1. Introduction
With a production of about 4.4 million tons and a cultivated area of 1.3 million hectares in 2015, Italy is one of the most important countries in the world for durum wheat (
Triticum turgidum subsp.
durum (Desf.) Husn.) cultivation [
1,
2]. In fact, durum wheat is the main crop in many regions of the peninsula [
2] and in the last decades, its cultivation expanded from the “typical” Southern Italian areas to Central and Northern parts of the country. For example, in Umbria (Central Italy) and in Emilia Romagna (Northern Italy), durum wheat production doubled during the 2005–2015 period [
2]. In detail, in 2015 about 65% of durum wheat national production was obtained in Southern regions, almost 23% in Central regions, and the remaining 12% in Northern regions [
2]. Italian durum wheat is mainly used for pasta production. In 2014, Italy was the country with the highest pasta production and consumption worldwide, with about 3.4 million tons and 25 kg per capita, respectively [
3]. For these reasons, durum wheat plays a key role in the Italian and European agri-food compartment. Therefore, the maintenance of a high qualitative standard is an essential aspect of its cultivation across the country. However, several fungal microorganisms, being able to infect/contaminate durum wheat kernels in the field, represent a serious threat to grain production and quality. Additionally, the aforementioned expansion of durum wheat cultivation from Southern to Northern Italian areas has significantly increased this risk, due to more favorable climatic conditions to diseases [
4,
5]. The fungal community colonizing durum wheat grains mainly consists of mycotoxigenic genera, principally
Alternaria and
Fusarium [
6,
7,
8,
9]. Most of the species belonging to these genera are able to biosynthesize mycotoxins, secondary fungal metabolites having strong acute and chronic adverse effects on animals and humans [
10]. Thus, the presence of toxigenic mycobiota in durum wheat kernels represents a non-negligible risk factor for consumers’ health [
11] because of the classification of some fungal secondary metabolites as carcinogens or possible carcinogens by the International Agency for Research on Cancer (IARC). Therefore, the European Union (EU) has established maximum or recommended levels for several mycotoxins in various food matrixes, such as raw durum wheat and some derived products such as pasta [
12,
13]. For other compounds, the European Food Safety Authority (EFSA) performed risk assessments and gave scientific opinions [
14,
15].
Among the various fungal microorganisms, those belonging to the genus
Fusarium are particularly dangerous. They are the causal agents of
Fusarium head blight (FHB) a disease capable of strongly impairing not only crop yield but also its quality. In fact, FHB is caused by many distinct species belonging to multiple
Fusarium species complexes [
16] that, in addition to being able to infect and damage wheat heads in the field, may biosynthesize a wide range of toxic secondary metabolites. Since durum wheat is more susceptible to FHB than soft wheat (
Triticum aestivum L.) [
17,
18],
Fusarium infections and mycotoxin contaminations are of particular concern in Italy [
19]. Moreover, FHB pathogens negatively affect durum wheat quality as they may damage the protein fraction of the kernels [
20,
21]. Among FHB causal agents,
Fusarium graminearum is globally considered as the most important, because of its widespread incidence and aggressiveness [
22] as well as because it is the main producer of type B trichothecenes such as deoxynivalenol (DON) and nivalenol (NIV) [
23]. A similar toxigenic profile also characterizes
Fusarium culmorum, an important FHB agent in Southern Mediterranean areas [
24]. In recent years, other
Fusarium species such as
Fusarium poae and
Fusarium avenaceum increased their importance in the FHB community in many wheat-cultivation areas worldwide [
25,
26,
27,
28,
29,
30]. In particular, in certain Italian regions and cultivation seasons, these two species were found to be the most prominent members of the FHB complex of wheat, even more present than
F. graminearum [
7,
8,
18,
31,
32,
33].
F. avenaceum is mainly able to biosynthesize depsipetides, such as enniatin analogues (ENs) and moniliformin (MON) [
34,
35].
F. poae, in addition to NIV, produces depsipeptides such as beauvericin (BEA) [
34,
35]. Type A trichothecenes, such as T-2 and HT-2 toxins [
35], are mainly produced by
Fusarium sporotrichioides and
Fusarium langsethiae [
36,
37]. The isolation of these last two species from the grains has been demonstrated to be difficult [
36,
38,
39] and their presence might consequently be often underestimated. Other
Fusarium species may also be detected with a low incidence in wheat grains. FHB complexity is exacerbated by the fact that, in addition to well-known mycotoxins, a wide range of other secondary metabolites produced by
Fusarium species and having an unknown impact on health, might also potentially occur in the kernels [
7,
40,
41].
The FHB complex composition as well as the type and quantity of mycotoxins in the grain, strongly depend on climatic factors such as temperature, humidity and rainfall, especially those occurring during crop anthesis [
42,
43,
44,
45]. Thus, seasonal patterns usually show a strong influence on the incidence of the different
Fusarium species and of their mycotoxins in the grains [
46,
47]. In addition, agronomic factors such as crop rotations, crop management systems [
27,
47,
48,
49], deployed varieties [
50] and the use of fungicides [
51] may affect FHB complex composition and the presence of secondary metabolites. One or more of these factors might potentially be the reason for regional differences within the FHB complex as well as for the presence of the mycotoxins associated with wheat grains in different world cultivation areas [
30,
52,
53,
54,
55,
56]. For example, in Italy, FHB incidence and DON occurrence usually increase from Southern to Northern regions [
18,
19].
Given this diversity, a better understanding of the regional composition of the FHB complex as well as of mycotoxin occurrence are critical to understanding the effective suitability of a certain geographic area for producing high-quality durum wheat and for setting up effective disease and mycotoxin integrated management strategies in different cultivation areas [
52].
