Delayed Release of Intracellular Microcystin Following Partial Oxidation of Cultured and Naturally Occurring Cyanobacteria

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
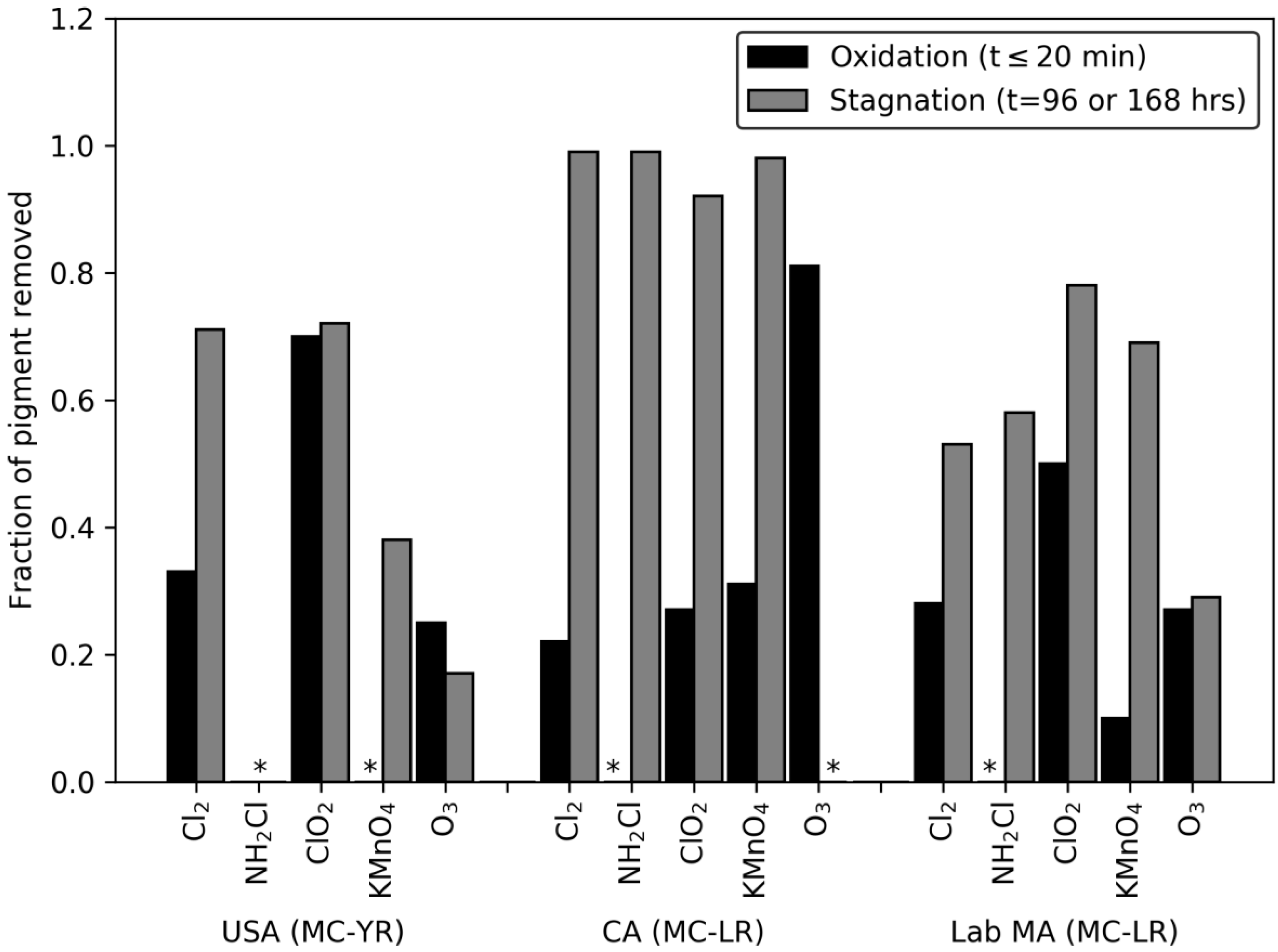
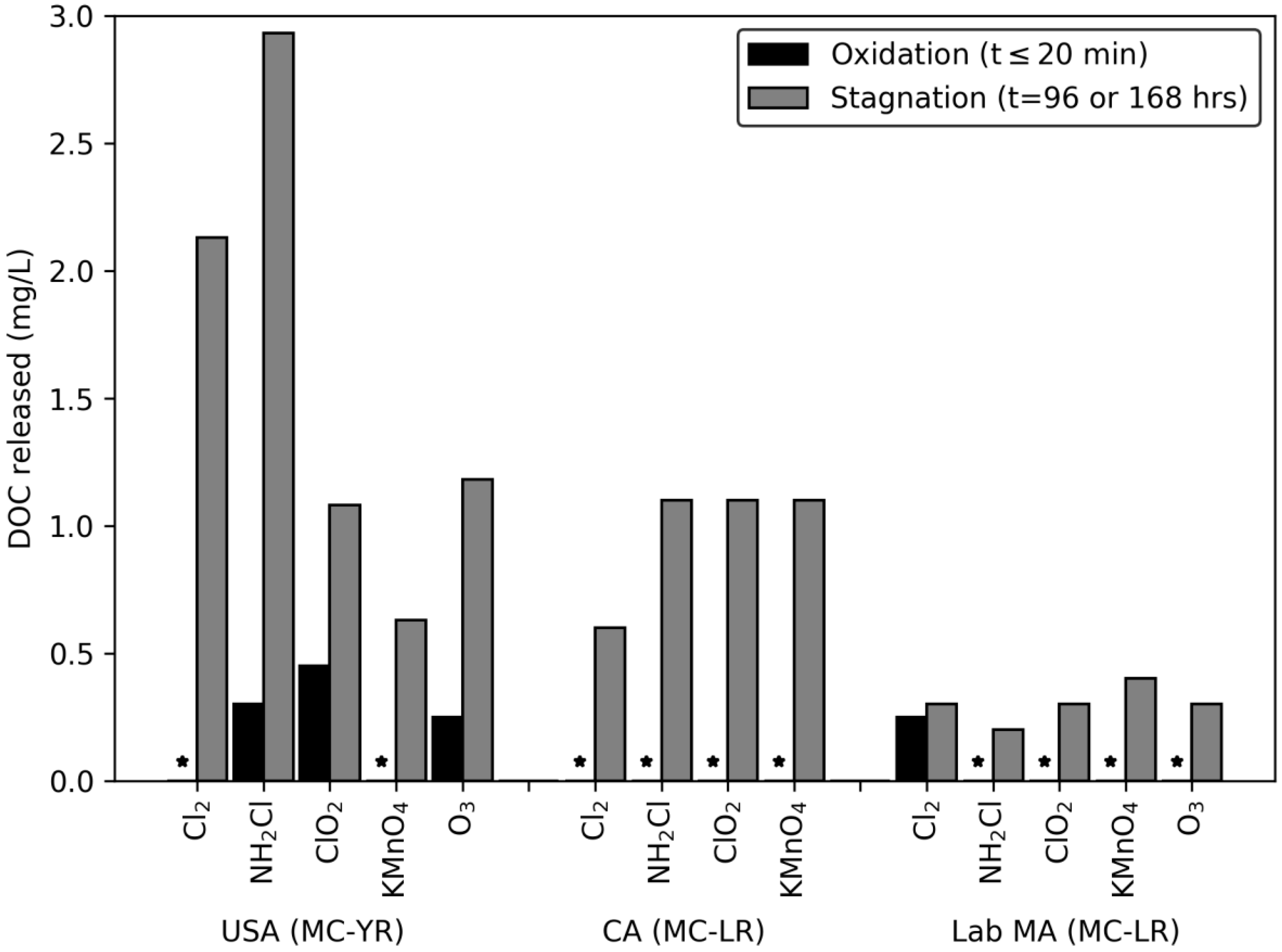
2.1. Oxidation (Time ≤ 20 min) Induced Degradation of Pigment and Release of DOC
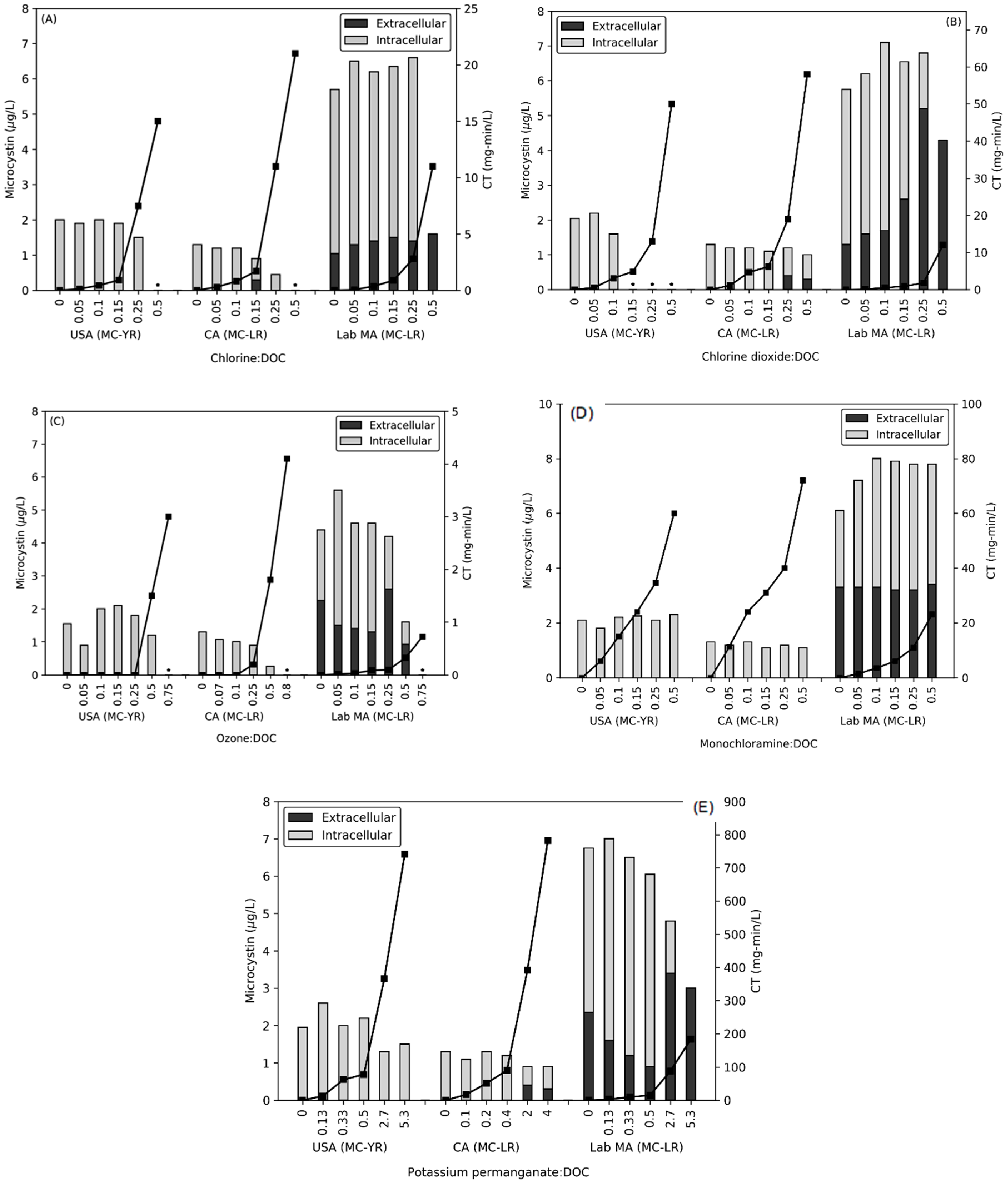
