Host–Receptor Post-Translational Modifications Refine Staphylococcal Leukocidin Cytotoxicity

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
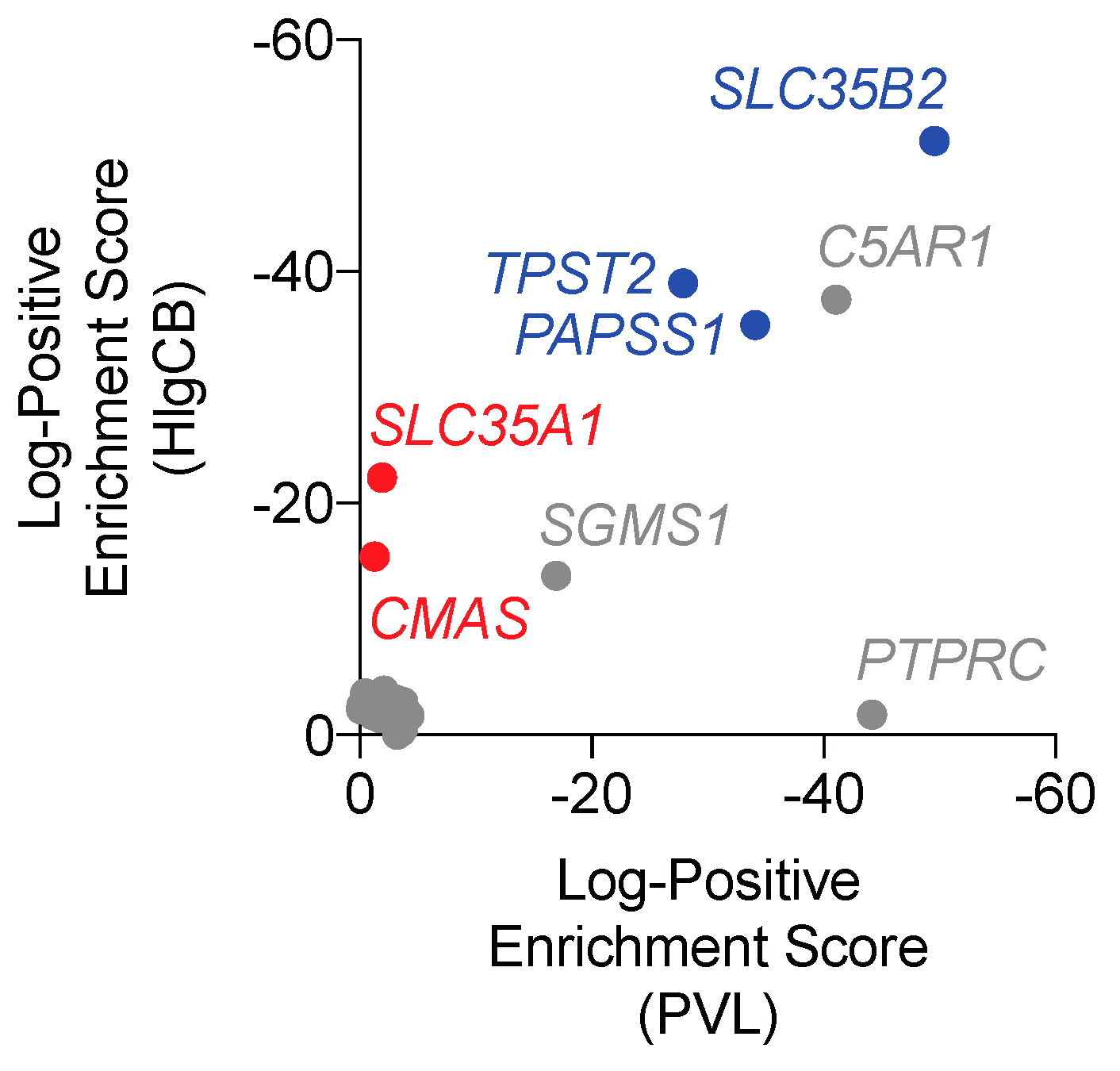
2.1. PTM Pathways Affect Susceptibility to PVL and HlgCB Cytotoxicity.
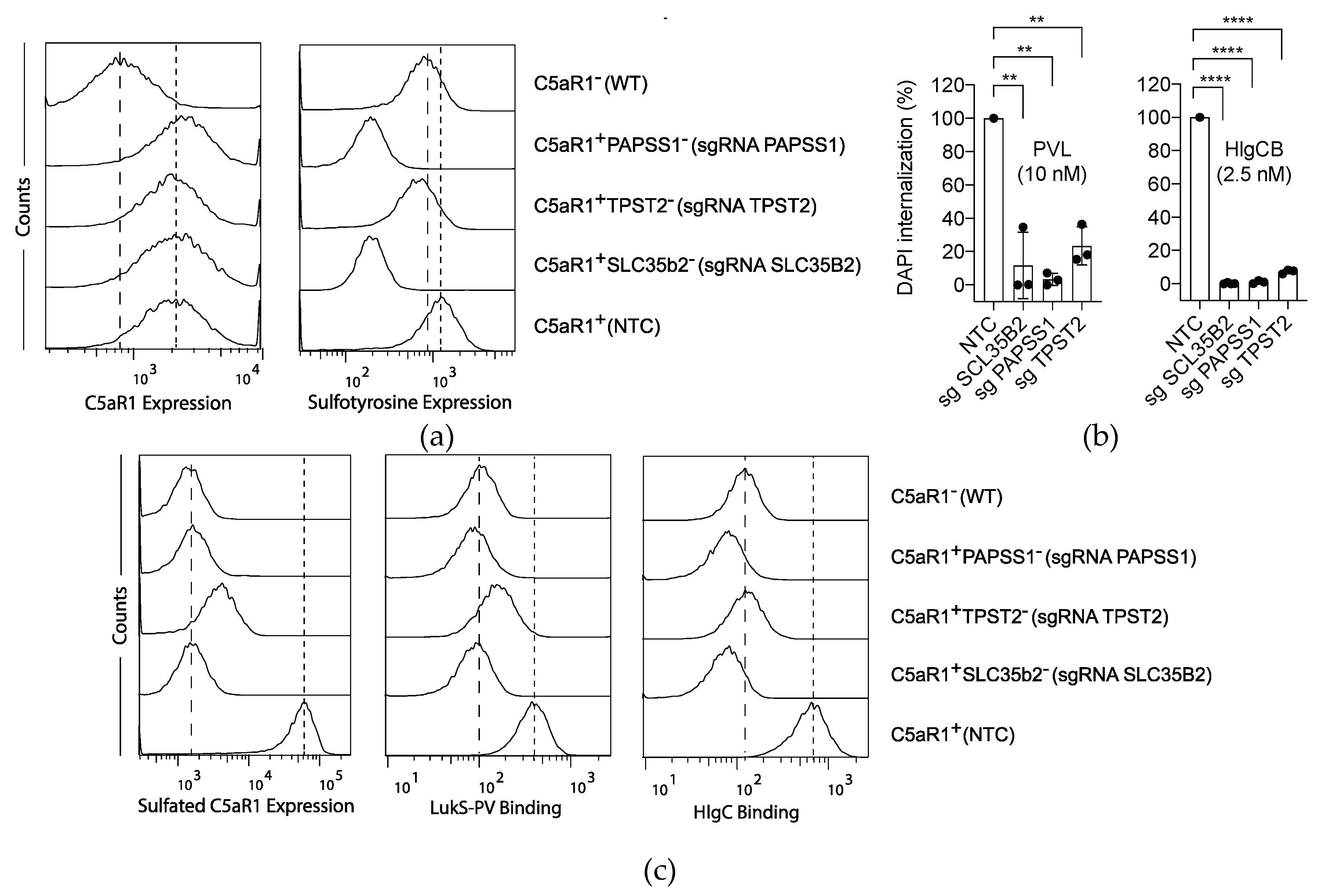
2.2. Sulfation of C5aR1 Facilitates both PVL and HlgCB Cytotoxicity.
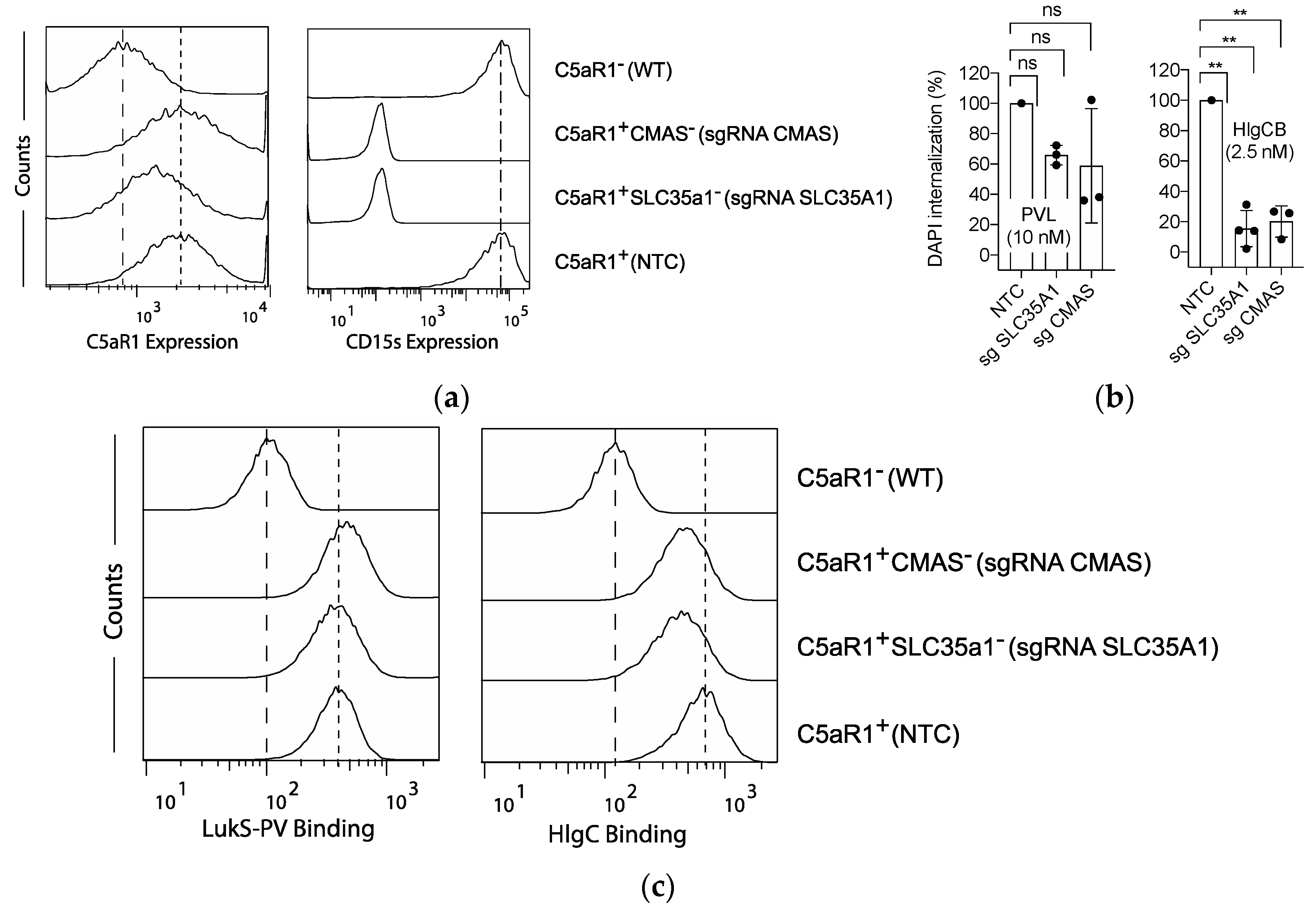
2.3. The role of C5aR1 Sialylation in HlgCB- and PVL-Induced Cytotoxicity.
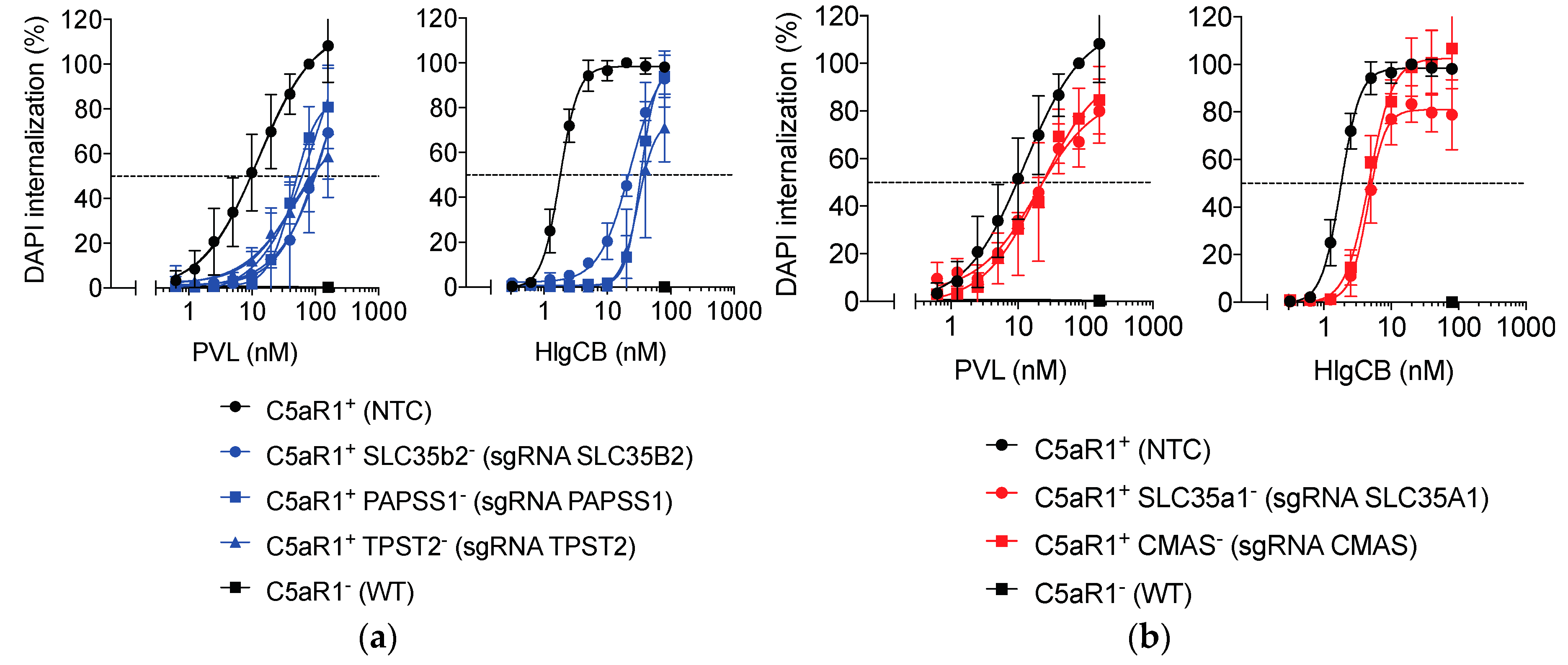
2.4. Sulfation and to a Lesser Extent Sialylation Refine Susceptibility to PVL and HlgCB.
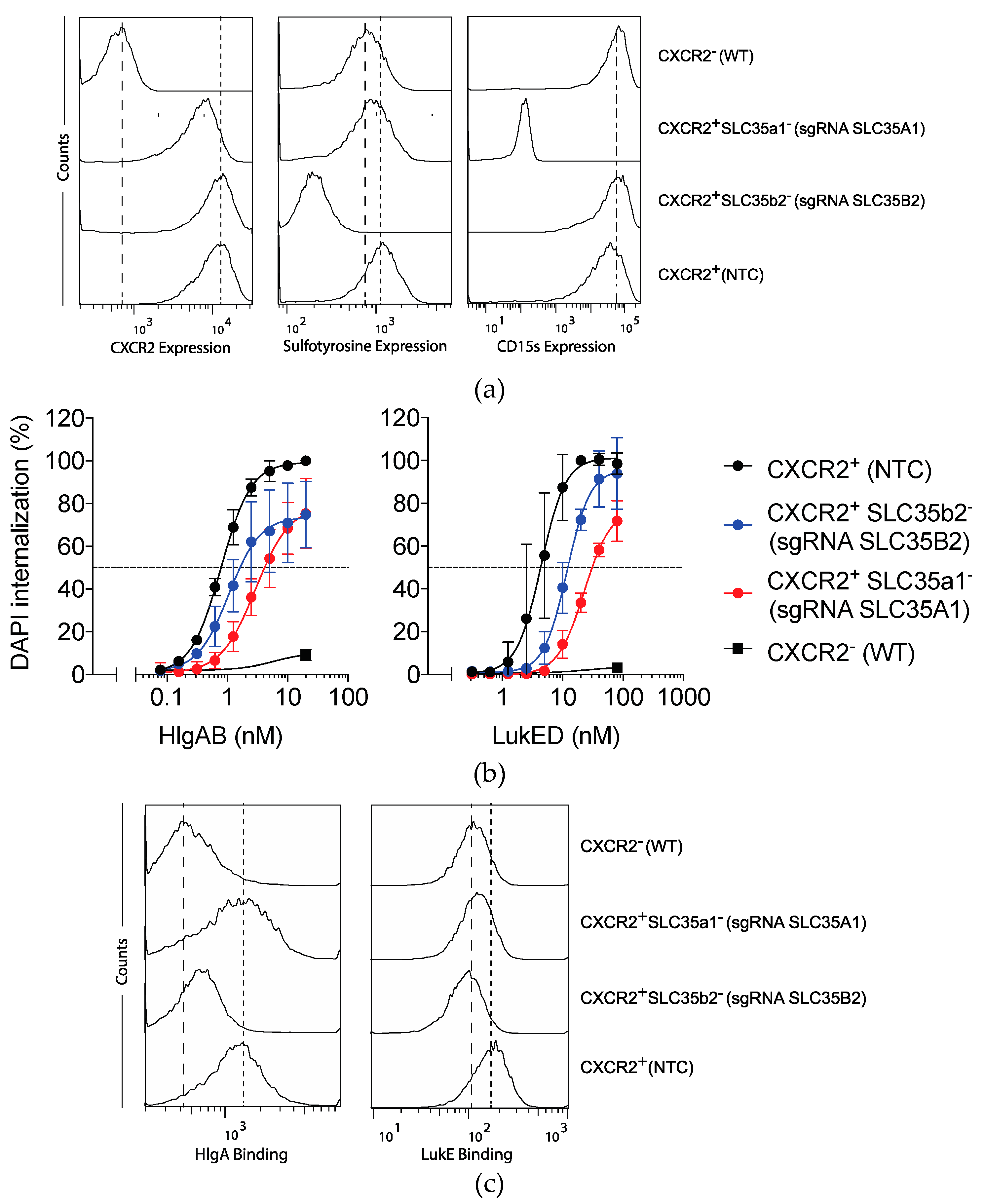
2.5. Sialyation and to a Lesser Extent Sulfation Refine Susceptibility to LukED and HlgAB.
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Constructs
4.2. Genome-Wide CRISPR/Cas9 Library Screen in U937-C5aR1 Cells
4.3. Recombinant Protein Production and Cell Permeability Assays
4.4. Determination of Receptor Expression Levels and Binding Assays
4.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Thwaites, G.E.; Edgeworth, J.D.; Gkrania-Klotsas, E.; Kirby, A.; Tilley, R.; Torok, M.E.; Walker, S.; Wertheim, H.F.; Wilson, P.; Llewelyn, M.J.; et al. Clinical management of Staphylococcus aureus bacteraemia. Lancet Infect. Dis. 2011, 11, 208–222. [Google Scholar] [CrossRef]
- Beutler, B. Innate immunity: An overview. Mol. Immunol. 2004, 40, 845–859. [Google Scholar] [CrossRef] [PubMed]
- Spaan, A.N.; Surewaard, B.G.; Nijland, R.; van Strijp, J.A. Neutrophils Versus Staphylococcus aureus: A Biological Tug of War. Annu. Rev. Microbiol. 2013, 67, 629–650. [Google Scholar] [CrossRef] [Green Version]