Therefore, in the present study, durum wheat kernels harvested in three climatically different Italian regions (Emilia Romagna, Umbria and Sardinia), representative of three different cultivation areas (North, Center and South, respectively) were analyzed to: (1) determine the mycobiota infecting the grains with two different isolation methods (potato dextrose agar, PDA and deep-freezing blotter, DFB); (2) molecularly determine FHB complex composition; (3) quantify the main Fusarium species in the grains by quantitative real-time polymerase chain reaction (qPCR); (4) detect a wide range of fungal secondary metabolites in the grains by liquid chromatography tandem mass spectrometry (LC-MS/MS). The main objective of this investigation was to determine the importance of different cultivation areas in affecting the presence of fungal communities, including FHB causal agents, as well as of secondary metabolites in Italian durum wheat grains.
2. Results
2.1. Mycobiota Composition
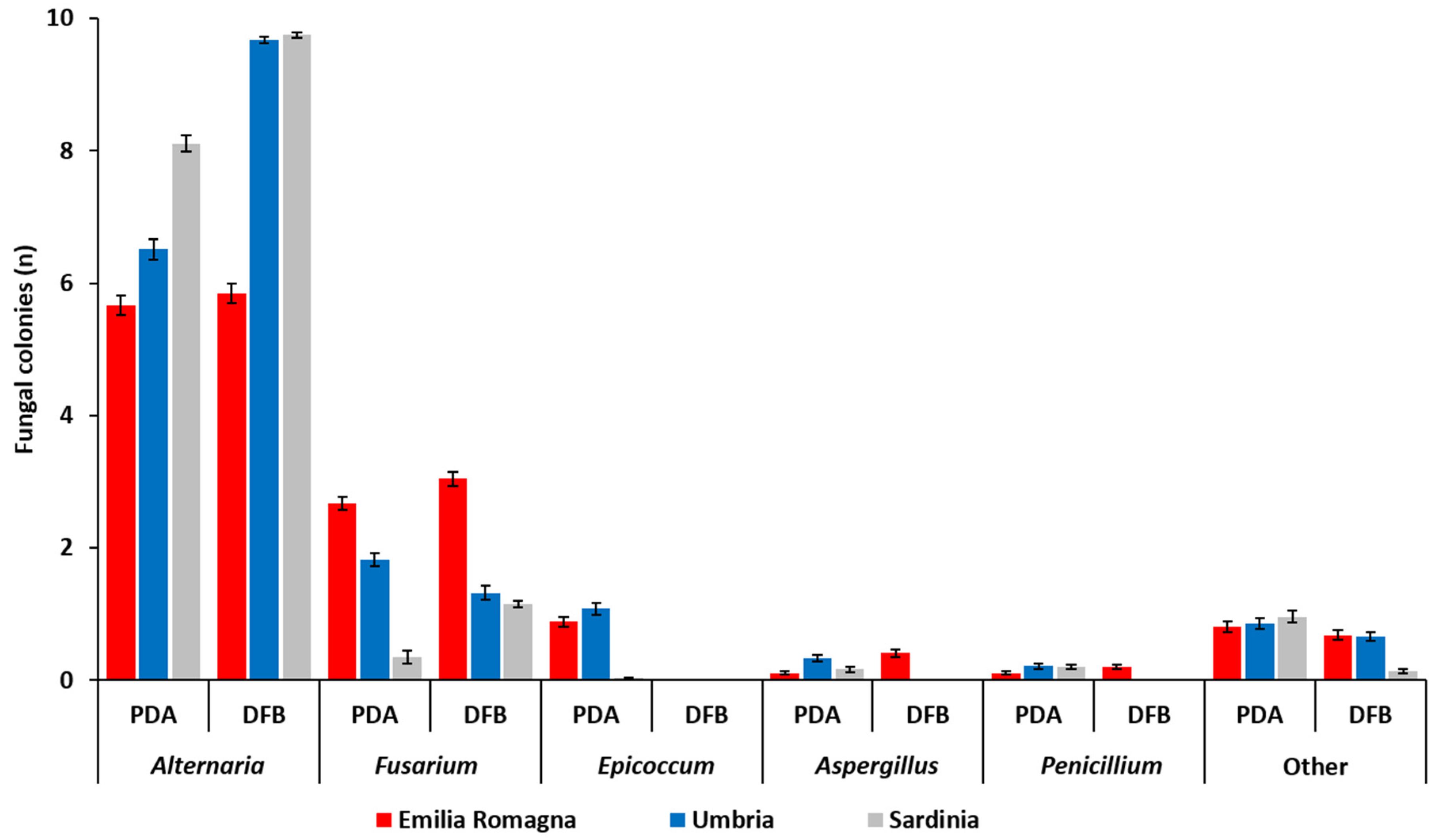
The number (
n) of fungal colonies belonging to the genera
Alternaria,
Fusarium,
Epicoccum,
Aspergillus and
Penicillium developing after 5 (PDA) or 7 (DFB) days of incubation from grains sampled in each of the three Italian regions is shown in
Figure 1 and detailed in
Table S1. Fungi that were not included in the aforementioned genera were classified as “other” (
Figure 1,
Table S1).
The fungal colonies belonging to the genus
Alternaria showed a significantly higher presence (
p ≤ 1 × 10
−4) within the fungal community developed from durum wheat grains following both PDA and DFB isolations in all the surveyed regions. In addition, with the exception of samples from Emilia Romagna (
p = 0.41), the number of
Alternaria colonies recovered with the DFB was significantly higher (
p ≤ 1 × 10
−4) than those detected on PDA (
Figure 1,
Table S1).
