Staphylococcus aureus Pathogenicity in Cystic Fibrosis Patients—Results from an Observational Prospective Multicenter Study Concerning Virulence Genes, Phylogeny, and Gene Plasticity


 and
and
Abstract
1. Introduction
2. Results
2.1. Demographics and Statistics
2.2. Quantity of Virulence Genes and Clinical Parameters
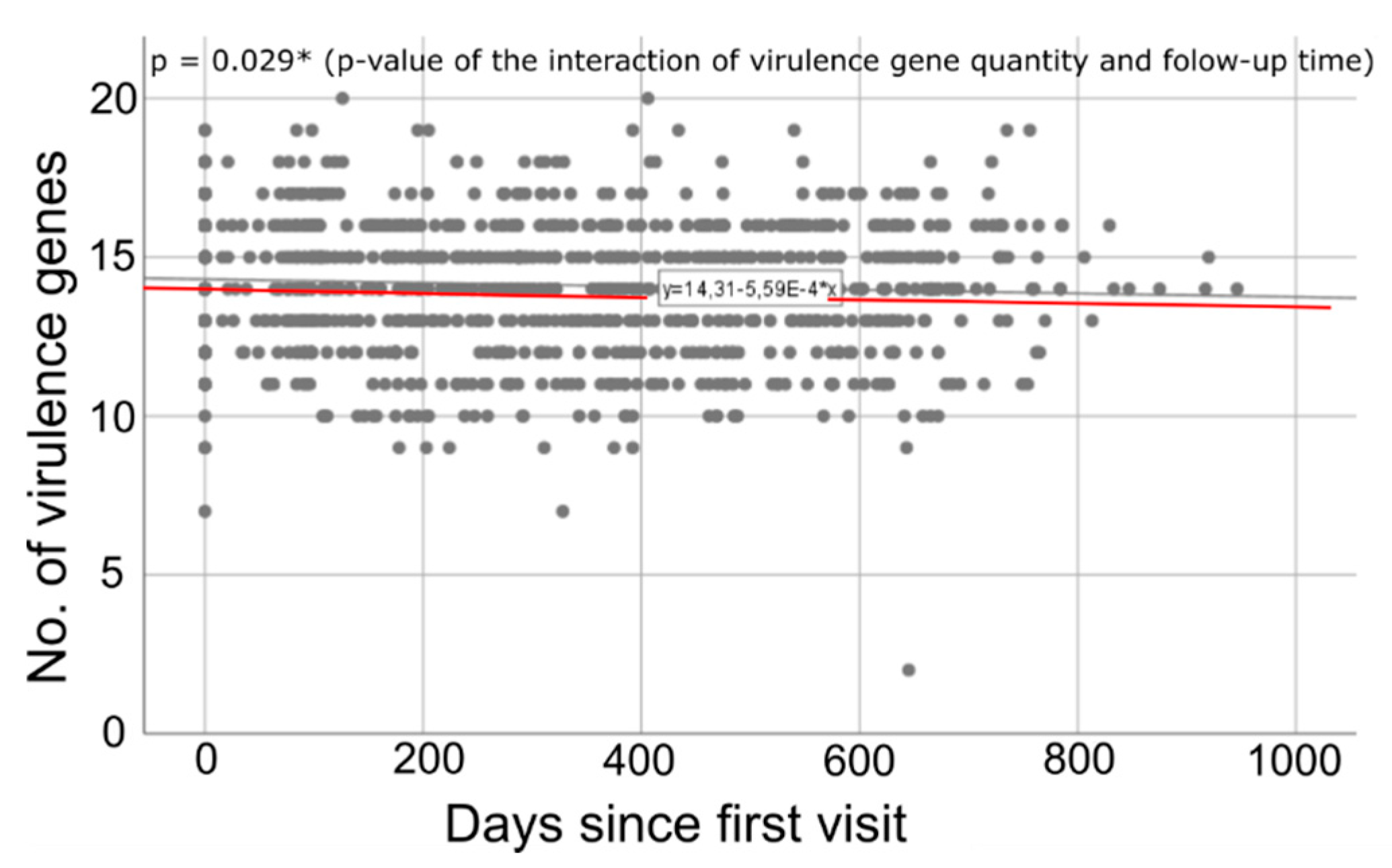
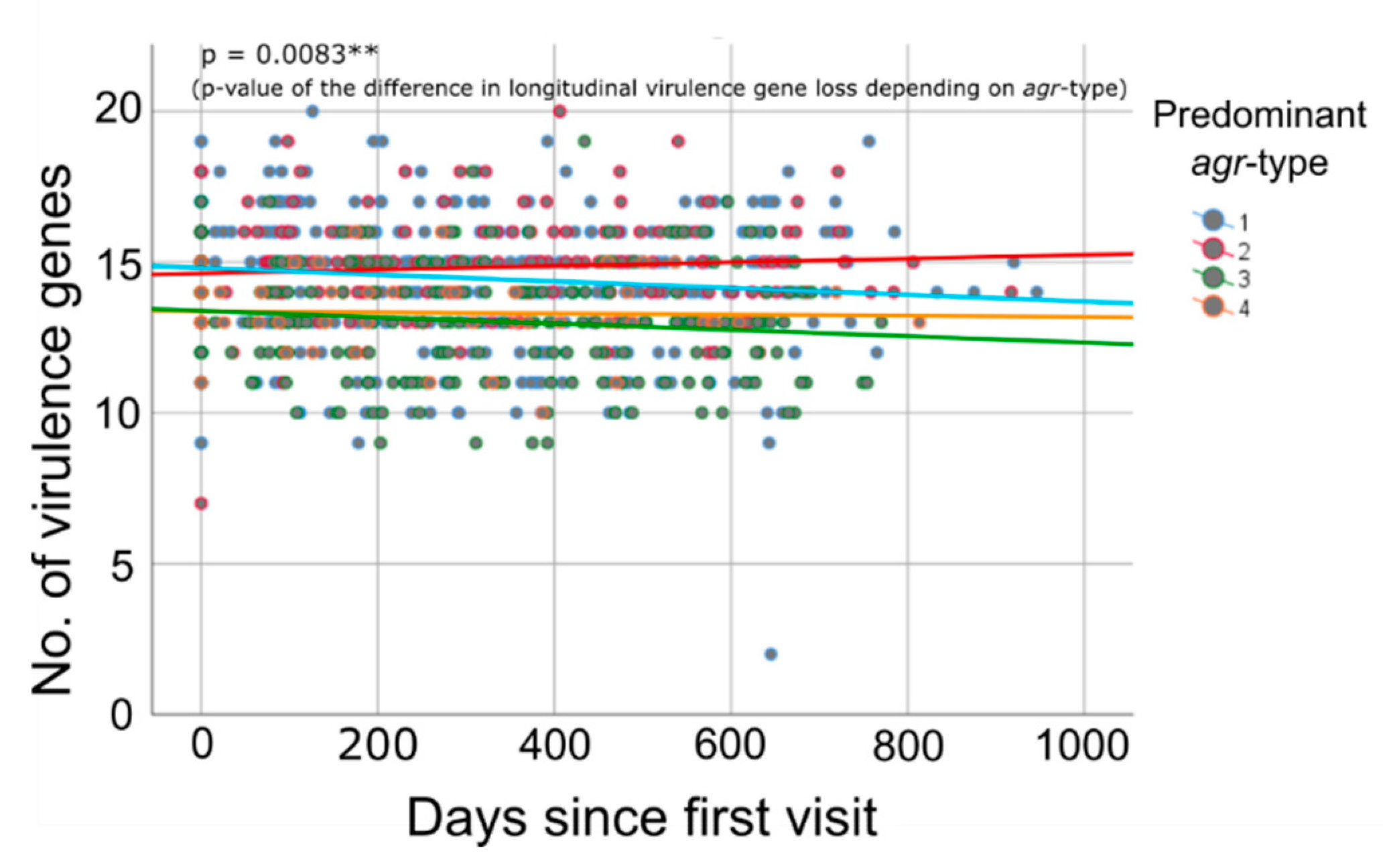
2.3. Time-Dependent Quantity of Virulence Genes
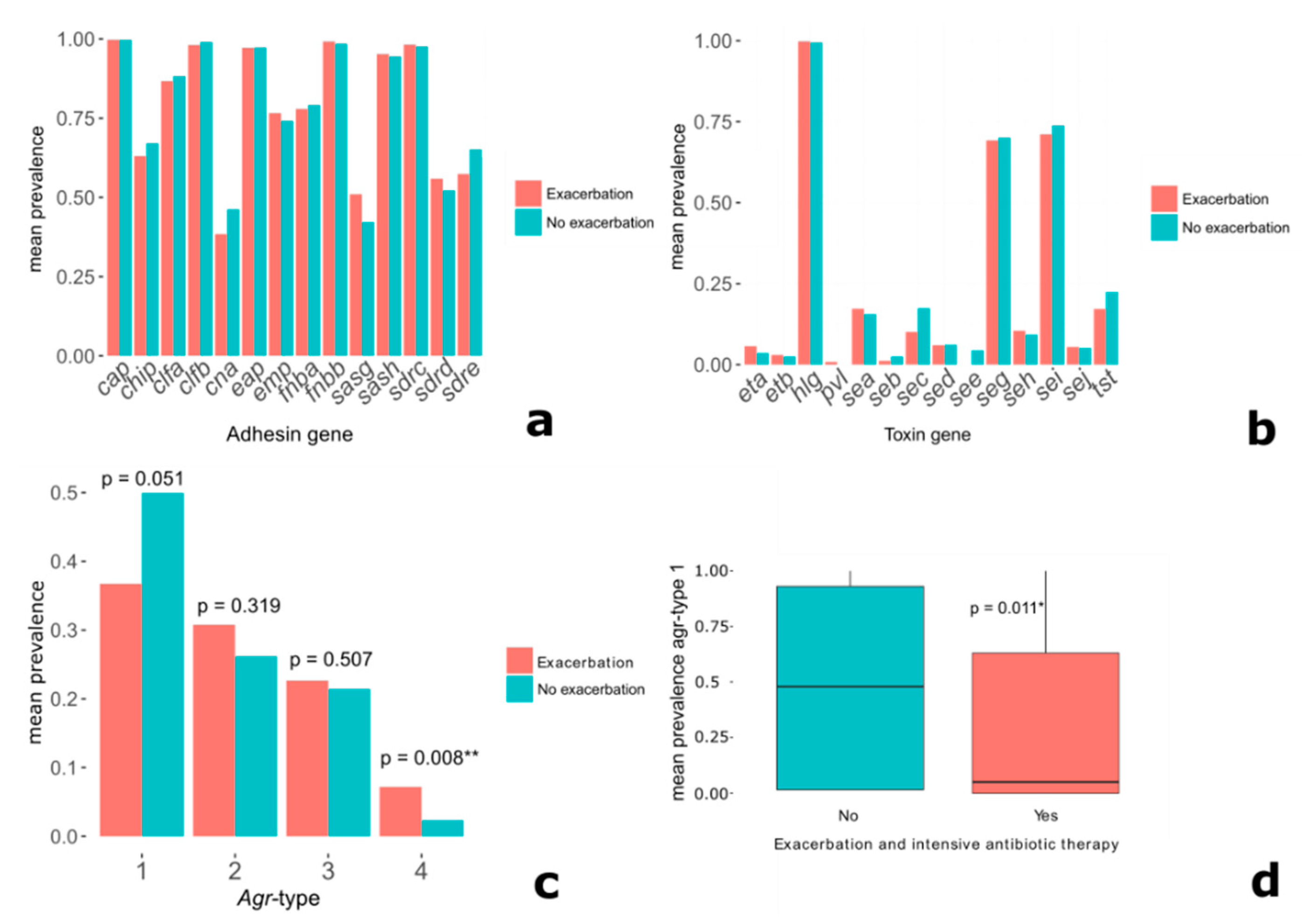
2.4. Specific Virulence Genes and Clinical Parameters
2.5. Complementary Data Concerning a Humoral Response Towards S. aureus Antigens
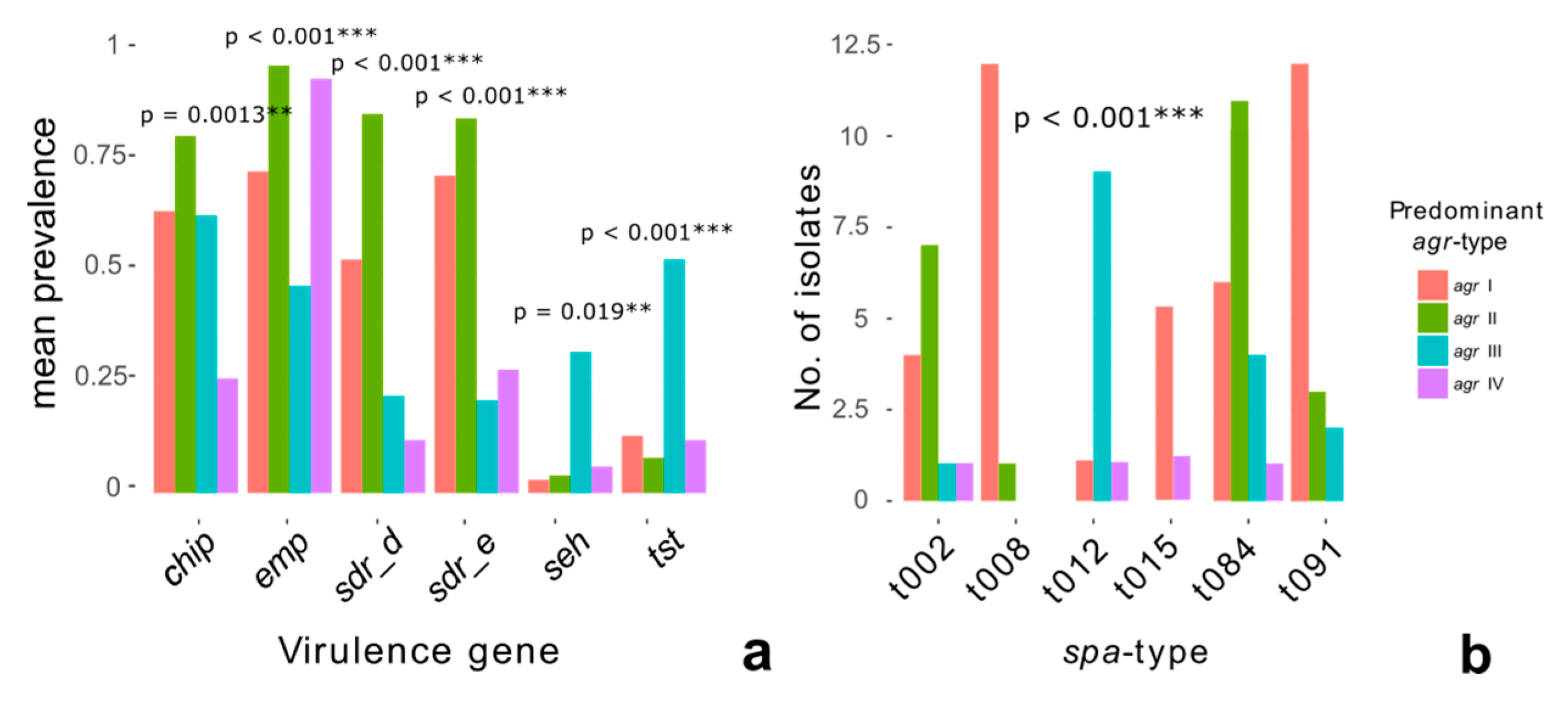
2.6. Phylogeny and Virulence Genes
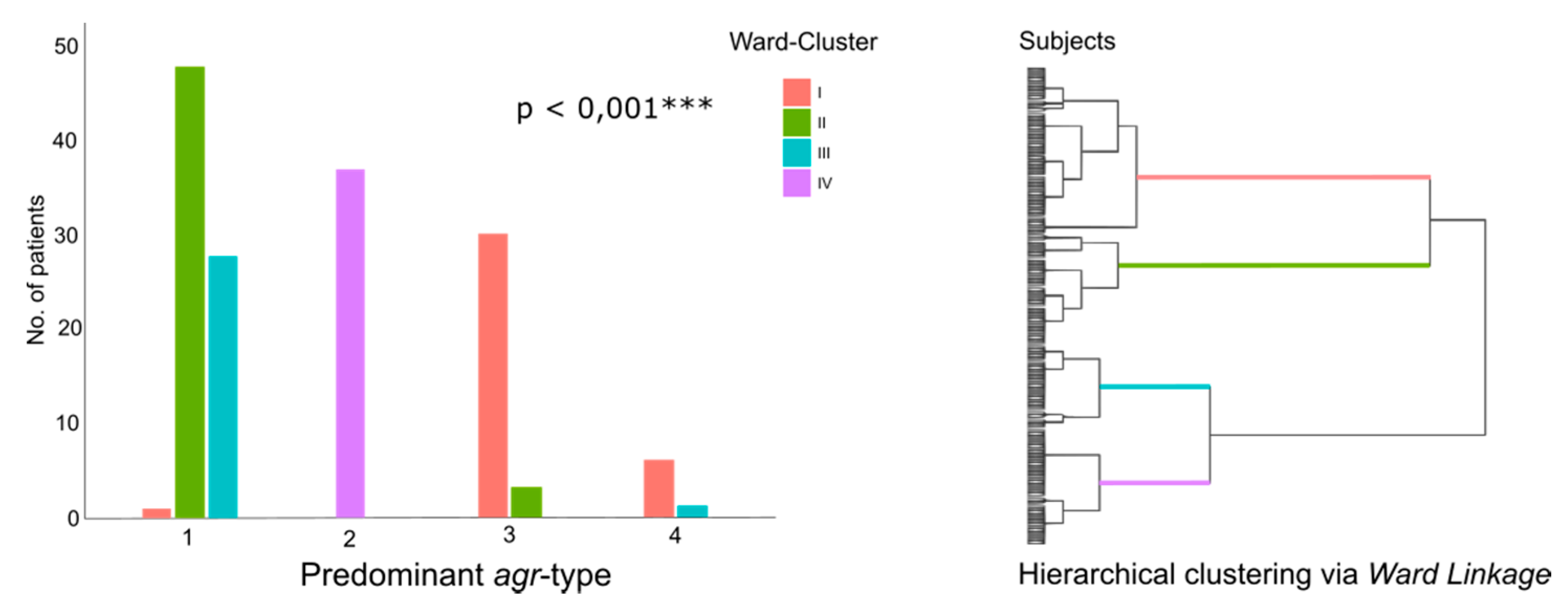
2.7. Cluster Analyses
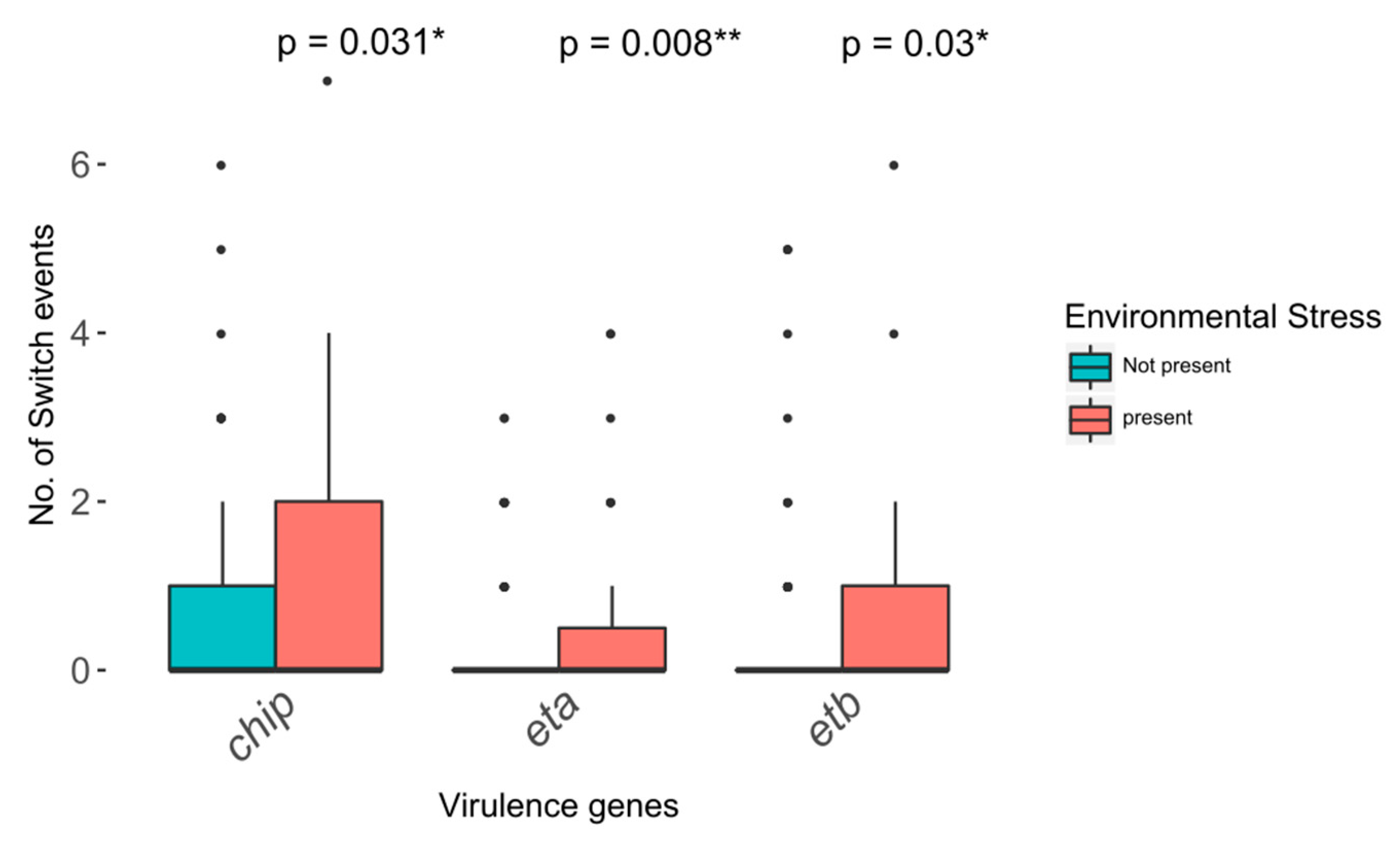
2.8. Virulence Gene Plasticity and Environmental Stress
3. Discussion
3.1. Specific Virulence Genes and Clinical Parameters
3.2. Phylogeny and Pathogenicity
3.3. Longitudinal Decline in Virulence Gene Quantity
3.4. Virulence Gene Plasticity and Environmental Stress
3.5. Strengths and Limitations of the Study and Analyses
4. Conclusions
5. Material and Methods
5.1. Study Design
5.2. Virulence Gene Profiles
5.3. Parameters
5.3.1. Exacerbation
5.3.2. Lung Function
5.3.3. Virulence Gene Prevalence Counts
5.3.4. Quantity of Virulence Genes
5.3.5. Agr-Type
5.3.6. Environmental Stress
5.3.7. Virulence Gene Plasticity and Horizontal Gene Transfer
5.3.8. Antistaphylococcal Antibodies
5.4. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Elborn, J.S. Cystic fibrosis. Lancet 2016, 388, 2519–2531. [Google Scholar] [CrossRef]
- Farrell, P.M. The prevalence of cystic fibrosis in the European Union. J. Cyst. Fibros. 2008, 7, 450–453. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, M.; Gibson, R.L.; McNamara, S.; Emerson, J.; Burns, J.L.; Castile, R.; Hiatt, P.; McCoy, K.; Wilson, C.B.; Inglis, A.; et al. Early pulmonary infection, inflammation, and clinical outcomes in infants with cystic fibrosis. Pediatr. Pulmonol. 2001, 32, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Kahl, B.C. Impact of Staphylococcus aureus on the pathogenesis of chronic cystic fibrosis lung disease. Int. J. Med. Microbiol. 2010, 300, 514–519. [Google Scholar] [CrossRef]
- Barley, M.; Mcnally, J.; Marshall, B.; Faro, A.; Elbert, A.; Fink, A.; Sewall, A.; Loeffler, D.; Petren, K.; O’Neil, T.; et al. Annual Data Report 2016 Cystic Fibrosis Foundation Patient Registry. Cyst. Fibros. Found. Patient Regist. 2016, 2016, 1–92. [Google Scholar]
- Goerke, C.; Wolz, C. Adaptation of Staphylococcus aureus to the cystic fibrosis lung. Int. J. Med. Microbiol. 2010, 300, 520–525. [Google Scholar] [CrossRef]
- Westphal, C.; Görlich, D.; Kampmeier, S.; Herzog, S.; Braun, N.; Hitschke, C.; Mellmann, A.; Peters, G.; Kahl, B.C.; Junge, S.; et al. Antibiotic Treatment and Age Are Associated with Staphylococcus aureus Carriage Profiles During Persistence in the Airways of Cystic Fibrosis Patients. Front. Microbiol. 2020, 11, 230. [Google Scholar] [CrossRef]
