228. [Google Scholar] [CrossRef]
- Gonçalves, C.; Dubreuil, J.D. Effect of Escherichia coli STb toxin on NIH-3T3 cells. FEMS Immunol. Med. Microbiol. 2009, 55, 432–441. [Google Scholar] [CrossRef]
- Escherichia coli STb Enterotoxin Toxicity and Internalization Investigations: A Mini-Review. Available online: https://scialert.net/abstract/?doi=jm.2007.209.215 (accessed on 17 May 2020).
- Fulda, S.; Debatin, K.-M. Extrinsic versus intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene 2006, 25, 4798–4811. [Google Scholar] [CrossRef]
- Iwanaga, T.; Han, H.; Adachi, K.; Fujita, T. A novel mechanism for disposing of effete epithelial cells in the small intestine of guinea pigs. Gastroenterology 1993, 105, 1089–1097. [Google Scholar] [CrossRef]
- Kim, J.M.; Eckmann, L.; Savidge, T.C.; Lowe, D.C.; Witthöft, T.; Kagnoff, M.F. Apoptosis of human intestinal epithelial cells after bacterial invasion. J. Clin. Investig. 1998, 102, 1815–1823. [Google Scholar] [CrossRef]
- Hall, P.A.; Coates, P.J.; Ansari, B.; Hopwood, D. Regulation of cell number in the mammalian gastrointestinal tract: The importance of apoptosis. J. Cell Sci. 1994, 107, 3569–3577. [Google Scholar]
- Degterev, A.; Boyce, M.; Yuan, J. A decade of caspases. Oncogene 2003, 22, 8543–8567. [Google Scholar] [CrossRef]
- Duprez, L.; Wirawan, E.; Berghe, T.V.; Vandenabeele, P. Major cell death pathways at a glance. Microbes Infect. 2009, 11, 1050–1062. [Google Scholar] [CrossRef]
- Xia, Y.; Bin, P.; Liu, S.; Chen, S.; Yin, J.; Liu, G.; Tang, Z.; Ren, W. Enterotoxigenic Escherichia coli infection promotes apoptosis in piglets. Microb. Pathog. 2018, 125, 290–294. [Google Scholar] [CrossRef]
- Wapnir, R.A.; Teichberg, S. Regulation mechanisms of intestinal secretion: Implications in nutrient absorption. J. Nutr. Biochem. 2002, 13, 190–199. [Google Scholar] [CrossRef]
- Powell, D.W. Ion and Water Transport in the Intestine. In Membrane Transport Processes in Organized Systems; Andreoli, T.E., Hoffman, J.F., Fanestil, D.D., Schultz, S.G., Eds.; Springer: Boston, MA, USA, 1987; pp. 175–212. ISBN 978-1-4684-5404-8. [Google Scholar]
- Pappenheimer, J.R.; Reiss, K.Z. Contribution of solvent drag through intercellular junctions to absorption of nutrients by the small intestine of the rat. J. Membr. Biol. 1987, 100, 123–136. [Google Scholar] [CrossRef]
- Kiela, P.R.; Ghishan, F.K. Physiology of Intestinal Absorption and Secretion. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 145–159. [Google Scholar] [CrossRef]
- Whipp, S.C.; Moon, H.W.; Kemeny, L.J.; Argenzio, R.A. Effect of virus-induced destruction of villous epithelium on intestinal secretion induced by heat-stable Escherichia coli enterotoxins and prostaglandin E1 in swine. Am. J. Vet. Res. 1985, 46, 637–642. [Google Scholar]
- Whipp, S.C.; Moseley, S.L.; Moon, H.W. Microscopic alterations in jejunal epithelium of 3-week-old pigs induced by pig-specific, mouse-negative, heat-stable Escherichia coli enterotoxin. Am. J. Vet. Res. 1986, 47, 615–618. [Google Scholar]
- Whipp, S.C.; Kokue, E.; Morgan, R.W.; Rose, R.; Moon, H.W. Functional significance of histologic alterations induced by Escherichiacoli pig-specific, mouse-negative, heat-stable enterotoxin (STb). Vet. Res. Commun. 1987, 11, 41–55. [Google Scholar] [CrossRef]
- Sternini, C.; Anselmi, L.; Rozengurt, E. Enteroendocrine cells: A site of ‘taste’ in gastrointestinal chemosensing. Curr. Opin. Endocrinol. Diabetes Obes. 2008, 15, 73–78. [Google Scholar] [CrossRef]
- The New Biology of Gastrointestinal Hormones|Physiological Reviews. Available online: https://journals.physiology.org/doi/full/10.1152/physrev.1998.78.4.1087 (accessed on 20 May 2020).