Alternaria presence in the Emilia Romagna samples was significantly lower (
p ≤ 1 × 10
−4) than that recovered in the Sardinian and in the Umbrian ones with the DFB and on PDA. Conversely, in the Sardinian samples, the isolates belonging to this genus were more frequent than in the Umbrian ones, even if only on PDA (
p = 1 × 10
−4) and not with the DFB (
p = 0.34) (
Figure 1,
Table S1).
The genus
Fusarium was the second component of the durum wheat grain mycobiota following both DFB (
p ≤ 1 × 10
−4) and PDA (
p ≤ 0.04) isolations in all surveyed areas. Significant differences between the two isolation methods were recorded in Sardinia, where
Fusarium colonies were significantly higher with the DFB than on PDA (
p = 1 × 10
−4) as well as in Umbria, where
Fusarium colonies grown on PDA were significantly higher than those isolated with the DFB (
p = 0.002) (
Figure 1,
Table S1). In Emilia Romagna, the presence of
Fusarium colonies developing from the grains on PDA was significantly higher than that of the Umbrian samples (
p = 1 × 10
−4), in which they were significantly higher than in the Sardinian ones (
p = 1 × 10
−4). Similar results were also observed following DFB isolations (
p ≤ 1 × 10
−4), even if in this case Umbria and Sardinia did not significantly differ from each other (
p = 0.24) in terms of isolated
Fusarium colonies.
In addition to the two most frequently recovered fungal genera (
Alternaria and
Fusarium), the mycobiota of durum wheat grains harvested in the three investigated Italian regions was also composed of other fungal genera, such as
Epicoccum,
Aspergillus and
Penicillium. The colonies belonging to the genus
Epicoccum developed only on PDA while they were not isolated with the other method (
Figure 1). In addition, in Emilia Romagna and Umbria the incidence of
Epicoccum spp. on PDA was significantly higher than that recorded in Sardinia (
p ≤ 1 × 10
−4). With the exception of Emilia Romagna samples, also colonies of
Aspergillus and
Penicillium were obtained only on PDA and not with the DFB. On PDA, the presence of
Aspergillus was higher in the Umbrian samples (
p ≤ 0.01). On the same medium, no significant differences between different regions (
p ≥ 0.07) in terms of number of developed
Penicillium colonies were found.
2.2. Fusarium Head Blight (FHB) Complex Composition
With regard to the total
Fusarium isolates developed from durum wheat with the two isolation methods and successively identified by partial
translation elongation factor 1α (
tef1α) region sequencing, significant differences between cultivation regions were detected. In detail, the average number of total
Fusarium isolates infecting the grains followed the statistically significant gradient: Emilia Romagna >> Umbria > Sardinia. This was observed both for PDA (11.8, 4.7, 0.74, respectively) (
p ≤ 0.002) and for the DFB method (15.2, 6.10, 1.96, respectively) (
p ≤ 0.01). Even if in the three surveyed regions the number of
Fusarium isolates recovered with the DFB was higher than that obtained on PDA, no significant differences between the two methods in any of the investigated areas (
p ≥ 0.1) were found. The number (
n) of
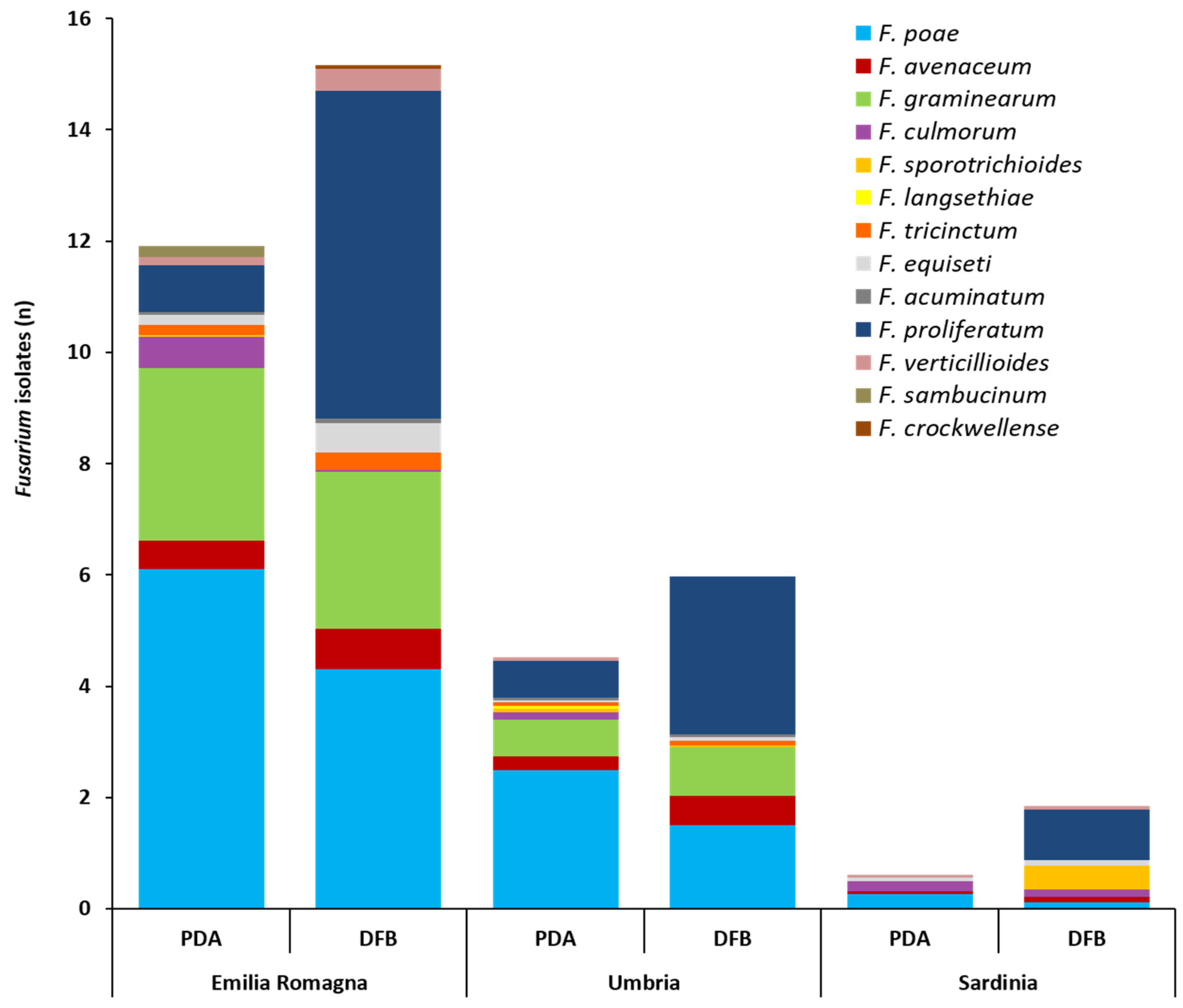
Fusarium isolates ascribable to the different species is shown in
Figure 2 and detailed in
Table S2.