- Ankrum, A.; Hall, B.G. Population dynamics of Staphylococcus aureus in cystic fibrosis patients to determine transmission events by use of whole-genome sequencing. J. Clin. Microbiol. 2017, 55, 2143–2152. [Google Scholar] [CrossRef]
- Azarian, T.; Ridgway, J.P.; Yin, Z.; David, M.Z. Long-Term Intrahost Evolution of Methicillin Resistant Staphylococcus aureus Among Cystic Fibrosis Patients With Respiratory Carriage. Front. Genet. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Hirschhausen, N.; Block, D.; Bianconi, I.; Bragonzi, A.; Birtel, J.; Lee, J.C.; Dübbers, A.; Küster, P.; Kahl, J.; Peters, G.; et al. Extended Staphylococcus aureus persistence in cystic fibrosis is associated with bacterial adaptation. Int. J. Med. Microbiol. 2013, 303, 685–692. [Google Scholar] [CrossRef]
- Langhanki, L.; Berger, P.; Treffon, J.; Catania, F.; Kahl, B.C.; Mellmann, A. In vivo competition and horizontal gene transfer among distinct Staphylococcus aureus lineages as major drivers for adaptational changes during long-term persistence in humans. BMC Microbiol. 2018, 18, 152. [Google Scholar] [CrossRef] [PubMed]
- Goerke, C.; Gressinger, M.; Endler, K.; Breitkopf, C.; Wardecki, K.; Stern, M.; Wolz, C.; Kahl, B.C. High phenotypic diversity in infecting but not in colonizing Staphylococcus aureus populations. Environ. Microbiol. 2007, 9, 3134–3142. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival Mechanisms of Clinically Relevant Microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [PubMed]
- Schwartbeck, B.; Birtel, J.; Treffon, J.; Langhanki, L.; Mellmann, A.; Kale, D.; Kahl, J.; Hirschhausen, N.; Neumann, C.; Lee, J.C.; et al. Dynamic in vivo mutations within the ica operon during persistence of Staphylococcus aureus in the airways of cystic fibrosis patients. PLoS Pathog. 2016, 12, e1006024. [Google Scholar] [CrossRef] [PubMed]
- Kahl, B.; Herrmann, M.; Everding, A.S.; Koch, H.G.; Becker, K.; Harms, E.; Proctor, R.A.; Peters, G. Persistent Infection with Small Colony Variant Strains of Staphylococcus aureus in Patients with Cystic Fibrosis. J. Infect. Dis. 1998, 177, 1023–1029. [Google Scholar] [CrossRef]
- Wolter, D.J.; Emerson, J.C.; McNamara, S.; Buccat, A.M.; Qin, X.; Cochrane, E.; Houston, L.S.; Rogers, G.B.; Marsh, P.; Prehar, K.; et al. Staphylococcus aureus Small-Colony Variants Are Independently Associated with Worse Lung Disease in Children with Cystic Fibrosis. Clin. Infect. Dis. 2013, 57, 384–391. [Google Scholar] [CrossRef]
- Besier, S.; Ludwig, A.; Ohlsen, K.; Brade, V.; Wichelhaus, T.A. Molecular analysis of the thymidine-auxotrophic small colony variant phenotype of Staphylococcus aureus. Int. J. Med. Microbiol. 2007, 297, 217–225. [Google Scholar] [CrossRef]