- Spaan, A.N.; van Strijp, J.A.G.; Torres, V.J. Leukocidins: Staphylococcal bi-component pore-forming toxins find their receptors. Nat. Rev. Microbiol. 2017, 15, 435. [Google Scholar] [CrossRef]
- Alonzo, F., 3rd; Torres, V.J. The Bicomponent Pore-Forming Leucocidins of Staphylococcus aureus. Microbiol. Mol. Biol. Rev. 2014, 78, 199–230. [Google Scholar] [CrossRef] [Green Version]
- Spaan, A.N.; Henry, T.; van Rooijen, W.J.; Perret, M.; Badiou, C.; Aerts, P.C.; Kemmink, J.; de Haas, C.J.; van Kessel, K.P.; Vandenesch, F.; et al. The staphylococcal toxin Panton-Valentine Leukocidin targets human C5a receptors. Cell Host Microbe 2013, 13, 584–594. [Google Scholar] [CrossRef] [Green Version]
- Spaan, A.N.; Vrieling, M.; Wallet, P.; Badiou, C.; Reyes-Robles, T.; Ohneck, E.A.; Benito, Y.; de Haas, C.J.; Day, C.J.; Jennings, M.P.; et al. The staphylococcal toxins gamma-haemolysin AB and CB differentially target phagocytes by employing specific chemokine receptors. Nat. Commun. 2014, 5, 5438. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Robles, T.; Alonzo, F., 3rd; Kozhaya, L.; Lacy, D.B.; Unutmaz, D.; Torres, V.J. Staphylococcus aureus Leukotoxin ED Targets the Chemokine Receptors CXCR1 and CXCR2 to Kill Leukocytes and Promote Infection. Cell Host Microbe 2013, 14, 453–459. [Google Scholar] [CrossRef] [Green Version]