The
Fusarium community isolated on PDA and with the DFB method from the kernels collected across Emilia Romagna was composed of 11 different species (
Figure 2,
Table S2). On PDA,
F. poae was the most frequent species (6.1) (
p ≤ 0.001) followed by
F. graminearum (3.0) (
p ≤ 7 × 10
−4). Eight more species (
Fusarium proliferatum,
F. culmorum,
F. avenaceum,
Fusarium equiseti,
Fusarium sambucinum,
Fusarium tricinctum,
Fusarium verticillioides and
Fusarium acuminatum) (
Figure 2,
Table S2) were isolated from the kernels on PDA, showing a significantly lower presence than
F. poae and
F. graminearum (
p ≤ 7 × 10
−4) (
Table S2). With DFB,
F. proliferatum (5.98) and
F. poae (4.33) showed the significantly highest presence (
p ≤ 1 × 10
−4). These species were followed also in this case by
F. graminearum (2.82), which was significantly lower than
F. proliferatum (
p = 7 × 10
−4) but not than
F. poae (
p = 0.07). The other six species (
F. avenaceum,
F. equiseti,
F. verticillioides,
F. tricinctum,
F. acuminatum and
Fusarium crockwellense) were significantly lower than the previous three species (
p ≤ 2 × 10
−4). Differences in recovering
Fusarium species between the two isolation methods in terms of number of
Fusarium isolates were recorded only for
F. culmorum and
F. proliferatum. In detail,
F. culmorum showed a significantly higher presence on PDA (0.56) than with DFB (nd) (
p = 0.03), while,
F. proliferatum showed a significantly higher presence with DFB (5.98) than on PDA (0.84) (
p = 1 × 10
−4).
The
Fusarium community isolated on PDA and with DFB from durum wheat kernels collected in Umbria was composed of 11 different species (
Figure 2,
Table S2). On PDA,
F. poae showed a significantly higher presence (2.53) (
p ≤ 0.001) followed by
F. proliferatum (0.67) and
F. graminearum (0.66), which showed a non-significant difference (
p = 0.97) with each other. The other seven
Fusarium species (
F. avenaceum,
F. culmorum,
F. langsethiae,
F. sporotrichioides,
F. tricinctum,
F. acuminatum and
F. verticillioides) isolated on PDA showed a similar presence (
p ≥ 0.29) and only
F. aveanceum and
F. culmorum were not significantly different from
F. proliferatum and
F. graminearum (
p ≥ 0.06). With DFB,
F. proliferatum (2.82) was the most isolated species even if it was not significantly different from
F. poae (1.55) (
p = 0.06). These two species were followed by
F. graminearum (0.87), which was significantly lower than
F. proliferatum (
p = 0.001) only. The other four
Fusarium species (
F. avenaceum,
F. tricinctum,
F. equiseti and
F. acuminatum) showed a similar presence.
F. avenaceum (0.53) was the only species not being significantly different from
F. graminearum (
p = 0.37). In addition, in this case, only
F. proliferatum showed a significantly higher presence with the DFB (2.82) than on PDA (0.67) (
p = 4 × 10
−4).
The
Fusarium community isolated on PDA and with the DFB method from durum wheat kernels collected in Sardinia was composed of seven different species (
Figure 2,
Table S2). On PDA, no significant differences were recorded among the five different species isolated (
F. poae,
F. culmorum,
F. graminearum,
F. proliferatum,
F. avenaceum) (
p ≥ 0.24). With DFB,
F. proliferatum showed a significantly higher presence (0.92) (
p ≤ 0.02) than the other five species (
F. culmorum,
F. poae,
F. avenaceum,
F. equiseti,
F. verticillioides), with the exception of
F. sporotrichioides (0.47) (
p = 0.27). Differences between the two isolation methods in terms of number of
Fusarium isolates were recorded not only for
F. proliferatum but also for
F. sporotrichioides. Both species showed a significantly higher presence with DFB than on PDA (
p = 0.005 and 0.04, respectively).