- Garzoni, C.; Kelley, W.L. Staphylococcus aureus: New evidence for intracellular persistence. Trends Microbiol. 2009, 17, 59–65. [Google Scholar] [CrossRef]
- Goerke, C.; Campana, S.; Bayer, M.G.; Döring, G.; Botzenhart, K.; Wolz, C.; Do, G.; Botzenhart, K.; Wolz, C. Direct quantitative transcript analysis of the agr regulon of Staphylococcus aureus during human infection in comparison to the expression profile in vitro. Infect. Immun. 2000, 68, 1304–1311. [Google Scholar] [CrossRef]
- Herzog, S.; Dach, F.; de Buhr, N.; Niemann, S.; Schlagowski, J.; Chaves-Moreno, D.; Neumann, C.; Goretzko, J.; Schwierzeck, V.; Mellmann, A.; et al. High nuclease activity of long persisting Staphylococcus aureus isolates within the airways of cystic fibrosis patients protects against NET-mediated killing. Front. Immunol. 2019, 10, 2552. [Google Scholar] [CrossRef]
- Treffon, J.; Block, D.; Moche, M.; Reiss, S.; Fuchs, S.; Engelmann, S.; Becher, D.; Langhanki, L.; Mellmann, A.; Peters, G.; et al. Adaptation of Staphylococcus aureus to airway environments in patients with cystic fibrosis by upregulation of superoxide dismutase M and iron-scavenging proteins. J. Infect. Dis. 2018, 217, 1453–1461. [Google Scholar] [CrossRef] [PubMed]
- Treffon, J.; Chaves-Moreno, D.; Niemann, S.; Pieper, D.H.; Vogl, T.; Roth, J.; Kahl, B.C. Importance of superoxide dismutases A and M for protection of Staphylococcus aureus in the oxidative stressful environment of cystic fibrosis airways. Cell. Microbiol. 2020, 22, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Gangell, C.; Gard, S.; Douglas, T.; Park, J.; De Klerk, N.; Keil, T.; Brennan, S.; Ranganathan, S.; Robins-Browne, R.; Sly, P.D. Inflammatory responses to individual microorganisms in the lungs of children with cystic fibrosis. Clin. Infect. Dis. 2011, 53, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, D.S.; Hook, S.M.; Jamsen, K.M.; Nixon, G.M.; Carzino, R.; Carlin, J.B.; Robertson, C.F.; Grimwood, K. Lower airway inflammation in infants with cystic fibrosis detected by newborn screening. Pediatr. Pulmonol. 2005, 40, 500–510. [Google Scholar] [CrossRef]
- Sagel, S.D.; Gibson, R.L.; Emerson, J.; McNamara, S.; Burns, J.L.; Wagener, J.S.; Ramsey, B.W.; Konstan, M.; Chatfield, B.; Retsch-Bogart, G.; et al. Impact of Pseudomonas and Staphylococcus Infection on Inflammation and Clinical Status in Young Children with Cystic Fibrosis. J. Pediatr. 2009, 154, 183–188. [Google Scholar] [CrossRef]
- Junge, S.; Görlich, D.; Den Reijer, M.; Wiedemann, B.; Tümmler, B.; Ellemunter, H.; Dübbers, A.; Küster, P.; Ballmann, M.; Koerner-Rettberg, C.; et al. Factors associated with worse lung function in cystic fibrosis patients with persistent Staphylococcus aureus. PLoS ONE 2016, 11, e0166220. [Google Scholar] [CrossRef]
- Smyth, A. Prophylactic antibiotics in cystic fibrosis: A conviction without evidence? Pediatr. Pulmonol. 2005, 40, 471–476. [Google Scholar] [CrossRef]