- Baggio, L.L.; Drucker, D.J. Biology of Incretins: GLP-1 and GIP. Gastroenterology 2007, 132, 2131–2157. [Google Scholar] [CrossRef]
- Gagnon, J.; Brubaker, P.L. NCI-H716 Cells. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 221–228. ISBN 978-3-319-16104-4. [Google Scholar]
- Röder, P.V.; Wu, B.; Liu, Y.; Han, W. Pancreatic regulation of glucose homeostasis. Exp. Mol. Med. 2016, 48, e219. [Google Scholar] [CrossRef]
- Suzuki, K.; Jayasena, C.N.; Bloom, S.R. The Gut Hormones in Appetite Regulation. J. Obes. 2011, 2011. [Google Scholar] [CrossRef]
- Guo, C.; Huang, T.; Chen, A.; Chen, X.; Wang, L.; Shen, F.; Gu, X. Glucagon-like peptide 1 improves insulin resistance in vitro through anti-inflammation of macrophages. Braz. J. Med. Biol. Res. 2016, 49. [Google Scholar] [CrossRef]
- Amato, A.; Baldassano, S.; Mulè, F. GLP2: An underestimated signal for improving glycaemic control and insulin sensitivity. J. Endocrinol. 2016, 229, R57–R66. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cameron, H.L.; Perdue, M.H. Stress Impairs Murine Intestinal Barrier Function: Improvement by Glucagon-Like Peptide-2. J. Pharmacol. Exp. Ther. 2005, 314, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Cameron, H.L.; Yang, P.-C.; Perdue, M.H. Glucagon-like peptide-2-enhanced barrier function reduces pathophysiology in a model of food allergy. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 284, G905–G912. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Jia, G.; Jiang, Y.; Deng, Q.; Chen, Z.; Xu, Z.; Chen, X.; Wang, K. Effect of Glucagon-like Peptide 2 on Tight Junction in Jejunal Epithelium of Weaned Pigs though MAPK Signaling Pathway. Asian Australas. J. Anim. Sci. 2014, 27, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Kochar, B.; Long, M.D.; Shelton, E.; Young, L.; Farraye, F.A.; Yajnik, V.; Herfarth, H. Safety and Efficacy of Teduglutide (Gattex) in patients with Crohn’s disease and need for parenteral support due to short bowel syndrome associated intestinal failure. J. Clin. Gastroenterol. 2017, 51, 508–511. [Google Scholar] [CrossRef]
- Jeppesen, P.B.; Sanguinetti, E.L.; Buchman, A.; Howard, L.; Scolapio, J.S.; Ziegler, T.R.; Gregory, J.; Tappenden, K.A.; Holst, J.; Mortensen, P.B. Teduglutide (ALX-0600), a dipeptidyl peptidase IV resistant glucagon-like peptide 2 analogue, improves intestinal function in short bowel syndrome patients. Gut 2005, 54, 1224–1231. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Butt, S.; Saleh, M.; Gagnon, J. Impact of the Escherichia coli Heat-Stable Enterotoxin b (STb) on Gut Health and Function. Toxins 2020, 12, 760. https://doi.org/10.3390/toxins12120760
Butt S, Saleh M, Gagnon J. Impact of the Escherichia coli Heat-Stable Enterotoxin b (STb) on Gut Health and Function. Toxins. 2020; 12(12):760. https://doi.org/10.3390/toxins12120760
Chicago/Turabian StyleButt, Shahnawaz, Mazen Saleh, and Jeffrey Gagnon. 2020. "Impact of the Escherichia coli Heat-Stable Enterotoxin b (STb) on Gut Health and Function" Toxins 12, no. 12: 760. https://doi.org/10.3390/toxins12120760
APA StyleButt, S., Saleh, M., & Gagnon, J. (2020). Impact of the Escherichia coli Heat-Stable Enterotoxin b (STb) on Gut Health and Function. Toxins, 12(12), 760. https://doi.org/10.3390/toxins12120760