Cytotoxic 4-Hydroxyprorocentrolide and Prorocentrolide C from Cultured Dinoflagellate Prorocentrum lima Induce Human Cancer Cell Death through Apoptosis and Cell Cycle Arrest



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
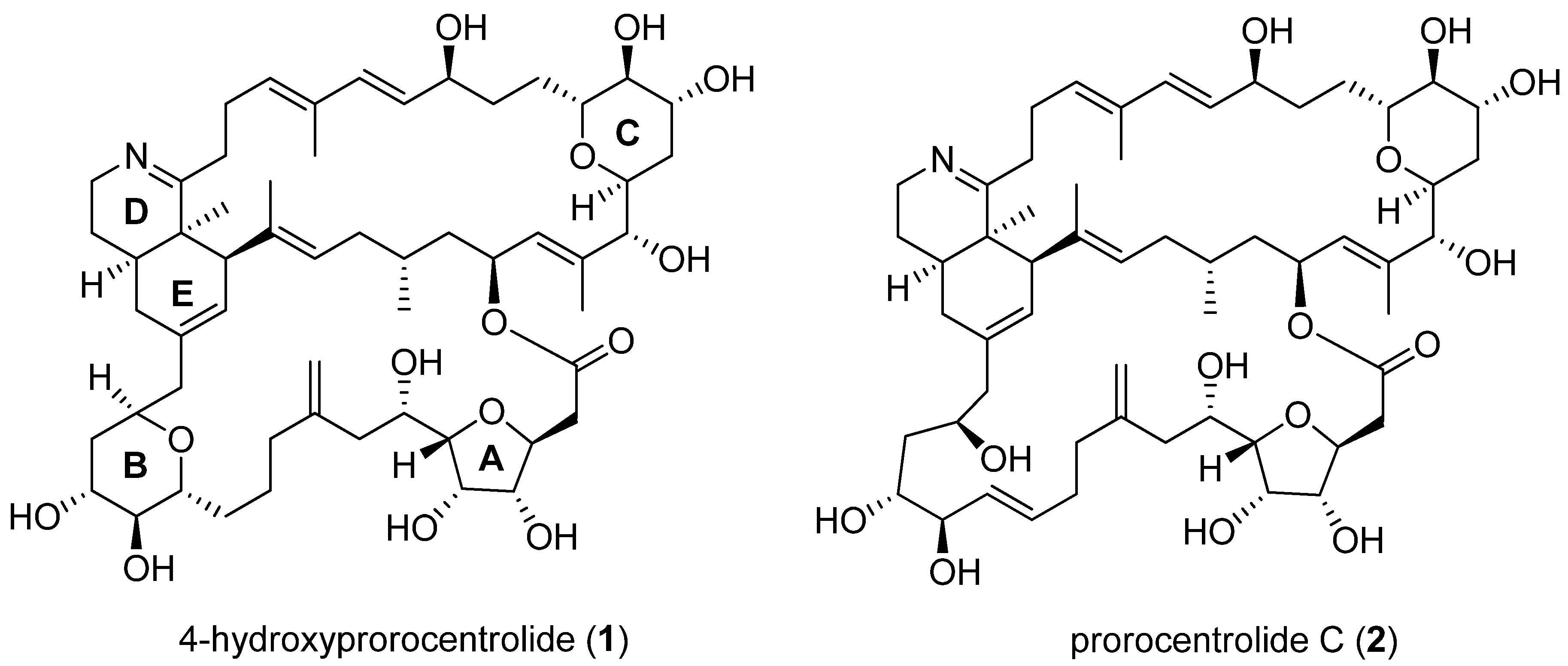
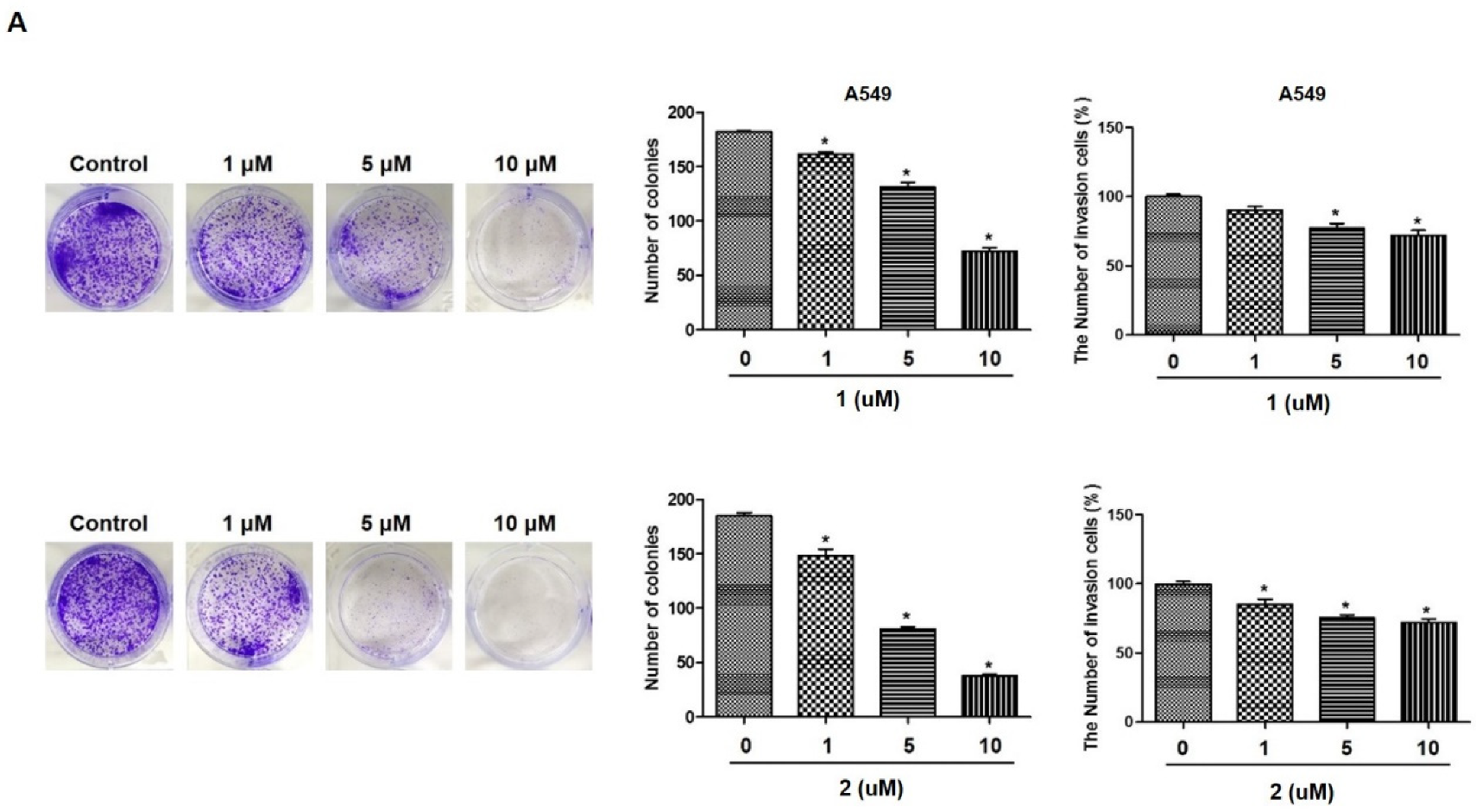
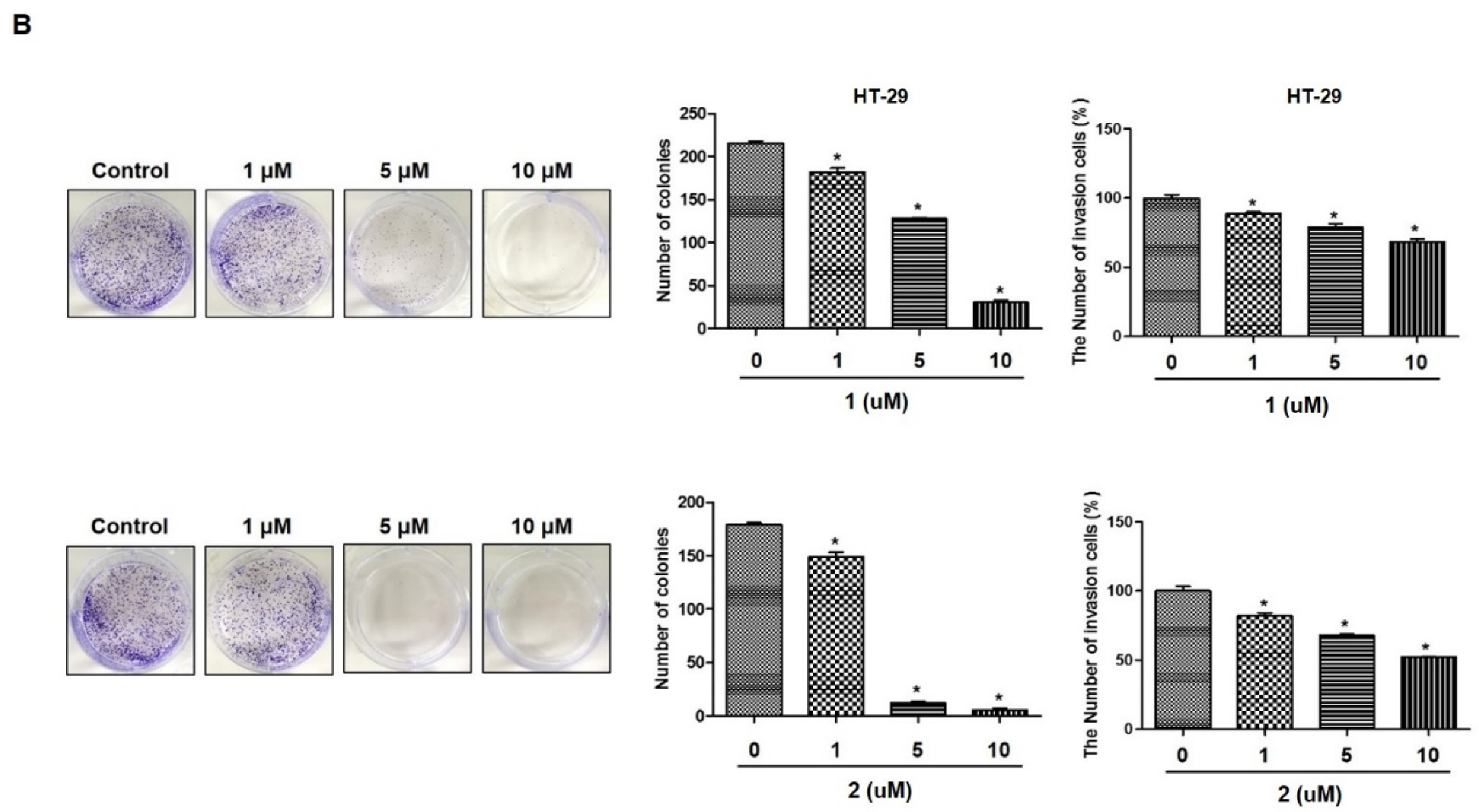
2.1. 4-Hydroxyprorocentrolide and Prorocentrolide C Inhibit the Proliferation of A549 and HT-29 Cancer Cells
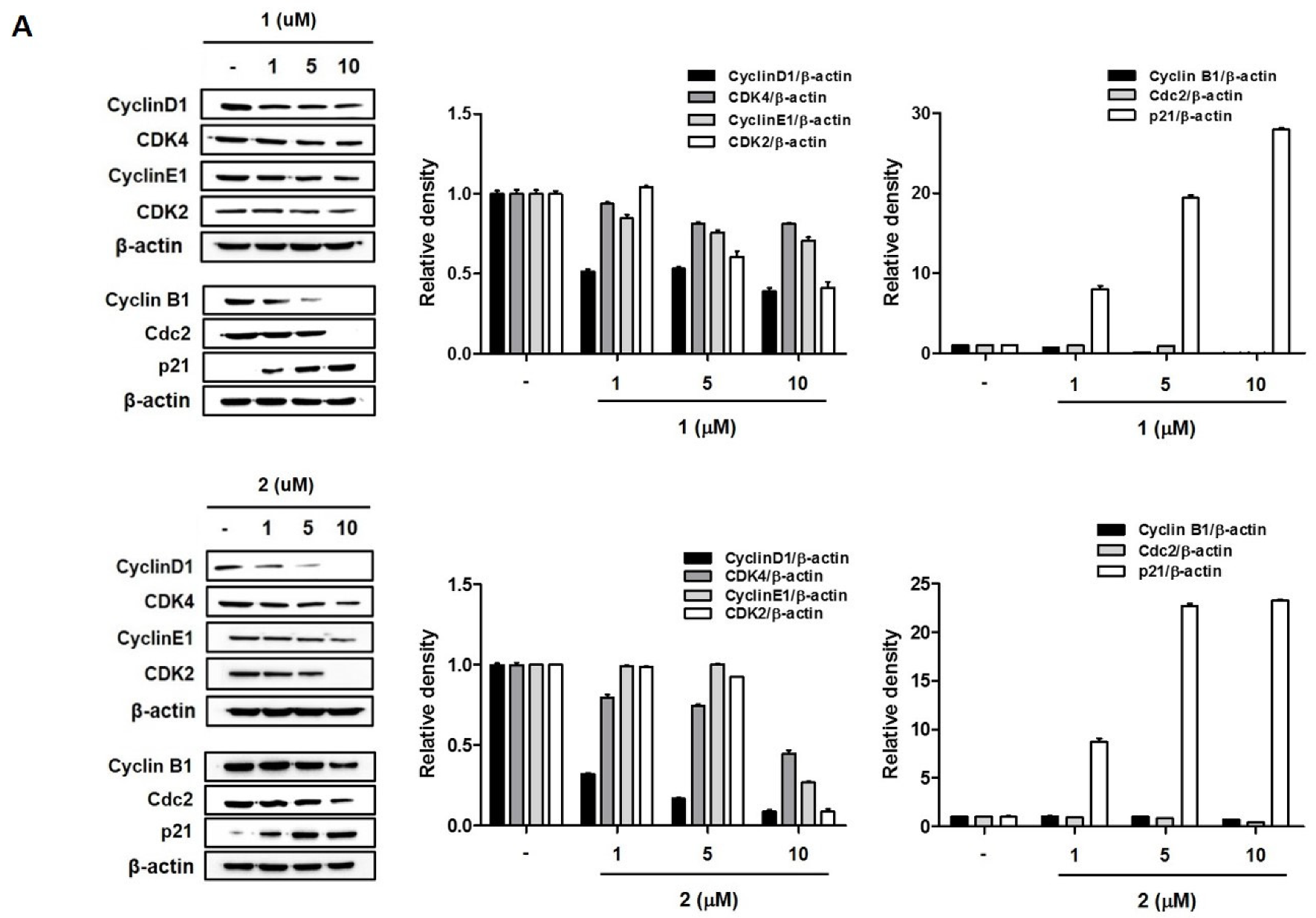
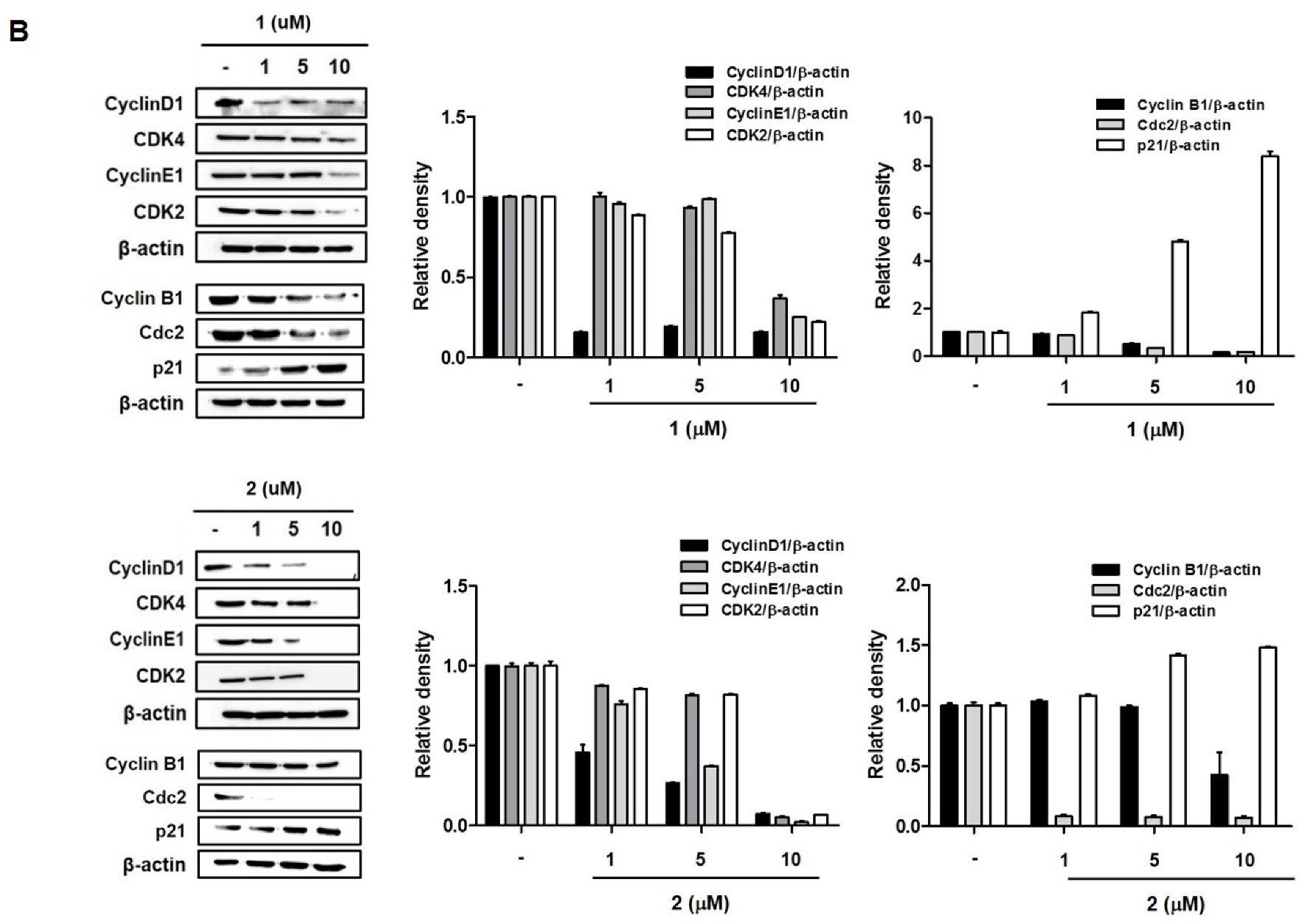
2.2. 4-Hydroxyprorocentrolide and Prorocentrolide C Induce S and G2/M Phase Arrest by Regulating Cell Cycle-Regulated Proteins
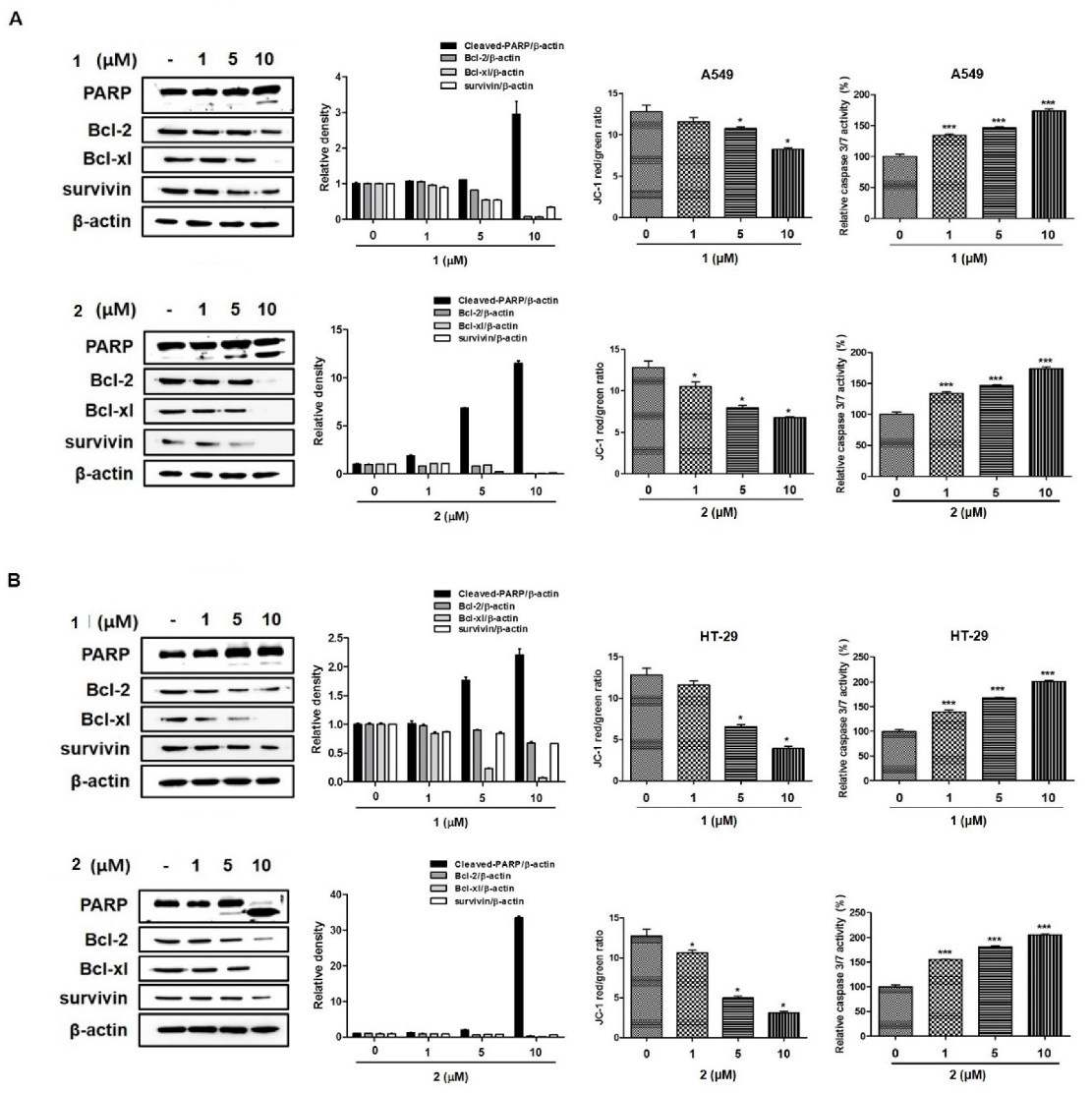
2.3. 4-Hydroxyprorocentrolide and Prorocentrolide C Induce Apoptosis in A549 and HT-29 Cancer Cells
2.4. 4-Hydroxyprorocentrolide and Prorocentrolide C Induce Caspase-Dependent Apoptosis in A549 and HT-29 Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Cultures
4.3. Cell Viability Assay
4.4. Colony Formation Assay
4.5. Apoptosis Analysis
4.6. Cell Cycle Analysis
4.7. Morphological Apoptosis
4.8. Mitochondrial Membrane Potential Assay