- Hurley, M.N.; Smyth, A.R. Staphylococcus aureus in cystic fibrosis: Pivotal role or bit part actor? Curr. Opin. Pulm. Med. 2018, 24, 586–591. [Google Scholar] [CrossRef]
- Westphal, C.; Görlich, D.; Herzog, S.; Braun, N.; Vogel, C.; Peters, G.; Kahl, B. Antibiotic treatment and age have an impact on Staphylococcus aureus clonality during persistence in the airways of cystic fibrosis patients. Front Microbiol. 2020, 11, 230. [Google Scholar] [CrossRef]
- den Reijer, P.M.; Lemmens-den Toom, N.; Kant, S.; Snijders, S.V.; Boelens, H.; Tavakol, M.; Verkaik, N.J.; van Belkum, A.; Verbrugh, H.A.; van Wamel, W.J.B. Characterization of the Humoral Immune Response during Staphylococcus aureus Bacteremia and Global Gene Expression by Staphylococcus aureus in Human Blood. PLoS ONE 2013, 8, e53391. [Google Scholar] [CrossRef]
- Verkaik, N.J.; de Vogel, C.P.; Boelens, H.A.; Grumann, D.; Hoogenboezem, T.; Vink, C.; Hooijkaas, H.; Foster, T.J.; Verbrugh, H.A.; van Belkum, A.; et al. Anti-Staphylococcal Humoral Immune Response in Persistent Nasal Carriers and Noncarriers of Staphylococcus aureus. J. Infect. Dis. 2009, 199, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Chadha, A.D.; Thomsen, I.P.; Jimenez-Truque, N.; Soper, N.R.; Jones, L.S.; Sokolow, A.G.; Torres, V.J.; Creech, C.B. Host response to Staphylococcus aureus cytotoxins in children with cystic fibrosis. J. Cyst. Fibros. 2016, 15, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Goerke, C.; Koller, J.; Wolz, C. Ciprofloxacin and trimethoprim cause phage induction and virulence modulation in Staphylococcus aureus. Antimicrob. Agents Chemother. 2006, 50, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Goerke, C.; Matias y Papenberg, S.; Dasbach, S.; Dietz, K.; Ziebach, R.; Kahl, B.C.; Wolz, C. Increased frequency of genomic alterations in Staphylococcus aureus during chronic infection is in part due to phage mobilization. J. Infect. Dis. 2004, 189, 724–734. [Google Scholar] [CrossRef]
- Úbeda, C.; Maiques, E.; Knecht, E.; Lasa, Í.; Novick, R.P.; Penadés, J.R. Antibiotic-induced SOS response promotes horizontal dissemination of pathogenicity island-encoded virulence factors in staphylococci. Mol. Microbiol. 2005, 56, 836–844. [Google Scholar] [CrossRef]
- Lindsay, J.A.; Moore, C.E.; Day, N.P.; Peacock, S.J.; Witney, A.A.; Stabler, R.A.; Husain, S.E.; Butcher, P.D.; Hinds, J. Microarrays reveal that each of the ten dominant lineages of Staphylococcus aureus has a unique combination of surface-associated and regulatory genes. J. Bacteriol. 2006, 188, 669–676. [Google Scholar] [CrossRef]
- Shallcross, L.J.; Fragaszy, E.; Johnson, A.M.; Hayward, A.C. The role of the Panton-Valentine leucocidin toxin in staphylococcal disease: A systematic review and meta-analysis. Lancet Infect. Dis. 2013, 13, 43–54. [Google Scholar] [CrossRef]
- Laabei, M.; Uhlemann, A.C.; Lowy, F.D.; Austin, E.D.; Yokoyama, M.; Ouadi, K.; Feil, E.; Thorpe, H.A.; Williams, B.; Perkins, M.; et al. Evolutionary Trade-Offs Underlie the Multi-faceted Virulence of Staphylococcus aureus. PLoS Biol. 2015, 13, e1002229. [Google Scholar] [CrossRef]