Significant differences (
p ≤ 0.04) between cultivation regions in terms of single species presence (
Table S2) were also detected. In detail,
F. poae followed the significant gradient Emilia Romagna > Umbria > Sardinia (both for PDA and DFB);
F. graminearum followed the significant gradient Emilia Romagna > Umbria >> Sardinia (absent) (both for PDA and DFB);
F. avenaceum followed the significant gradient Emilia Romagna ≥ Umbria ≥ Sardinia (only for PDA);
F. proliferatum followed the significant gradient Emilia Romagna ≥ Umbria > Sardinia (both for PDA and DFB).
2.3. Fungal Biomass Accumulation
R2 values calculated from the linear equations of the six standard curves were 0.99 for all Fusarium species, with the exception of F. avenaceum whose R2 value was 0.97. Reaction efficiencies obtained from the linear equations of the six standard curves were 1.97 for F. graminearum and F. avenaceum, 1.94 for F. poae, 1.98 for F. langsethiae, 1.99 for F. culmorum and 1.96 for F. sporotrichioides. Dissociation curve analysis showed specific amplification products in the presence of pure fungal DNA (standard curves) and in the presence of DNA of the six investigated Fusarium species (samples). No target amplification was detected in negative controls. For these reasons, cycle threshold (Ct) values used to quantify fungal biomass were those for which dissociation curve analysis showed the presence of specific amplification products.
The fungal biomass of six
Fusarium species (pg of fungal DNA/ng durum wheat grains DNA) as quantified by qPCR directly in durum wheat grains collected in three different Italian regions (Emilia Romagna, Umbria, Sardinia) is shown in
Figure 3 and detailed in
Table S3.
In the durum wheat grains harvested in Emilia Romagna, all six analyzed species were detected. F. poae was present in all Emilia Romagna samples and F. graminearum and F. avenaceum were both detected in 9 out of 10 Emilia Romagna samples. In addition, F. langsethiae, F. culmorum and F. sporotrichioides were present in 4, 3 and 1 Emilia Romagna samples, respectively. F. poae showed the highest fungal biomass levels, followed by F. graminearum and F. avenaceum even if no significant differences between these three species were recorded (p ≥ 0.20). Similarly, F. langsethiae, F. culmorum and F. sporotrichioides were not significantly different from each other (p ≥ 0.15), but they showed significantly lower fungal biomass levels than the previous ones (p ≤ 0.03).
In the durum wheat grains harvested in the Umbria region, all the species analyzed by qPCR, with the exception of F. sporotrichioides, were detected. F. poae, present in all the analyzed samples (n = 10), showed a significantly higher biomass level (p ≤ 0.01) than those of the other four species. No significant differences (p ≥ 0.16) between F. avenaceum, F. graminearum, F. langsethiae and F. culmorum fungal biomasses were detected. These species were present in 4 (F. avenaceum and F. langsethiae), 2 (F. graminearum) and 1 (F. culmorum) out of 10 durum wheat grains samples, respectively.
In the durum wheat grains collected across Sardinia, 4 out of 6 analyzed species were detected. F. poae was present in 6 out of 10 samples. F. culmorum and F. langsethiae were both present in 2 samples, while, F. avenaceum was present in only one sample. No significant differences (p ≥ 0.10) in fungal biomass accumulation for these four species were detected in the Sardinian samples. No F. graminearum and F. sporotrichioides were detected in the durum wheat grains collected in this area.
Significant differences between cultivation areas in terms of fungal accumulation in the grains (
Figure 3 and
Table S3) were also detected for
F. poae and
F. avenaceum. In detail,
F. poae followed the significant gradient (
p ≤ 0.04) Emilia Romagna >> Umbria > Sardinia;
F. avenaceum followed the significant gradient (
p ≤ 0.001) Emilia Romagna > Umbria = Sardinia. For
F. graminearum and
F. langsethiae, although their presence was higher in the Emilia Romagna region, no significant differences were recorded between the different surveyed areas (
p = 0.125 and
p ≥ 0.07, respectively).
Finally, considering the total Fusarium fungal biomass (calculated as the sum of the fungal biomasses of each of the six Fusarium species analyzed) detected in durum wheat grains by qPCR, significant differences between cultivation areas were detected. In detail, the total Fusarium fungal biomass followed the significant gradient (p ≤ 0.04) Emilia Romagna >> Umbria > Sardinia.
2.4. Fungal Secondary Metabolites Accumulation
The fungal secondary metabolites (μg/kg) as quantified by LC/MS-MS in the durum wheat grains collected in the three Italian regions are summarized by category in
Table 1 and detailed within each group in
Table 2 (trichothecenes),
Table 3 (zearalenone and fumonisins),
Table 4 (depsipeptides),
Table 5 (other
Fusarium secondary metabolites),
Table 6 (
Alternaria secondary metabolites) and
Table 7 (
Claviceps secondary metabolites).
In general, samples collected in Emilia Romagna were particularly contaminated by
Fusarium secondary metabolites (
Table 1). Total trichothecenes followed the significant gradient (
p ≤ 0.05) Emilia Romagna >> Umbria > Sardinia (
Table 1). Total depsipetides followed the significant gradient (
p ≤ 0.03) Emilia Romagna > Umbria = Sardinia (
Table 1). The other
Fusarium secondary metabolites followed the significant gradient (
p ≤ 0.04): Emilia Romagna > Umbria > Sardinia (
Table 1). The other two classes of compounds biosynthesized by
Fusarium, zearalenone and fumonisins, were present in a low percentage of samples only in Emilia Romagna (
Table 1 and
Table 3). Conversely,
Alternaria secondary metabolites were particularly present in durum wheat grains harvested in Umbria (
Table 1). In detail, total
Alternaria secondary metabolites accumulation followed the significant gradient (
p ≤ 0.04): Umbria > Emilia Romagna > Sardinia. Finally, regarding
Claviceps metabolites, even if they were mainly present in the Umbrian samples, no significant differences (
p ≥ 0.26) between the three analyzed regions were detected, being only a few samples positive to these compounds (
Table 1 and
Table 7).