4.9. Transwell Invasion Assay
4.10. Caspase-3/7 Activity
4.11. Western Blot Analysis
Author Contributions
Funding
Conflicts of Interest
References
- American Cancer Society. Global Cancer Facts & Figures, 3rd ed.; American Cancer Society: Atlanta, GA, USA, 2015. [Google Scholar]
- American Cancer Society. Cancer Facts & Figures; American Cancer Society: Atlanta, GA, USA, 2016. [Google Scholar]
- Tubaro, A.; Sosa, S.; Bornancin, A.; Hungerford, J. Pharmacology and Toxicology of Diarrheic Shellfish Toxins. In Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection; CRC Press: Boca Raton, FL, USA, 2008; pp. 229–253. [Google Scholar]
- Gestal-Otero, J.J. Epidemiology of Marine Toxins. In Seafood and Freshwater Toxins: Pharmacology, Physiology, and Detection; Botana, L.M., Ed.; CRC Press: Boca Raton, FL, USA, 2014; p. 123. [Google Scholar]
- Bøe, R.; Gjertsen, B.T.; Vintermyr, O.K.; Houge, G.; Lanotte, M.; Døskeland, S.O. The Protein Phosphatase Inhibitor Okadaic Acid Induces Morphological Changes Typical of Apoptosis in Mammalian Cells. Exp. Cell Res. 1991, 195, 237–246. [Google Scholar] [CrossRef]