2.2. Impact of Oxidants on Release of Microcystin (t ≤ 20 min)
2.3. Impact of Stagnation (Time Max = 96 or 168 h) on Pigment and DOC
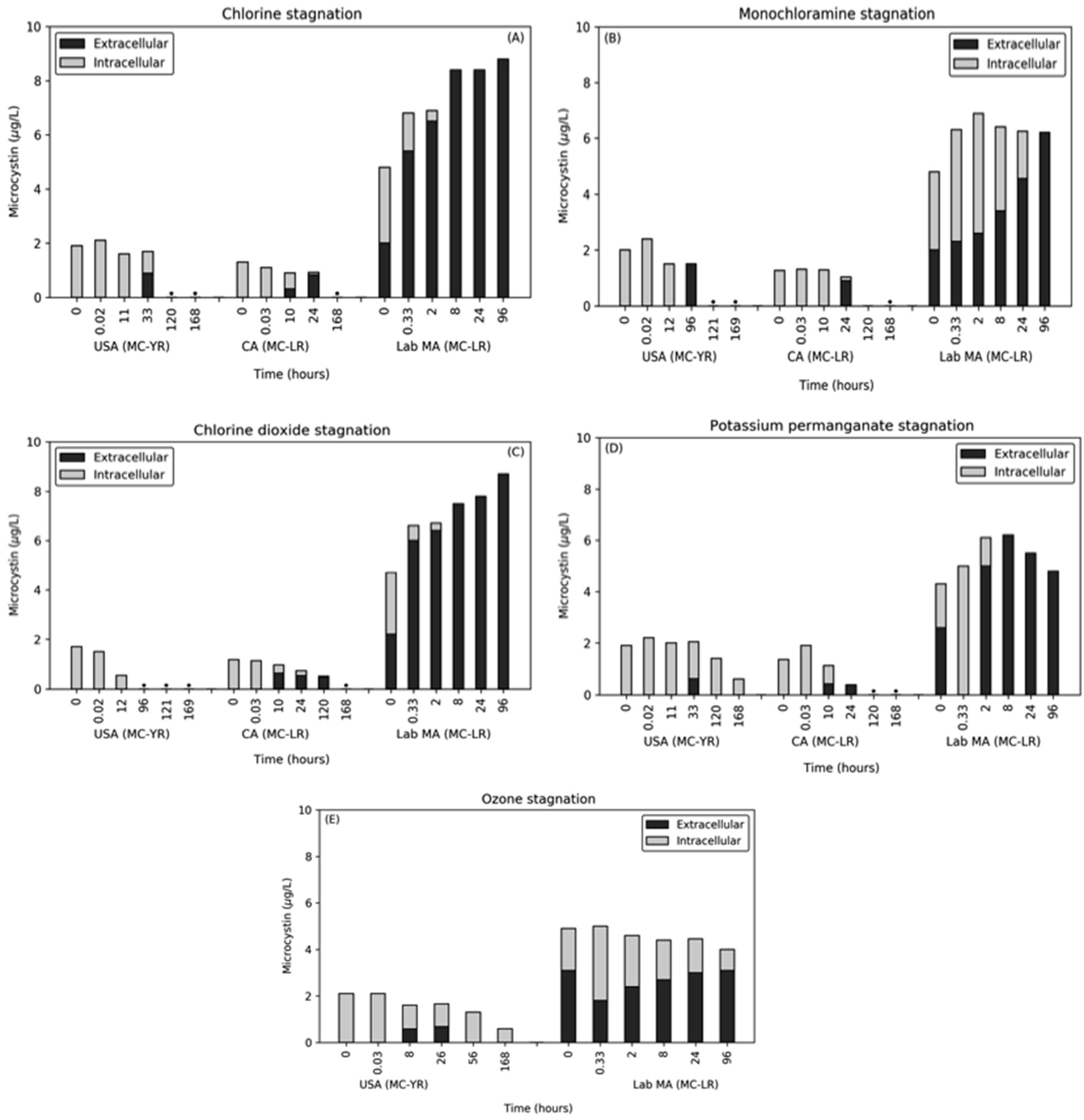
2.4. Impact of Stagnation (Time Max = 96 or 168 h) on Microcystin
3. Conclusions
4. Materials and Methods
4.1. Cyanobacteria Culturing, Sampling, and Characterization
4.1.1. Laboratory-Cultured Microcystis aeruginosa Cells Transferred into Colorado River Water (CRW)
4.1.2. United States (USA) Bloom: Grand Lake St. Marys
4.1.3. Canadian (CA) Bloom: Lake Champlain
4.2. Pre-Oxidation of Cyanobacteria Suspensions
4.2.1. Varied Oxidant: DOC Ratios
4.2.2. Varied Stagnation Times
4.3. Sample Analyses
4.3.1. Cyanobacteria Bloom Characterization and Water Quality Parameters
4.3.2. Oxidant Residuals
4.3.3. Microcystin
4.3.4. Calculation of Oxidant Decay, MC Decay or Release Rates, and Cell Damage Rates
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- USEPA Drinking Water Health Advisory for the Cyanobacterial Microcystin Toxins: 820R15100. Available online: https://www.epa.gov/sites/production/files/2017-06/documents/microcystins-report-2015.pdf (accessed on 18 November 2018).