Focusing on trichothecenes (
Table 2), LC-MS/MS analysis showed that DON, culmorin, 15-hydroxyculmorin, 15-hydroxyculmoron and 5-hydroxyculmorin were particularly present in durum wheat grains harvested in Emilia Romagna. In particular, DON followed the significant gradient (
p ≤ 0.02) Emilia Romagna >> Umbria = Sardinia. A similar gradient (Emilia Romagna > Umbria = Sardinia) was also observed for culmorin and 15-hydroxyculmorin (
p ≤ 0.04 and 0.03, respectively). The secondary metabolites 15-hydroxyculmoron and 5-hydroxyculmorin were present only in samples harvested in Emilia Romagna. Other trichothecenes, present in all surveyed regions were NIV, HT-2 toxin, HT-2 glucoside and T-2 toxin. These compounds were present at higher levels in Emilia Romagna samples, however, no significant differences were detected between Emilia Romagna and Umbria samples. In fact, they followed the significant gradient (
p ≤ 0.05) Emilia Romagna = Umbria > Sardinia. Finally, samples positive to the presence of deacetylneosolaniol, diacetoxyscirpenol, T-2 tetraol and monoacetoxyscirpenol were found only in the Emilia Romagna and Umbria samples and, despite the higher levels of these compounds recovered in Emilia Romagna, no significant differences were detected between regions (
p ≥ 0.18).
With regard to depsipeptides (
Table 4), the EN analogues analyzed in this study (enniatin A, ENA; enniatin A1, ENA1; enniatin B, ENB; enniatin B1, ENB1; enniatin B2, ENB2; enniatin B3, ENB3) and BEA were all particularly present in Emilia Romagna samples followed by Umbria and Sardinia samples. In general, EN analogues amounts were higher than BEA in all the analyzed regions. The EN analogues accumulated in durum wheat grains harvested in Emilia Romagna and Umbria following the gradient ENB1 > ENB > ENA1 > ENB2 > ENA > ENB3. In durum wheat harvested in Sardinia, EN analogues followed the gradient ENB1 > ENB > ENB2 > ENB3. Significant differences between cultivation areas for ENB, ENB1 and ENB2 were recorded. All these three analogues were significantly higher in the Emilia Romagna samples (
p ≤ 0.05), while, despite the higher levels detected in the Umbrian samples, no significant differences (
p ≥ 0.12) were recorded between Umbrian and Sardinian ones. The analogues ENA, ENA1 and ENB3 were present only in Emilia Romagna and Umbria without (ENA and ENA1) or with (ENB3) significant differences between these two cultivation areas (
p = 0.27,
p = 0.20 and
p = 0.02, respectively). With respect to BEA, the accumulation of this secondary metabolite followed a significant gradient (
p ≤ 0.03) Emilia Romagna > Umbria > Sardinia.
Regarding other
Fusarium secondary metabolites (
Table 5), they were all present in the grains harvested in Emilia Romagna with the exception of epiequisetin that was present in the Umbrian grains. In detail, several compounds (antibiotic Y, chlamidospordiol, fusarin C, fusarinolic acid, sambucinol) were detected only in the Emilia Romagna grains. Some others (aminodimethyloctadecanol, aurofusarin, butenolide, chalmydosporol, epiequisetin and equisetin) were only present in the Emilia Romagna and Umbrian grains with significant differences recorded only for aurofusarin (
p = 0.02). The remaining
Fusarium metabolites (MON, apicidin and chrysogin) were detected in the grains from all surveyed regions, with significant differences found for MON (
p ≤ 0.02, Emilia Romagna > Umbria = Sardinia) and chrysogin (
p ≤ 0.003, Emilia Romagna > Umbria = Sardinia).
With respect to
Alternaria secondary metabolites (
Table 6), several compounds (tenuazonic acid, tentoxin, pyrenophorol, macrosporin) were notably higher in the Umbrian grains, while some others (altersetin, alternariol, infectopyrone and alternariol methyl ether) were notably higher in the Emilia Romagna samples. However, no significant differences were recorded between Umbria and Emilia Romagna for the content of these single compounds. Conversely, significant differences between these two regions and Sardinia were detected for tenuazonic acid, alternariol methyl ether and infectopyrone (Umbria = Emilia Romagna > Sardinia,
p ≤ 0.04).
Finally, legislated mycotoxins did not exceed the maximum or recommended levels [
12,
13]. However, two samples harvested in Emilia Romagna showed DON levels (1740 and 1710 μg/kg) close to the maximum permitted for durum wheat grains (1750 μg/kg) and one sample collected in the same region showed T2+HT2 toxins level (97.4 μg/kg) close to the recommended concentration for durum wheat (100 μg/kg).
2.5. Correlation between Fungal Biomass and Fungal Secondary Metabolites in the Grains
With regard to F. poae, a typical BEA and NIV producer, its biomass was positively related to the amounts of BEA in the kernels, both for the Emilia Romagna and Umbrian samples, with correlation coefficients respectively of 0.72 (p = 0.018; n = 10) and 0.81 (p = 0.0048; n = 10). Likewise, F. poae biomass was positively correlated with the amounts of NIV in the kernels, only for the Umbrian samples (correlation coefficient = 0.79; p = 0.0071; n = 10)). Similarly, for F. avenaceum, one of the main ENs and MON producers, its biomass was positively related to the amounts of ENs (calculated by the sum of all six analyzed analogues) as well as with MON in the kernels of the Umbrian samples with correlation coefficients of 0.92 (p = 2 × 10−4; n = 10) and 0.93 (p = 1 × 10−4, n = 10), respectively. With respect to F. graminearum, a DON producer, its biomass was positively related to DON accumulation in the Emilia Romagna and Umbrian grains with correlation coefficients of 0.80 (p = 0.05, n = 10) and 0.95 (p = 1 × 10−4, n = 10), respectively. Finally, the sum of F. sporotrichioides and F. langsethiae biomasses, two of the main T-2 and HT-2 toxin producers, were positively correlated with the content of these two mycotoxins in the samples collected in Emilia Romagna and Umbria, with correlation coefficients of 0.95 (p = 1 × 10−4, n = 10) and 0.87 (p = 0.01, n = 10), respectively.