- Lerga, A.; Richard, C.; Delgado, M.D.; Cañelles, M.; Frade, P.; Cuadrado, M.A.; León, J. Apoptosis and Mitotic Arrest Are Two Independent Effects of the Protein Phosphatases Inhibitor Okadaic Acid in K562 Leukemia Cells. Biochem. Biophys. Res. Commun. 1999, 260, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Jayaraj, R.; Gupta, N.; Rao, P.V.L. Multiple Signal Transduction Pathways in Okadaic Acid Induced Apoptosis in hela Cells. Toxicology 2009, 256, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Fessard, V.; Grosse, Y.; Pfohl-Leszkowicz, A.; Puiseux-Dao, S. Okadaic Acid Treatment Induces DNA Adduct Formation in BHK21 C13 Fibroblasts and HESV Keratinocytes. Mutat. Res. 1996, 361, 133–141. [Google Scholar] [CrossRef]
- Traoré, A.; Baudrimont, I.; Ambaliou, S.; Dano, S.D.; Creppy, E.E. DNA Breaks and Cell Cycle Arrest Induced by Okadaic Acid in Caco-2 Cells, a Human Colonic Epithelial Cell Line. Arch. Toxicol. 2001, 75, 110–117. [Google Scholar] [CrossRef]
- Hégarat, L.L.; Orsière, T.; Botta, A.; Fessard, V. Okadaic Acid: Chromosomal Non-Disjunction Analysis in Human Lymphocytes and Study of Aneugenic Pathway in CHO-K1 Cells. Mutat. Res. 2005, 578, 53–63. [Google Scholar] [CrossRef]