- USEPA Drinking Water Health Advisory for the Cyanobacterial Toxin Cylindrospermopsin: 820R15101. Available online: https://www.epa.gov/sites/production/files/2017-06/documents/cylindrospermopsin-report-2015.pdf (accessed on 18 November 2018).
- Health Canada Guidelines for Canadian Drinking Water Quality Cyanobacterial Toxins. Available online: https://www.canada.ca/en/health-canada/services/publications/healthy-living/guidelines-canadian-drinking-water-quality-guideline-technical-document-cyanobacterial-toxins-document.html (accessed on 28 December 2018).
- American Water Works Association (AWWA) Cyanotoxins in US Drinking Water: Occurrence, Case Studies and State Approaches to Regulation. Available online: https://www.awwa.org/Portals/0/AWWA/Government/201609_Cyanotoxin_Occurrence_States_Approach.pdf?ver=2018-12-13-101832-037 (accessed on 18 November 2018).
- USEPA Recommendations for Public Water Systems to Manage Cyanotoxins in Drinking Water: 815R15100. Available online: https://www.epa.gov/sites/production/files/2017-06/documents/cyanotoxin-management-drinking-water.pdf (accessed on 18 November 2018).
- USEPA. Cyanobacteria and Cyanotoxins: Information for Drinking Water Systems; EPA-810F11; USEPA: Washington, DC, USA, 2019.
- American Water Works Association (AWWA); Water Research Foundation (WRF) A Water Utility Manager’s Guide to Cyanotoxins. Available online: https://www.awwa.org/Portals/0/AWWA/Government/WaterUtilityManagersGuideToCyanotoxins.pdf?ver=2018-12-13-101839-130 (accessed on 28 November 2018).
- Global Water Research Coalition (GWRC); Water Quality Research Australia (WQRA). International Guidance Manual for the Management of Toxic Cyanobacteria; Newcombe, G., Ed.; Alliance House: London, UK, 2009; ISBN 978-90-77622-21-6. [Google Scholar]
- Newcombe, G. International Guidance Manual for the Management of Toxic Cyanobacteria. Available online: https://www.iwapublishing.com/books/9781780401355/international-guidance-manual-management-toxic-cyanobacteria (accessed on 27 April 2020).
- Crittenden, J.C.; Trussell, R.R.; Hand, D.W.; Howe, K.J.; Tchobanoglous, G. MWH’s Water Treatment: Principles and Design: Third Edition; John Wiley and Sons: Hoboken, NJ, USA, 2012; ISBN 9780470405390. [Google Scholar]
- Oregon Department of Human Services: Public Health Division Fast Facts: Blue-green Alage Q&A for Water System Operators. Available online: https://www.oregon.gov/oha/PH/HEALTHYENVIRONMENTS/RECREATION/HARMFULALGAEBLOOMS/Pages/resources_for_samplers.aspx (accessed on 28 December 2018).