- Alonzo, F., 3rd; Kozhaya, L.; Rawlings, S.A.; Reyes-Robles, T.; DuMont, A.L.; Myszka, D.G.; Landau, N.R.; Unutmaz, D.; Torres, V.J. CCR5 is a receptor for Staphylococcus aureus leukotoxin ED. Nature 2013, 493, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Spaan, A.N.; Reyes-Robles, T.; Badiou, C.; Cochet, S.; Boguslawski, K.M.; Yoong, P.; Day, C.J.; de Haas, C.J.; van Kessel, K.P.; Vandenesch, F.; et al. Staphylococcus aureus Targets the Duffy Antigen Receptor for Chemokines (DARC) to Lyse Erythrocytes. Cell Host Microbe 2015, 18, 363–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tromp, A.T.; Van Gent, M.; Abrial, P.; Martin, A.; Jansen, J.P.; De Haas, C.J.C.; Van Kessel, K.P.M.; Bardoel, B.W.; Kruse, E.; Bourdonnay, E.; et al. Human CD45 is an F-component-specific receptor for the staphylococcal toxin Panton-Valentine leukocidin. Nat. Microbiol. 2018, 3, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Virreira Winter, S.; Zychlinsky, A.; Bardoel, B.W. Genome-wide CRISPR screen reveals novel host factors required for Staphylococcus aureus alpha-hemolysin-mediated toxicity. Sci. Rep. 2016, 6, 24242. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.S.; Wang, C.C.; Chen, B.H.; Hou, Y.H.; Hung, K.S.; Mao, Y.C. Tyrosine sulfation as a protein post-translational modification. Molecules 2015, 20, 2138–2164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhide, G.P.; Colley, K.J. Sialylation of N-glycans: Mechanism, cellular compartmentalization and function. Histochem. Cell Biol. 2017, 147, 149–174. [Google Scholar] [CrossRef] [PubMed]
- Spaan, A.N.; Schiepers, A.; de Haas, C.J.; van Hooijdonk, D.D.; Badiou, C.; Contamin, H.; Vandenesch, F.; Lina, G.; Gerard, N.P.; Gerard, C.; et al. Differential Interaction of the Staphylococcal Toxins Panton-Valentine Leukocidin and gamma-Hemolysin CB with Human C5a Receptors. J. Immunol. 