2.6. Weather Conditions in the Three Surveyed Regions
Weather data (rainfall, soil humidity and temperature) collected in the three different Italian regions (Emilia Romagna, Umbria and Sardinia) during the anthesis phase of durum wheat are shown in
Figure 4. The highest rainfall levels (sum and daily average) during the considered 30-day period for each of the three regions, were recorded in Emilia Romagna (75.3 mm and 2.5 mm), followed by Umbria (27.7 mm and 0.9 mm) and then Sardinia (17.2 mm and 0.5 mm). In particular, in Emilia Romagna, heavy rainfalls occurred between 20 and 27 May. Differently, in the Umbria region, significant rain events were recorded between 26 and 30 April. Finally, in Sardinia, significant rain events occurred only on 27 and 28 April, indicating that in this region the anthesis phase was characterized by particularly dry conditions. Similarly, the highest soil humidity average level was detected in Emilia Romagna with a peak of 66.9% on the day after (23 May) the main rain event (22 May) which occurred in this region during crop anthesis.
3. Discussion
Climate is an important driver of different aspects of fungal biogeography, including global distribution of common fungi as well as composition and diversity of fungal communities [
57]. Thus, this paper reports the results of a study about fungal communities and their secondary metabolites in durum wheat grains harvested in 2015 in three Italian regions (Emilia Romagna, Umbria and Sardinia) characterized by three different climatic conditions.
The mycobiota composition of durum wheat grains was investigated with two of the most common methods used for the isolation of seed-borne fungal pathogens worldwide (PDA and DFB) [
58]. With regard to the global presence of
Fusarium species, no differences between the two methods were detected, while, in these experimental conditions, the DFB method seemed to favor the development of microorganisms belonging to the genus
Alternaria, probably due to the absence of grain external disinfection. Conversely, PDA appeared to favor the development of
Epicoccum,
Aspergillus and
Penicillium species.
Species of the genus
Alternaria were the main components of durum wheat mycobiota in all the investigated regions, showing their ubiquity across Italian cultivation areas. In Umbria, the prevalence of this genus in the durum wheat fungal community has been already highlighted [
7]. Similarly, the high abundance of
Alternaria species in wheat grains was previously detected also in other countries worldwide [
9,
55,
59,
60,
61,
62,
63,
64]. In fact,
Alternaria species could be associated with wheat grains as saprophytes, causing black (sooty) head mold [
65], or as pathogens, causing black point of kernels [
66]. In addition, these species are potentially able to biosynthesize mycotoxins [
6].
In this study, different incidence levels of this genus and of its related secondary metabolites in the three cultivation areas were detected, suggesting that environmental conditions may have influenced the presence of this fungal genus and of its mycotoxins. In particular, durum wheat samples harvested in Umbria (Central Italy) showed the highest levels of
Alternaria species as well as, in general, of their secondary metabolites. Some
Alternaria mycotoxins have been previously reported in durum wheat grains harvested in the Umbria region [
40], however this study, expanding the number of compounds analyzed, revealed that tenuazonic acid, alternariol, alternariol methyl ether, altersetin, tentoxin, infectopyrone, macrosporin and pyrenophorol were widespread in this cultivation area. The presence of all or some of the analyzed
Alternaria secondary metabolites in Emilia Romagna and Sardinia, even if with different levels, revealed the remarkable diffusion of these compounds in Italian durum wheat. Further studies will be necessary to identify the
Alternaria species associated with durum wheat grains as well as to better understand the toxicity of
Alternaria mycotoxins. In fact, despite their presence in cereals has been documented for years, no regulation has been applied so far worldwide, apart from the limits for tenuazonic acid recently established by the Bavarian Health and Food Safety Authority in infant food in Germany [
6].
Fusarium species were the second most prevalent components of durum wheat mycobiota in all three cultivation environments. Previous studies have shown the presence of
Fusarium spp. in the wheat grains fungal community globally [
9,
25,
59,
61,
62,
67,
68,
69,
70] as well as in several Italian regions, including those investigated in this research [
7,
8,
31,
33,
71] as well as in others [
8,
18,
19,
32]. In detail, Infantino et al. [
32] and Shah et al. [
18] showed that
Fusarium incidence decreased from Northern to Southern Italian areas. Also in this study, differences among the three cultivation areas were observed. In general,
Fusarium incidence was higher in Emilia Romagna followed by Umbria and then Sardinia, showing that durum wheat cultivated in Northern Italy might be exposed to a higher FHB infection risk, followed by Central Italy. This suggests that environmental conditions at the regional level (macroscale) play a remarkable effect on
Fusarium incidence in durum wheat grains, also confirming that the gradual expansion of durum wheat from Southern to Northern areas of Italy might have increased the risks of FHB infections [
4]. However, Scala et al. [
19] suggested that environmental conditions at the local level (microscale) might drive FHB outbreaks even in areas suitable for growing durum wheat, such as Southern Italy. One of the reasons that may also have contributed to these regional differences (Emilia Romagna > Umbria > Sardinia) is represented by the rainfalls recorded during the anthesis phase of the crop (
Figure 4). In fact, concerning the season investigated in this research, the rainfall levels occurred in the three surveyed regions followed the same gradient as of
Fusarium infections. However, in association with climatic parameters, also other factors, such as crop rotation, fungicide application and cultivated varieties [
49,
51] may have played a role in
Fusarium incidence in wheat grains.