- Zamyadi, A.; Fan, Y.; Daly, R.I.; Prévost, M. Chlorination of Microcystis aeruginosa: Toxin release and oxidation, cellular chlorine demand and disinfection by-products formation. Water Res. 2013, 47, 1080–1090. [Google Scholar] [CrossRef]
- Wert, E.C.; Korak, J.A.; Trenholm, R.A.; Rosario-Ortiz, F.L. Effect of oxidant exposure on the release of intracellular microcystin, MIB, and geosmin from three cyanobacteria species. Water Res. 2014, 52, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Daly, R.I.; Ho, L.; Brookes, J.D. Effect of chlorination on Microcystis aeruginosa cell integrity and subsequent microcystin release and degradation. Environ. Sci. Technol. 2007, 41, 4447–4453. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Shi, H.; Timmons, T.; Adams, C. Release and removal of microcystins from Microcystis during oxidative-, physical-, and UV-Based disinfection. J. Environ. Eng. 2010, 136, 2–11. [Google Scholar] [CrossRef]
- He, X.; Wert, E.C. Colonial cell disaggregation and intracellular microcystin release following chlorination of naturally occurring Microcystis. Water Res. 2016, 101. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Dan, Y.; Adams, C.D.; Shi, H.; Ma, Y.; Eichholz, T. Effect of oxidant demand on the release and degradation of microcystin-LR from Microcystis aeruginosa during oxidation. Chemosphere 2017, 181, 562–568. [Google Scholar] [CrossRef]
- Zamyadi, A.; Greenstein, K.E.; Glover, C.M.; Adams, C.; Rosenfeldt, E.; Wert, E.C. Impact of hydrogen peroxide and copper sulfate on the delayed release of microcystin. Water 2020, 12, 1105. [Google Scholar] [CrossRef]
- Moradinejad, S.; Glover, C.M.; Mailly, J.; Seighalani, T.Z.; Peldszus, S.; Barbeau, B.; Dorner, S.; Prévost, M.; Zamyadi, A. Using advanced spectroscopy and organic matter characterization to evaluate the impact of oxidation on cyanobacteria. Toxins 2019, 11, 278. [Google Scholar] [CrossRef] [Green Version]
- Zamyadi, A.; Coral, L.A.; Barbeau, B.; Dorner, S.; Lapolli, F.R.; Prévost, M. Fate of toxic cyanobacterial genera from natural bloom events during ozonation. Water Res. 2015, 73, 204–215. [Google Scholar] [CrossRef]
- Coral, L.A.; Zamyadi, A.; Barbeau, B.; Bassetti, F.J.; Lapolli, F.R.; Prévost, M. Oxidation of Microcystis aeruginosa and Anabaena flos-aquae by ozone: Impacts on cell integrity and chlorination by-product formation. Water Res. 2013, 47, 2983–2994. [Google Scholar] [CrossRef]
- Zamyadi, A.; Ho, L.; Newcombe, G.; Bustamante, H.; Prévost, M. Fate of toxic cyanobacterial cells and disinfection by-products formation after chlorination. Water Res. 2012, 46, 1524–1535. [Google Scholar] [CrossRef] [PubMed]
- Wert, E.C.; Dong, M.M.; Rosario-Ortiz, F.L. Using digital flow cytometry to assess the degradation of three cyanobacteria species after oxidation processes. Water Res. 2013, 47, 3752–3761. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhu, C.; Xie, C.; Shao, C.; Yu, S.; Zhao, L.; Gao, N. Kinetics and mechanism of Pseudoanabaena cell inactivation, 2-MIB release and degradation under exposure of ozone, chlorine and permanganate. Water Res. 2018, 147, 422–428. [Google Scholar] [CrossRef]
- Zamyadi, A.; Choo, F.; Newcombe, G.; Stuetz, R.; Henderson, R.K. A review of monitoring technologies for real-time management of cyanobacteria: Recent advances and future direction. TrAC Trends Anal. Chem. 2016, 85, 83–96. [Google Scholar] [CrossRef]
- Zhou, S.; Shao, Y.; Gao, N.; Li, L.; Deng, J.; Zhu, M.; Zhu, S. Effect of chlorine dioxide on cyanobacterial cell integrity, toxin degradation and disinfection by-product formation. Sci. Total Environ. 2014, 482–483, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Laszakovits, J.R.; MacKay, A.A. Removal of cyanotoxins by potassium permanganate: Incorporating competition from natural water constituents. Water Res. 2019, 155, 86–95. [Google Scholar] [CrossRef]