2015, 195, 1034–1043. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.T.; Garneau-Tsodikova, S.; Gatto, G.J., Jr. Protein posttranslational modifications: The chemistry of proteome diversifications. Angew. Chem. Int. Ed. Engl. 2005, 44, 7342–7372. [Google Scholar] [CrossRef]
- Farzan, M.; Schnitzler, C.E.; Vasilieva, N.; Leung, D.; Kuhn, J.; Gerard, C.; Gerard, N.P.; Choe, H. Sulfated tyrosines contribute to the formation of the C5a docking site of the human C5a anaphylatoxin receptor. J. Exp. Med. 2001, 193, 1059–1066. [Google Scholar] [CrossRef]
- Ludeman, J.P.; Stone, M.J. The structural role of receptor tyrosine sulfation in chemokine recognition. Br. J. Pharmacol. 2014, 171, 1167–1179. [Google Scholar] [CrossRef] [Green Version]
- Park, R.J.; Wang, T.; Koundakjian, D.; Hultquist, J.F.; Lamothe-Molina, P.; Monel, B.; Schumann, K.; Yu, H.; Krupzcak, K.M.; Garcia-Beltran, W.; et al. A genome-wide CRISPR screen identifies a restricted set of HIV host dependency factors. Nat. Genet. 2017, 49, 193–203. [Google Scholar] [CrossRef]
- Choe, H.; Moore, M.J.; Owens, C.M.; Wright, P.L.; Vasilieva, N.; Li, W.; Singh, A.P.; Shakri, R.; Chitnis, C.E.; Farzan, M. Sulphated tyrosines mediate association of chemokines and Plasmodium vivax Duffy binding protein with the Duffy antigen/receptor for chemokines (DARC). Mol. Microbiol. 2005, 55, 1413–1422. [Google Scholar] [CrossRef] [PubMed]
- Poole, J.; Day, C.J.; von Itzstein, M.; Paton, J.C.; Jennings, M.P. Glycointeractions in bacterial pathogenesis. Nat. Rev. Microbiol. 2018, 16, 440–452. [Google Scholar] [CrossRef] [PubMed]