- Jarraud, S.; Mougel, C.; Thioulouse, J.; Lina, G.; Meugnier, H.; Forey, F.; Nesme, X.; Etienne, J.; Vandenesch, F. Relationships between Staphylococcus aureus genetic background, virulence factors, agr groups (alleles), and human disease. Infect. Immun. 2002, 70, 631–641. [Google Scholar] [CrossRef]
- Cafiso, V.; Bertuccio, T.; Spina, D.; Campanile, F.; Bongiorno, D.; Santagati, M.; Sciacca, A.; Sciuto, C.; Stefani, S. Methicillin resistance and vancomycin heteroresistance in Staphylococcus aureus in cystic fibrosis patients. Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 1277–1285. [Google Scholar] [CrossRef]
- Shilo, N.; Quach, C. Pulmonary Infections and Community Associated Methicillin Resistant Staphylococcus aureus: A Dangerous Mix? Paediatr. Respir. Rev. 2011, 12, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Glikman, D.; Siegel, J.D.; David, M.Z.; Okoro, N.M.; Boyle-Vavra, S.; Dowell, M.L.; Daum, R.S. Complex Molecular Epidemiology of Methicillin-Resistant Staphylococcus aureus Isolates from Children with Cystic Fibrosis in the Era of Epidemic Community-Associated Methicillin-Resistant S aureus. Chest 2008, 133, 1381–1387. [Google Scholar] [CrossRef] [PubMed]
- Goerke, C.; Kümmel, M.; Dietz, K.; Wolz, C. Evaluation of intraspecies interference due to agr polymorphism in Staphylococcus aureus during infection and colonization. J. Infect. Dis. 2003, 188, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.S.; Traber, K.E.; Corrigan, R.; Benson, S.A.; Musser, J.M.; Novick, R.P. The agr Radiation: An Early Event in the Evolution of Staphylococci. J. Bacteriol. 2005, 187, 5585–5594. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.; Singh, P.K. Evolving stealth: Genetic adaptation of Pseudomonas aeruginosa during cystic fibrosis infections. Proc. Natl. Acad. Sci. USA 2006, 103, 8305–8306. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.E.; Buckley, D.G.; Wu, Z.; Saenphimmachak, C.; Hoffman, L.R.; D’Argenio, D.A.; Miller, S.I.; Ramsey, B.W.; Speert, D.P.; Moskowitz, S.M.; et al. Genetic adaptation by Pseudomonas aeruginosa to the airways of cystic fibrosis patients. Proc. Natl. Acad. Sci. USA 2006, 103, 8487–8492. [Google Scholar] [CrossRef]
- Knöppel, A.; Lind, P.A.; Lustig, U.; Näsvall, J.; Andersson, D.I. Minor fitness costs in an experimental model of horizontal gene transfer in bacteria. Mol. Biol. Evol. 2014, 31, 1220–1227. [Google Scholar] [CrossRef]
- Sorek, R.; Zhu, Y.; Creevey, C.J.; Francino, M.P.; Bork, P.; Rubin, E.M. Genome-Wide Experimental Determination of Barriers to Horizontal Gene Transfer. Science 2007, 318, 1449–1452. [Google Scholar] [CrossRef]
- Yachi, S.; Loreau, M. Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proc. Natl. Acad. Sci. USA 1999, 96, 1463–1468. [Google Scholar] [CrossRef]