- Ramseier, M.K.; von Gunten, U.; Freihofer, P.; Hammes, F. Kinetics of membrane damage to high (HNA) and low (LNA) nucleic acid bacterial clusters in drinking water by ozone, chlorine, chlorine dioxide, monochloramine, ferrate(VI), and permanganate. Water Res. 2011, 45, 1490–1500. [Google Scholar] [CrossRef]
- Rodríguez, E.; Onstad, G.D.; Kull, T.P.J.; Metcalf, J.S.; Acero, J.L.; von Gunten, U. Oxidative elimination of cyanotoxins: Comparison of ozone, chlorine, chlorine dioxide and permanganate. Water Res. 2007, 41, 3381–3393. [Google Scholar] [CrossRef]
- Fan, J.; Ho, L.; Hobson, P.; Daly, R.; Brookes, J. Application of various oxidants for cyanobacteria control and cyanotoxin removal in wastewater treatment. J. Environ. Eng. 2014, 140, 04014022. [Google Scholar] [CrossRef]
- Onstad, G.D.; Strauch, S.; Meriluoto, J.; Codd, G.A.; Von Gunten, U. Selective oxidation of key functional groups in cyanotoxins during drinking water ozonation. Environ. Sci. Technol. 2007, 41, 4397–4404. [Google Scholar] [CrossRef]
- Acero, J.L.; Rodriguez, E.; Meriluoto, J. Kinetics of reactions between chlorine and the cyanobacterial toxins microcystins. Water Res. 2005, 39, 1628–1638. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Stanford, B.D.; Adams, C.; Rosenfeldt, E.J.; Wert, E.C. Varied influence of microcystin structural difference on ELISA cross-reactivity and chlorination efficiency of congener mixtures. Water Res. 2017, 126, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.; Onstad, G.; Von Gunten, U.; Rinck-Pfeiffer, S.; Craig, K.; Newcombe, G. Differences in the chlorine reactivity of four microcystin analogues. Water Res. 2006, 40, 1200–1209. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; von Gunten, U. Oxidative transformation of micropollutants during municipal wastewater treatment: Comparison of kinetic aspects of selective (chlorine, chlorine dioxide, ferrateVI, and ozone) and non-selective oxidants (hydroxyl radical). Water Res. 2010, 44, 555–566. [Google Scholar] [CrossRef]
- Pestana, C.J.; Reeve, P.J.; Sawade, E.; Voldoire, C.F.; Newton, K.; Praptiwi, R.; Collingnon, L.; Dreyfus, J.; Hobson, P.; Gaget, V.; et al. Fate of cyanobacteria in drinking water treatment plant lagoon supernatant and sludge. Sci. Total Environ. 2016, 565, 1192–1200. [Google Scholar] [CrossRef]
- Zamyadi, A.; MacLeod, S.L.; Fan, Y.; McQuaid, N.; Dorner, S.; Sauvé, S.; Prévost, M. Toxic cyanobacterial breakthrough and accumulation in a drinking water plant: A monitoring and treatment challenge. Water Res. 2012, 46, 1511–1523. [Google Scholar] [CrossRef]
- Zamyadi, A.; Romanis, C.; Mills, T.; Neilan, B.; Choo, F.; Coral, L.A.; Gale, D.; Newcombe, G.; Crosbie, N.; Stuetz, R.; et al. Diagnosing water treatment critical control points for cyanobacterial removal: Exploring benefits of combined microscopy, next-generation sequencing, and cell integrity methods. Water Res. 2019, 152, 96–105. [Google Scholar] [CrossRef]
- Chen, W.; Song, L.; Peng, L.; Wan, N.; Zhang, X.; Gan, N. Reduction in microcystin concentrations in large and shallow lakes: Water and sediment-interface contributions. Water Res. 2008, 42, 763–773. [Google Scholar] [CrossRef]
- Maghsoudi, E.; Fortin, N.; Greer, C.; Duy, S.V.; Fayad, P.; Sauvé, S.; Prévost, M.; Dorner, S. Biodegradation of multiple microcystins and cylindrospermopsin in clarifier sludge and a drinking water source: Effects of particulate attached bacteria and phycocyanin. Ecotoxicol. Environ. Saf. 2015, 120, 409–417. [Google Scholar] [CrossRef]