- Noda, M.; Kato, I.; Hirayama, T.; Matsuda, F. Fixation and inactivation of staphylococcal leukocidin by phosphatidylcholine and ganglioside GM1 in rabbit polymorphonuclear leukocytes. Infect. Immun. 1980, 29, 678–684. [Google Scholar] [PubMed]
- Moussouras, N.A.; Getschman, A.E.; Lackner, E.R.; Veldkamp, C.T.; Dwinell, M.B.; Volkman, B.F. Differences in Sulfotyrosine Binding amongst CXCR1 and CXCR2 Chemokine Ligands. Int. J. Mol. Sci. 2017, 18, 1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann-Fatu, C.; Bayer, P. Determinants of tyrosylprotein sulfation coding and substrate specificity of tyrosylprotein sulfotransferases in metazoans. Chem. Biol. Interact. 2016, 259, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Westmuckett, A.D.; Hoffhines, A.J.; Borghei, A.; Moore, K.L. Early postnatal pulmonary failure and primary hypothyroidism in mice with combined TPST-1 and TPST-2 deficiency. Gen. Comp. Endocrinol. 2008, 156, 145–153. [Google Scholar] [CrossRef] [Green Version]
- Nicholls, J.M.; Moss, R.B.; Haslam, S.M. The use of sialidase therapy for respiratory viral infections. Antivir. Res. 2013, 98, 401–409. [Google Scholar] [CrossRef]
- Macauley, M.S.; Arlian, B.M.; Rillahan, C.D.; Pang, P.C.; Bortell, N.; Marcondes, M.C.; Haslam, S.M.; Dell, A.; Paulson, J.C. Systemic blockade of sialylation in mice with a global inhibitor of sialyltransferases. J. Biol. Chem. 2014, 289, 35149–35158. [Google Scholar] [CrossRef] [Green Version]
- Mishiro, E.; Sakakibara, Y.; Liu, M.C.; Suiko, M. Differential enzymatic characteristics and tissue-specific expression of human TPST-1 and TPST-2. J. Biochem. 2006, 140, 731–737. [Google Scholar] [CrossRef]