- Hodille, E.; Warre, R.; Diep, B.A.; Goutelle, S.; Lina, G.; Dumitrescu, O. The role of antibiotics in modulating virulence in Staphylococcus aureus. Clin. Microbiol. Rev. 2017, 30, 887–917. [Google Scholar] [CrossRef]
- Liou, T.G.; Elkin, E.P.; Pasta, D.J.; Jacobs, J.R.; Konstan, M.W.; Morgan, W.J.; Wagener, J.S. Year-to-year changes in lung function in individuals with cystic fibrosis. J. Cyst. Fibros. 2010, 9, 250–256. [Google Scholar] [CrossRef]
- Kerem, E.; Viviani, L.; Zolin, A.; MacNeill, S.; Hatziagorou, E.; Ellemunter, H.; Drevinek, P.; Gulmans, V.; Krivec, U.; Olesen, H. Factors associated with FEV1 decline in cystic fibrosis: Analysis of the ECFS patient registry. Eur. Respir. J. 2014, 43, 125–133. [Google Scholar] [CrossRef] [PubMed]
- McKone, E.F.; Emerson, S.S.; Edwards, K.L.; Aitken, M.L. Effect of genotype on phenotype and mortality in cystic fibrosis: A retrospective cohort study. Lancet 2003, 361, 1671–1676. [Google Scholar] [CrossRef]
- Wolf, C.; Kusch, H.; Monecke, S.; Albrecht, D.; Holtfreter, S.; von Eiff, C.; Petzl, W.; Rainard, P.; Bröker, B.M.; Engelmann, S. Genomic and proteomic characterization of Staphylococcus aureus mastitis isolates of bovine origin. Proteomics 2011, 11, 2491–2502. [Google Scholar] [CrossRef] [PubMed]
- Ziebandt, A.K.; Kusch, H.; Degner, M.; Jaglitz, S.; Sibbald, M.J.J.B.; Arends, J.P.; Chlebowicz, M.A.; Albrecht, D.; Pantuček, R.; Doškar, J.; et al. Proteomics uncovers extreme heterogeneity in the Staphylococcus aureus exoproteome due to genomic plasticity and variant gene regulation. Proteomics 2010, 10, 1634–1644. [Google Scholar] [CrossRef]
- Sibbald, M.J.J.B.; Ziebandt, A.K.; Engelmann, S.; Hecker, M.; de Jong, A.; Harmsen, H.J.M.; Raangs, G.C.; Stokroos, I.; Arends, J.P.; Dubois, J.Y.F.; et al. Mapping the Pathways to Staphylococcal Pathogenesis by Comparative Secretomics. Microbiol. Mol. Biol. Rev. 2006, 70, 755–788. [Google Scholar] [CrossRef]
- Hogardt, M.; Haeußler, S.; Balke, B.; Kahl, B.C.; Schmoldt, S.; Leitritz, L.; Jäger, G.; Kappler, M.; Suerbaum, S.; Heesemann, J. MIQ 24: Atemwegsinfektionen bei Mukoviszidose Qualitätsstandards in der Mikrobiologisch-Infektiologischen Diagnostik; Elsevier, Urban Fischer: München, Germany, 2006. [Google Scholar]
- Harmsen, D.; Claus, H.; Witte, W.; Rothganger, J.; Claus, H.; Turnwald, D.; Vogel, U. Typing of Methicillin-Resistant Staphylococcus aureus in a University Hospital Setting by Using Novel Software for spa Repeat Determination and Database Management. J. Clin. Microbiol. 2003, 41, 5442–5448. [Google Scholar] [CrossRef]
- Mellmann, A.; Weniger, T.; Berssenbrügge, C.; Rothgänger, J.; Sammeth, M.; Stoye, J.; Harmsen, D. Based Upon Repeat Pattern (BURP): An algorithm to characterize the long-term evolution of Staphylococcus aureus populations based on spa polymorphisms. BMC Microbiol. 2007, 7. [Google Scholar] [CrossRef]