- Tindall, D.R.; Dickey, R.W.; Carlson, R.D.; Morey-Gaines, G. Seafood Toxins. In ACS Symposium Series 262; Ragelis, E.P., Ed.; American Chemical Society: Washington, DC, USA, 1984; pp. 225–240. [Google Scholar]
- Torigoe, K.; Murata, M.; Yasumoto, T. Prorocentrolide, a Toxic Nitrogenous Macrocycle from a Marine Dinoflagellate, Prorocentrum Lima. J. Am. Chem. Soc. 1988, 110, 7876–7877. [Google Scholar] [CrossRef]
- Hu, T.M.; de Freitas, A.S.W.; Curtis, J.M.; Oshima, Y.; Walter, J.A.; Wright, J.L.C. Isolation and Structure of Prorocentrolide B, a Fast-Acting Toxin from Prorocentrum maculosum. J. Nat. Prod. 1996, 59, 1010–1014. [Google Scholar] [CrossRef]
- Krock, B.; Tillmann, U.; John, U.; Cembella, A. LCMS-MS aboard Ship: Tandem Mass Spectrometry in the Search for Phycotoxins and Novel Toxigenic Plankton from the North Sea. Anal. Bioanal. Chem. 2008, 392, 797–803. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Yang, A.R.; Yoo, Y.D.; Jeong, E.J.; Rho, J.R. Relative Configurational Assignment of 4-Hydroxyprorocentrolide and Prorocentrolide C Isolated from a Benthic Dinoflagellate (Prorocentrum lima). J. Nat. Prod. 2019, 82, 1034–1039. [Google Scholar] [CrossRef]