- Gillet, Y.; Issartel, B.; Vanhems, P.; Fournet, J.C.; Lina, G.; Bes, M.; Vandenesch, F.; Piemont, Y.; Brousse, N.; Floret, D.; et al. Association between Staphylococcus aureus strains carrying gene for Panton-Valentine leukocidin and highly lethal necrotising pneumonia in young immunocompetent patients. Lancet 2002, 359, 753–759. [Google Scholar] [CrossRef]
- Alcais, A.; Abel, L.; Casanova, J.L. Human genetics of infectious diseases: Between proof of principle and paradigm. J. Clin. Investig. 2009, 119, 2506–2514. [Google Scholar] [CrossRef] [PubMed]
- Casanova, J.L. Human genetic basis of interindividual variability in the course of infection. Proc. Natl. Acad. Sci. USA 2015, 112, E7118–E7127. [Google Scholar] [CrossRef] [Green Version]
- Deleo, F.R.; Otto, M.; Kreiswirth, B.N.; Chambers, H.F. Community-associated meticillin-resistant Staphylococcus aureus. Lancet 2010, 375, 1557–1568. [Google Scholar] [CrossRef] [Green Version]
- Niemann, S.; Ehrhardt, C.; Medina, E.; Warnking, K.; Tuchscherr, L.; Heitmann, V.; Ludwig, S.; Peters, G.; Loffler, B. Combined Action of Influenza Virus and Staphylococcus aureus Panton-Valentine Leukocidin Provokes Severe Lung Epithelium Damage. J. Infect. Dis. 2012, 206, 1138–1148. [Google Scholar] [CrossRef] [PubMed]
- DeLeo, F.R.; Otto, M. An antidote for Staphylococcus aureus pneumonia? J. Exp. Med. 2008, 205, 271–274. [Google Scholar] [CrossRef] [Green Version]