- van Wamel, W.J.B.; Rooijakkers, S.H.M.; Ruyken, M.; van Kessel, K.P.M.; van Strijp, J.A.G. The Innate Immune Modulators Staphylococcal Complement Inhibitor and Chemotaxis Inhibitory Protein of Staphylococcus aureus Are Located on -Hemolysin-Converting Bacteriophages. J. Bacteriol. 2006, 188, 1310–1315. [Google Scholar] [CrossRef]
- Gomes, A.R.; Vinga, S.; Zavolan, M.; Lencastre, H. De Analysis of the Genetic Variability of Virulence-Related Loci in Epidemic Clones of Methicillin-Resistant Staphylococcus aureus Analysis of the Genetic Variability of Virulence-Related Loci in Epidemic Clones of Methicillin-Resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2005, 49, 366–379. [Google Scholar]
- Goerke, C.; Esser, S.; Kümmel, M.; Wolz, C. Staphylococcus aureus strain designation by agr and cap polymorphism typing and delineation of agr diversification by sequence analysis. Int. J. Med. Microbiol. 2005, 295, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Roche, F.M.; Massey, R.; Peacock, S.J.; Day, N.P.J.; Visai, L.; Speziale, P.; Lam, A.; Pallen, M.; Foster, T.J. Characterization of novel LPXTG-containing proteins of Staphylococcus aureus identified from genome sequences. Microbiology 2003, 149, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Friedrich, A.W.; Lubritz, G.; Weilert, M.; Peters, G.; Eiff, C. Von Prevalence of Genes Encoding Pyrogenic Toxin Superantigens and Exfoliative Toxins among Strains of Staphylococcus aureus isolated from blood and nasal specimens. J. Clin. Microbiol. 2003, 41, 1434–1439. [Google Scholar] [CrossRef] [PubMed]
- Lina, G.; Boutite, F.; Tristan, A.; Bes, M.; Etienne, J.; Vandenesch, F. Bacterial competition for human nasal cavity colonization: Role of Staphylococcal agr alleles. Appl. Environ. Microbiol. 2003, 69, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Von Eiff, C.; Sinha, B.; Joost, I.; Herrmann, M.; Peters, G.; Becker, K. eap gene as novel target for specific identification of Staphylococcus aureus. J. Clin. Microbiol. 2008, 46, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Becker, K.; Von Eiff, C.; Schrenzel, J.; Peters, G.; Herrmann, M. Identification and characterization of a novel 38.5-Kilodalton cell surface protein of Staphylococcus aureus with extended-spectrum binding activity for extracellular matrix and plasma proteins. J. Bacteriol. 2001, 183, 6778–6786. [Google Scholar] [CrossRef][Green Version]
- Fuchs, H.J.; Borowitz, D.S.; Christiansen, D.H.; Morris, E.M.; Nash, M.L.; Ramsey, B.W.; Rosenstein, B.J.; Smith, A.L.; Wohl, M.E. Effect of Aerosolized Recombinant Human DNase on Exacerbations of Respiratory Symptoms and on Pulmonary Function in Patients with Cystic Fibrosis. N. Engl. J. Med. 1994, 331, 637–642. [Google Scholar] [CrossRef]
- Quanjer, P.H.; Stanojevic, S.; Cole, T.J.; Baur, X.; Hall, G.L.; Culver, B.H.; Enright, P.L.; Hankinson, J.L.; Ip, M.S.M.; Zheng, J.; et al. Multi-ethnic reference values for spirometry for the 3–95-yr age range: The global lung function 2012 equations. Eur. Respir. J. 2012, 40, 1324–1343. [Google Scholar] [CrossRef]