- Nagahama, Y.; Murray, S.; Tomaru, A.; Fukuyo, Y. Species Boundaries in the Toxic Dinoflagellate Prorocentrum Lima (Dinophyceae, Prorocentrales), Based on Morphological and Phylogenetic Characters (1). J. Phycol. 2011, 47, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Marr, J.; de Freitas, A.S.W.; Quilliam, M.A.; Walter, J.A.; Wright, J.L.C.; Pleasance, S. New Diol Esters of Okadaic Acid Isolated from Cultures of the Dinoflagellates Prorocentrum lima and P. Concavum. J. Nat. Prod. 1992, 55, 1631–1637. [Google Scholar] [CrossRef]
- Lee, T.C.; Fong, F.L.; Ho, K.C.; Lee, F.W. The Mechanism of Diarrhetic Shellfish Poisoning Toxin Production in Prorocentrum spp.: Physiological and Molecular Perspectives. Toxins 2016, 8, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sosa, S.; Tubaro, A. Okadaic Acid and Other Diarrheic Toxins: Toxicological Profile. In Marine and Freshwater Toxins; Gopalakrishnakone, P., Haddad, V., Jr., Tubaro, A., Kim, E., Kem, W.R., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 147–168. [Google Scholar]
- Yasumoto, T.; Murata, M.; Oshima, Y.; Sano, M.; Matsumoto, G.K.; Clardy, J. Diarrhetic Shellfish Toxins. Tetrahedron 1985, 41, 1019–1025. [Google Scholar] [CrossRef]
- Bialojan, C.; Takai, A. Inhibitory Effect of a Marine-Sponge Toxin, Okadaic Acid, on Protein Phosphatases. Specificity and Kinetics. Biochem. J. 1988, 256, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Takai, A.; Murata, M.; Torigoe, K.; Isobe, M.; Mieskes, G.; Yasumoto, T. Inhibitory Effect of Okadaic Acid Derivatives on Protein Phosphatases. A Study on Structure-Affinity Relationship. Biochem. J. 1992, 284, 539–544. [Google Scholar] [CrossRef]
- Le Hégarat, L.; Jacquin, A.G.; Bazin, E.; Fessard, V. Genotoxicity of the Marine Toxin Okadaic Acid, in Human Caco-2 Cells and in Mice Gut Cells. Environ. Toxicol. 2006, 21, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Rogers, C.G.; Héroux-Metcalf, C.; Langlois, I. Evaluation of Cytotoxicity and Genotoxicity of Okadaic Acid, a Nonphorbol Ester Type Tumor Promoter, in V79 Chinese Hamster Lung Cells. Toxicol. Vitro 1994, 8, 269–276. [Google Scholar] [CrossRef]