- Wert, E.C.; Rosario-Ortiz, F.L. Intracellular organic matter from cyanobacteria as a precursor for carbonaceous and nitrogenous disinfection byproducts. Environ. Sci. Technol. 2013, 47. [Google Scholar] [CrossRef]
- Dumouchelle, D.H.; Stelzer, E.A. Chemical and biological quality of water in Grand Lake St. Marys, Ohio, 2011–12, with emphasis on cyanobacteria. U.S. Geol. Surv. Sci. Investig. Rep. 2014, 5210. [Google Scholar] [CrossRef] [Green Version]
- Lyon, B.A.; Milsk, R.Y.; Deangelo, A.B.; Simmons, J.E.; Moyer, M.P.; Weinberg, H.S. Integrated chemical and toxicological investigation of UV-chlorine/chloramine drinking water treatment. Environ. Sci. Technol. 2014, 48, 6743–6753. [Google Scholar] [CrossRef] [PubMed]
- Almuhtaram, H.; Cui, Y.; Zamyadi, A.; Hofmann, R. Cyanotoxins and cyanobacteria cell accumulations in drinking water treatment plants with a low risk of bloom formation at the source. Toxins 2018, 10, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- APHA; AWWA; WEF. Standard Methods for Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 2012. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oxidant | Water | kdamage (M−1s−1) (R2) | ktotal (M−1s−1) (R2) |
|---|---|---|---|
| Cl2 | USA | 25 (0.96) | 74.6 (0.91) |
| CA | 133 * (0.98) | 82.9 (0.96) | |
| ClO2 | USA | 64 (0.76) | - |
| CA | 20.9 * (0.84) | 11.5 (0.82) | |
| KMnO4 | USA | - | - |
| CA | 2.69 * (0.84) | 1.22 (0.73) | |
| Ozone | USA | 273 (0.91) | 143 (0.99) |
| CA | 245 * (0.75) | 56 (0.75) |
| Cl2 | NH2Cl | ClO2 | O3 | KMnO4 | |
|---|---|---|---|---|---|
| Oxidant:DOC ratio (t ≤ 20 min) | 0.5 | No release | 0.5 * | 0.75/0.80 | >2 * |
| CTlab (mg-min/L) | 11 | 23 | 12 | 0.72 | 117 |
| CTUSA (mg-min/L) | 15 | 60 | 50 | 3.0 | 486 |
| CTCA (mg-min/L) | 21 | 72 | 58 | 4.1 | 391 |
| Stagnation time * | ≥2 h | ≥8 h | ≥20 min | ≥8 h | ≥2 h |
| Oxidant | Water | Stagnation (Time Max = 96 or 168 h) | |
|---|---|---|---|
| kbiodegradation (R2)/*krelease (s−1) (R2) | Half-Life (Days) | ||
| Chlorine | USA | 2.57 × 10−6 (0.93) | 3.12 |
| CA | 3.37 × 10−6 (0.99) | 2.38 | |
| Lab MA | *1.61 × 10−6 (0.67) | - | |
| Mono-chloramine | USA | 2.44 × 10−6 (0.88) | 3.29 |
| CA | 4.08 × 10−6 (0.95) | 1.97 | |
| Lab MA | *4.12 × 10−6 (0.79) | - | |
| Chlorine dioxide | USA | - | - |
| CA | 2.85 × 10−6 (0.90) | 2.81 | |
| Lab MA | *2.79 × 10−6 (0.83) | - | |
| KMnO4 | USA | 1.79 × 10−6 (0.83) | 4.48 |
| CA | 3.93 × 10−6 (0.81) | 2.04 | |
| Lab MA | - | - | |
| Ozone | USA | 1.90 × 10−6 (0.90) | 4.22 |
| Lab MA | 7.44 × 10−7 (0.76)/*2.50 × 10−6 (0.84) | 10.8 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Greenstein, K.E.; Zamyadi, A.; Glover, C.M.; Adams, C.; Rosenfeldt, E.; Wert, E.C. Delayed Release of Intracellular Microcystin Following Partial Oxidation of Cultured and Naturally Occurring Cyanobacteria. Toxins 2020, 12, 335. https://doi.org/10.3390/toxins12050335
Greenstein KE, Zamyadi A, Glover CM, Adams C, Rosenfeldt E, Wert EC. Delayed Release of Intracellular Microcystin Following Partial Oxidation of Cultured and Naturally Occurring Cyanobacteria. Toxins. 2020; 12(5):335. https://doi.org/10.3390/toxins12050335
Chicago/Turabian StyleGreenstein, Katherine E., Arash Zamyadi, Caitlin M. Glover, Craig Adams, Erik Rosenfeldt, and Eric C. Wert. 2020. "Delayed Release of Intracellular Microcystin Following Partial Oxidation of Cultured and Naturally Occurring Cyanobacteria" Toxins 12, no. 5: 335. https://doi.org/10.3390/toxins12050335
APA StyleGreenstein, K. E., Zamyadi, A., Glover, C. M., Adams, C., Rosenfeldt, E., & Wert, E. C. (2020). Delayed Release of Intracellular Microcystin Following Partial Oxidation of Cultured and Naturally Occurring Cyanobacteria. Toxins, 12(5), 335. https://doi.org/10.3390/toxins12050335