- Van de Weijer, M.L.; Bassik, M.C.; Luteijn, R.D.; Voorburg, C.M.; Lohuis, M.A.; Kremmer, E.; Hoeben, R.C.; LeProust, E.M.; Chen, S.; Hoelen, H.; et al. A high-coverage shRNA screen identifies TMEM129 as an E3 ligase involved in ER-associated protein degradation. Nat. Commun. 2014, 5, 3832. [Google Scholar] [CrossRef] [Green Version]
- Perret, M.; Badiou, C.; Lina, G.; Burbaud, S.; Benito, Y.; Bes, M.; Cottin, V.; Couzon, F.; Juruj, C.; Dauwalder, O.; et al. Cross-talk between S. aureus leukocidins-intoxicated macrophages and lung epithelial cells triggers chemokine secretion in an inflammasome-dependent manner. Cell. Microbiol. 2012, 14, 1019–1036. [Google Scholar] [CrossRef]
- Haapasalo, K.; Wollman, A.J.M.; de Haas, C.J.C.; van Kessel, K.P.M.; van Strijp, J.A.G.; Leake, M.C. Staphylococcus aureus toxin LukSF dissociates from its membrane receptor target to enable renewed ligand sequestration. FASEB J. 2019, 33, 3807–3824. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| sgRNA | PVL | HlgCB | ||||
|---|---|---|---|---|---|---|
| EC50 (nM) | 95% Conf. Interval (nM) | Fold Increased EC50 | EC50 (nM) | 95% Conf. Interval (nM) | Fold Increased EC50 | |
| NTC | 12.9 | 9.1–21.5 | 1.8 | 1.7–1.9 | ||
| SLC35B2 | 123.1 | 38.1–infin | 9.5 | 30.3 | 23.1–50.6 | 16.8 |
| PAPSS1 | 44.6 | 35.7–63.4 | 3.5 | 33.3 | 30.6–36.3 | 18.5 |
| TPST2 | 42.5 | 22.2–infin | 3.3 | 23.1 | 18.6–34.3 | 12.8 |
| SLC35A1 | 18.5 | 11.5–67.4 | 1.4 | 4.5 | 3.9–5.1 | 2.5 |
| CMAS | 19.4 | 11.3–79.6 | 1.5 | 4.7 | 4.0–5.6 | 2.6 |
| sgRNA | HlgAB | LukED | ||||
|---|---|---|---|---|---|---|
| EC50 (nM) | 95% Conf. Interval (nM) | Fold Increased EC50 | EC50 (nM) | 95% Conf. Interval (nM) | Fold Increased EC50 | |