- Amar, M.; Aráoz, R.; Iorga, B.I.; Yasumoto, T.; Servent, D.; Molgó, J. Prorocentrolide-A from Cultured Prorocentrum lima Dinoflagellates Collected in Japan Blocks Sub-Types of Nicotinic Acetylcholine Receptors. Toxins 2018, 10, 97. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Yuan, J. Caspases in apoptosis and beyond. Oncogene 2008, 27, 6194–6206. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, S.H.; Desnoyers, S.; Ottaviano, Y.; Davidson, N.E.; Poirier, G.G. Specific proteolytic cleavage of poly(ADP-ribose) polymerase: An early marker of chemotherapy-induced apoptosis. Cancer Res. 1993, 53, 3976–3985. [Google Scholar] [PubMed]
- Nigg, E.A. Cyclin-Dependent Protein Kinases: Key Regulators of the Eukaryotic Cell Cycle. Bioessays 1995, 17, 471–480. [Google Scholar] [CrossRef]
- Bunz, F.; Dutriaux, A.; Lengauer, C.; Waldman, T.; Zhou, S.; Brown, J.P.; Sedivy, J.M.; Kinzler, K.W.; Vogelstein, B. Requirement for p53 and p21 to Sustain G2 Arrest After DNA Damage. Science 1998, 282, 1497–1501. [Google Scholar] [CrossRef] [PubMed]
- Taylor, W.R.; Stark, G.R. Regulation of the G2/M Transition by p53. Oncogene 2001, 20, 1803–1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.M.; Kim, N.-H.; Jeong, E.J.; Rho, J.