- Recsei, P.; Kreiswirth, B.; O’Reilly, M.; Schlievert, P.; Gruss, A.; Novick, R.P. Regulation of exoprotein gene expression in Staphylococcus aureus by agr. MGG Mol. Gen. Genet. 1986, 202, 58–61. [Google Scholar] [CrossRef]
- Yarwood, J.M.; Schlievert, P.M. Quorum sensing in Staphylococcus infections. J. Clin. Invest. 2003, 112, 1620–1625. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Mean IgG Level All Patients (±SE) b | Mean IgG Level Controls (±SE) b | p Value c,d | Estimated Effect on FEV1% e | p Value d |
|---|---|---|---|---|---|
| CHIPS | 11492 (±220) | 11019 (±349) | 0.104 | −0.00073 | 0.0185 |
| ClfB | 4587 (±222) | 4092 (±338) | 0.3809 | 0.002079 | <0.0001 |
| Eta | 3999 (±447) | 2043 (±424) | 0.1180 | −0.00061 | 0.0033 |
| Etb | 613 (±113) | 320 (±94) | 0.0032 | −0.00700 | <0.0001 |
| HlgB | 13621 (±174) | 9878 (±407) | <0.0001 | −0.00109 | 0.0088 |
| LukF | 4079 (±133) | 2718 (±243) | <0.0001 | −0.00305 | <0.0001 |
| LukS | 14097 (±133) | 7134 (±524) | <0.0001 | −0.00169 | <0.0001 |
| SdrE | 3139 (±201) | 1992 (±221) | 0.0253 | −0.00179 | <0.0001 |
| SED | 1106 (±118) | 1292 (±268) | 0.0498 | 0.001490 | 0.0377 |
| SEH | 2290 (±291) | 2174 (±359) | 0.0113 | −0.00047 | 0.0986 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lange, J.; Heidenreich, K.; Higelin, K.; Dyck, K.; Marx, V.; Reichel, C.; Wamel, W.v.; Reijer, M.d.; Görlich, D.; Kahl, B.C. Staphylococcus aureus Pathogenicity in Cystic Fibrosis Patients—Results from an Observational Prospective Multicenter Study Concerning Virulence Genes, Phylogeny, and Gene Plasticity. Toxins 2020, 12, 279. https://doi.org/10.3390/toxins12050279
Lange J, Heidenreich K, Higelin K, Dyck K, Marx V, Reichel C, Wamel Wv, Reijer Md, Görlich D, Kahl BC. Staphylococcus aureus Pathogenicity in Cystic Fibrosis Patients—Results from an Observational Prospective Multicenter Study Concerning Virulence Genes, Phylogeny, and Gene Plasticity. Toxins. 2020; 12(5):279. https://doi.org/10.3390/toxins12050279
Chicago/Turabian StyleLange, Jonas, Kathrin Heidenreich, Katharina Higelin, Kristina Dyck, Vanessa Marx, Christian Reichel, Willem van Wamel, Martijn den Reijer, Dennis Görlich, and Barbara C. Kahl. 2020. "Staphylococcus aureus Pathogenicity in Cystic Fibrosis Patients—Results from an Observational Prospective Multicenter Study Concerning Virulence Genes, Phylogeny, and Gene Plasticity" Toxins 12, no. 5: 279. https://doi.org/10.3390/toxins12050279
APA StyleLange, J., Heidenreich, K., Higelin, K., Dyck, K., Marx, V., Reichel, C., Wamel, W. v., Reijer, M. d., Görlich, D., & Kahl, B. C. (2020). Staphylococcus aureus Pathogenicity in Cystic Fibrosis Patients—Results from an Observational Prospective Multicenter Study Concerning Virulence Genes, Phylogeny, and Gene Plasticity. Toxins, 12(5), 279. https://doi.org/10.3390/toxins12050279