| NTC | 0.8 | 0.7–0.9 | 4.4 | 3.4–5.7 | ||
| SLC35B2 | 1.1 | 0.7–1.7 | 1.4 | 11.8 | 10.1–14.0 | 2.7 |
| SLC35A1 | 2.9 | 2.1–5.1 | 3.6 | 22.7 | 19.8–27.3 | 5.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tromp, A.T.; Van Gent, M.; Jansen, J.P.; Scheepmaker, L.M.; Velthuizen, A.; De Haas, C.J.C.; Van Kessel, K.P.M.; Bardoel, B.W.; Boettcher, M.; McManus, M.T.; et al. Host–Receptor Post-Translational Modifications Refine Staphylococcal Leukocidin Cytotoxicity. Toxins 2020, 12, 106. https://doi.org/10.3390/toxins12020106
Tromp AT, Van Gent M, Jansen JP, Scheepmaker LM, Velthuizen A, De Haas CJC, Van Kessel KPM, Bardoel BW, Boettcher M, McManus MT, et al. Host–Receptor Post-Translational Modifications Refine Staphylococcal Leukocidin Cytotoxicity. Toxins. 2020; 12(2):106. https://doi.org/10.3390/toxins12020106
Chicago/Turabian StyleTromp, Angelino T., Michiel Van Gent, Joris P. Jansen, Lisette M. Scheepmaker, Anneroos Velthuizen, Carla J.C. De Haas, Kok P.M. Van Kessel, Bart W. Bardoel, Michael Boettcher, Michael T. McManus, and et al. 2020. "Host–Receptor Post-Translational Modifications Refine Staphylococcal Leukocidin Cytotoxicity" Toxins 12, no. 2: 106. https://doi.org/10.3390/toxins12020106
APA StyleTromp, A. T., Van Gent, M., Jansen, J. P., Scheepmaker, L. M., Velthuizen, A., De Haas, C. J. C., Van Kessel, K. P. M., Bardoel, B. W., Boettcher, M., McManus, M. T., Van Strijp, J. A. G., Lebbink, R. J., Haas, P.-J. A., & Spaan, A. N. (2020). Host–Receptor Post-Translational Modifications Refine Staphylococcal Leukocidin Cytotoxicity. Toxins, 12(2), 106. https://doi.org/10.3390/toxins12020106