-R. Cytotoxic 4-Hydroxyprorocentrolide and Prorocentrolide C from Cultured Dinoflagellate Prorocentrum lima Induce Human Cancer Cell Death through Apoptosis and Cell Cycle Arrest. Toxins 2020, 12, 304. https://doi.org/10.3390/toxins12050304
Lee SM, Kim N-H, Jeong EJ, Rho J-R. Cytotoxic 4-Hydroxyprorocentrolide and Prorocentrolide C from Cultured Dinoflagellate Prorocentrum lima Induce Human Cancer Cell Death through Apoptosis and Cell Cycle Arrest. Toxins. 2020; 12(5):304. https://doi.org/10.3390/toxins12050304
Chicago/Turabian StyleLee, Seon Min, Na-Hyun Kim, Eun Ju Jeong, and Jung-Rae Rho. 2020. "Cytotoxic 4-Hydroxyprorocentrolide and Prorocentrolide C from Cultured Dinoflagellate Prorocentrum lima Induce Human Cancer Cell Death through Apoptosis and Cell Cycle Arrest" Toxins 12, no. 5: 304. https://doi.org/10.3390/toxins12050304
APA StyleLee, S. M., Kim, N.-H., Jeong, E. J., & Rho, J.-R. (2020). Cytotoxic 4-Hydroxyprorocentrolide and Prorocentrolide C from Cultured Dinoflagellate Prorocentrum lima Induce Human Cancer Cell Death through Apoptosis and Cell Cycle Arrest. Toxins, 12(5), 304. https://doi.org/10.3390/toxins12050304