In this study, the different Fusarium species were identified by partial tef1α region sequencing. With regard to the used isolation method (PDA or DFB), F. proliferatum growth was particularly promoted by DFB with respect to PDA. The lower F. proliferatum development on PDA might have been caused by the sterilization process applied before seed plating as well as by the quicker development of other fungal species. For this reason, the use of both isolation methods could be recommended to obtain a realistic indication of the Fusarium community associated with grains. In fact, the choice of a specific isolation method rather than another could lead to the underestimation or the overestimation of the incidence of one or more Fusarium species.
With respect to the detected
Fusarium spp., each of the three examined regions was characterized by the presence of a specific complex of different species. In general, a predominance of
F. poae, in particular in Emilia Romagna and Umbria, was recorded. Earlier surveys conducted in Italy showed the prevalence of this species in several growing seasons [
8,
18,
32,
33], particularly those characterized by unfavorable climatic conditions to the most damaging FHB agents, such as
F. graminearum, at anthesis [
31]. In fact, in Umbria,
F. poae was the most frequently detected species in malting barley grains in 2014 [
72] and, together with
F. avenaceum [
7,
73], they seemed to temporarily replace
F. graminearum as the main components of the FHB complex associated with wheat and barley. In addition, this study shows that, for the analyzed year and particularly based on PDA isolations,
F. poae was the dominant species of the FHB complex in durum wheat grains harvested in the three different examined Italian regions.
F. avenaceum, at least in Emilia Romagna and Umbria, never exceeded, in the examined year,
F. poae. However, in Umbria,
F. avenaceum was present at similar levels as of
F. graminearum. This is a further confirmation of what previously observed in the Umbria region over the last few years, where
F. poae and
F. avenaceum seemed to interchange as the dominant members of the
Fusarium community associated with durum wheat and malting barley, while,
F. graminearum showed a constant incidence, never acting as the main species of the complex [
7,
72,
73,
74]. Interestingly, particularly based on DFB isolations,
F. proliferatum showed a high incidence in the FHB complex of durum wheat grains harvested in all investigated regions. This species is typically associated with maize grains where it represents one of the ear rot causal agents, however, across the years, the occurrence of
F. proliferatum in wheat has increased in many world cultivation areas [
75] including Italy, and in particular in the Emilia Romagna region [
76].
The regional differences observed for the incidence of the genus
Fusarium was also recorded for the incidence of the single members within the complex. In particular,
F. poae,
F. graminearum and
F. proliferatum were all extensively present in the Northern area followed by Central and finally Southern areas. Noteworthy is the fact that
F. graminearum was totally absent in the samples from Sardinia. This highlights, as observed in other world cultivation areas [
30,
52,
54,
77], that also in Italy the environment plays an important role in the FHB complex composition both under quantitative (incidence of some species within the complex) and qualitative (presence/absence of certain species within the complex) points of view.
To further explore the FHB complex composition, the fungal biomass of six selected main
Fusarium species was quantified directly in grains by qPCR. This method confirmed that the total
Fusarium fungal biomass (sum of the fungal biomasses of each of the six species analyzed) as well as
F. poae biomass were found to be higher in the Northern area, followed by Central and, finally, Southern areas. The absence of
F. graminearum in the samples harvested in Sardinia was also confirmed by qPCR. Interestingly,
F. langsethiae was detected by qPCR in all the investigated regions, while, it was isolated with both methods only in the grains harvested in Umbria and with a low incidence. This may have happened because
F. langsethiae, a fungus characterized by a slow growth rate, might have been overgrown during the isolation process by other more rapidly growing species (such as
F. poae) [
38]. Therefore, to have a complete overview of the FHB complex composition of a certain cultivation area, the use of methods able to molecularly detect and quantify the different species directly in the grains, such as qPCR, may also be used. By contrast, the adoption of species-specific primers, such as those used in this study, may limit the detection to a few species only. As a consequence, the simultaneous adoption of qPCR assays for in grains fungal biomass quantification as well as of isolation methods coupled to
tef1α region sequencing of the obtained
Fusarium isolates, even if it is a time-consuming procedure, allows more realistic information to be obtained about the
Fusarium community associated with durum wheat grains in a specific cultivation area.
Finally, as reported for other mycotoxigenic fungi [
78] the observed regional differences in
Fusarium incidence and FHB composition have important consequences for the quality of raw materials of derived products as well as for final consumers’ health.
In fact, the fungal secondary metabolites associated with durum wheat grains broadly reflected, with a few exceptions, the FHB complex composition obtained with both the use of the two isolation methods and qPCR assays. Besides, durum wheat grains harvested in the Northern area were more contaminated by
Fusarium mycotoxins than those of Central and Southern areas. In particular, probably due to the higher presence of
F. graminearum, trichothecenes, and DON among them, were particularly present in the grains harvested in Emilia Romagna. These samples also showed the highest BEA and ENs levels, possibly because of the highest
F. poae and
F. avenaceum presence, respectively. This highlights that, also in Italy, regional differences, as detected in other world areas [
49,
77], have a marked influence on the final
Fusarium mycotoxin contamination. Surprisingly, despite the significant
F. proliferatum presence, very low fumonisin levels (compared to those recovered in maize by Covarelli et al. [
79]) were detected (a very low percentage of samples only in Emilia Romagna). Low levels of fumonisin contamination on wheat infected with
F. proliferatum were also recorded in other wheat-cultivation areas [
74,
80].

 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}