Characterization of Bacillus cereus in Dairy Products in China



Abstract
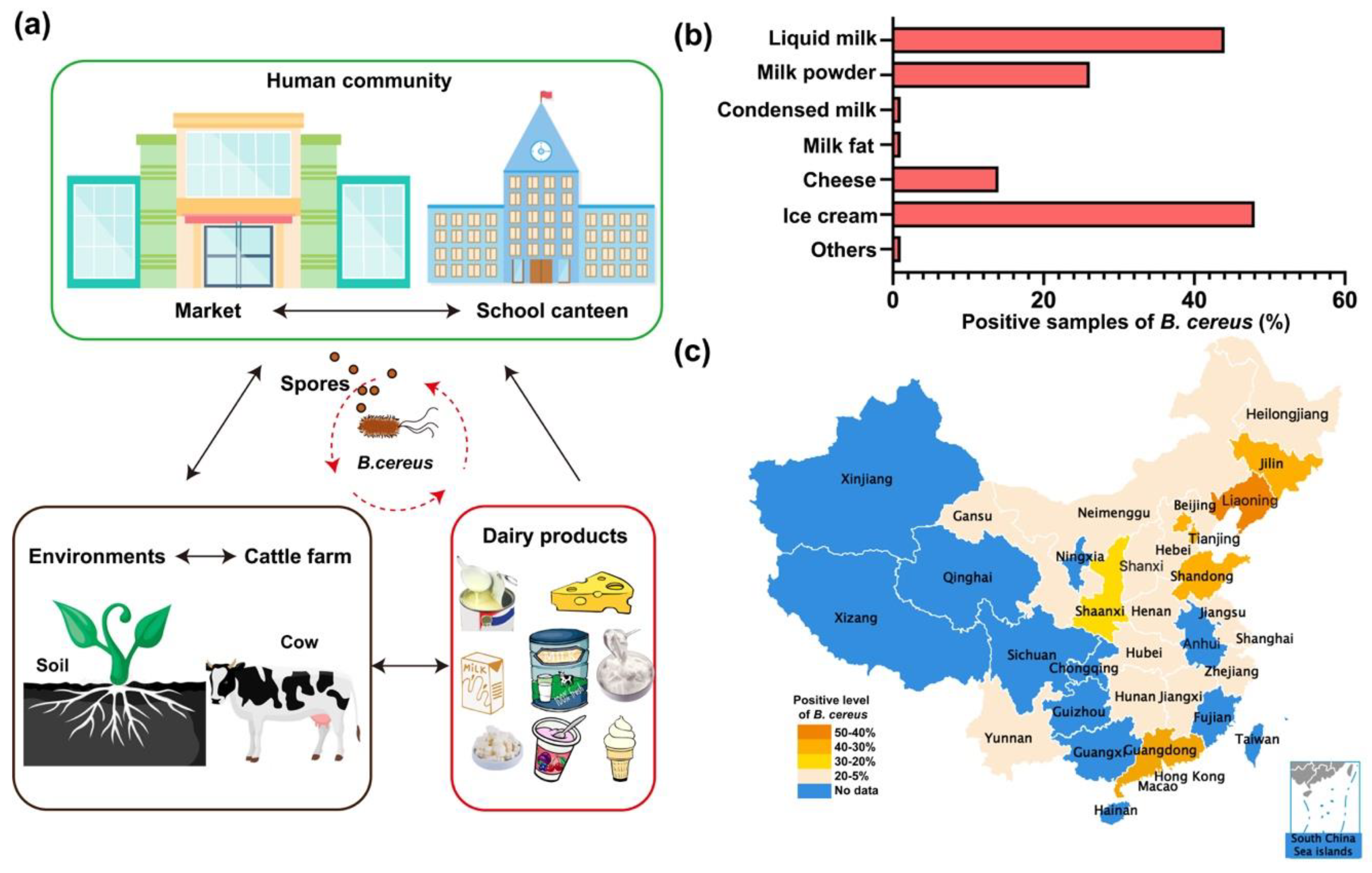
:1. Introduction
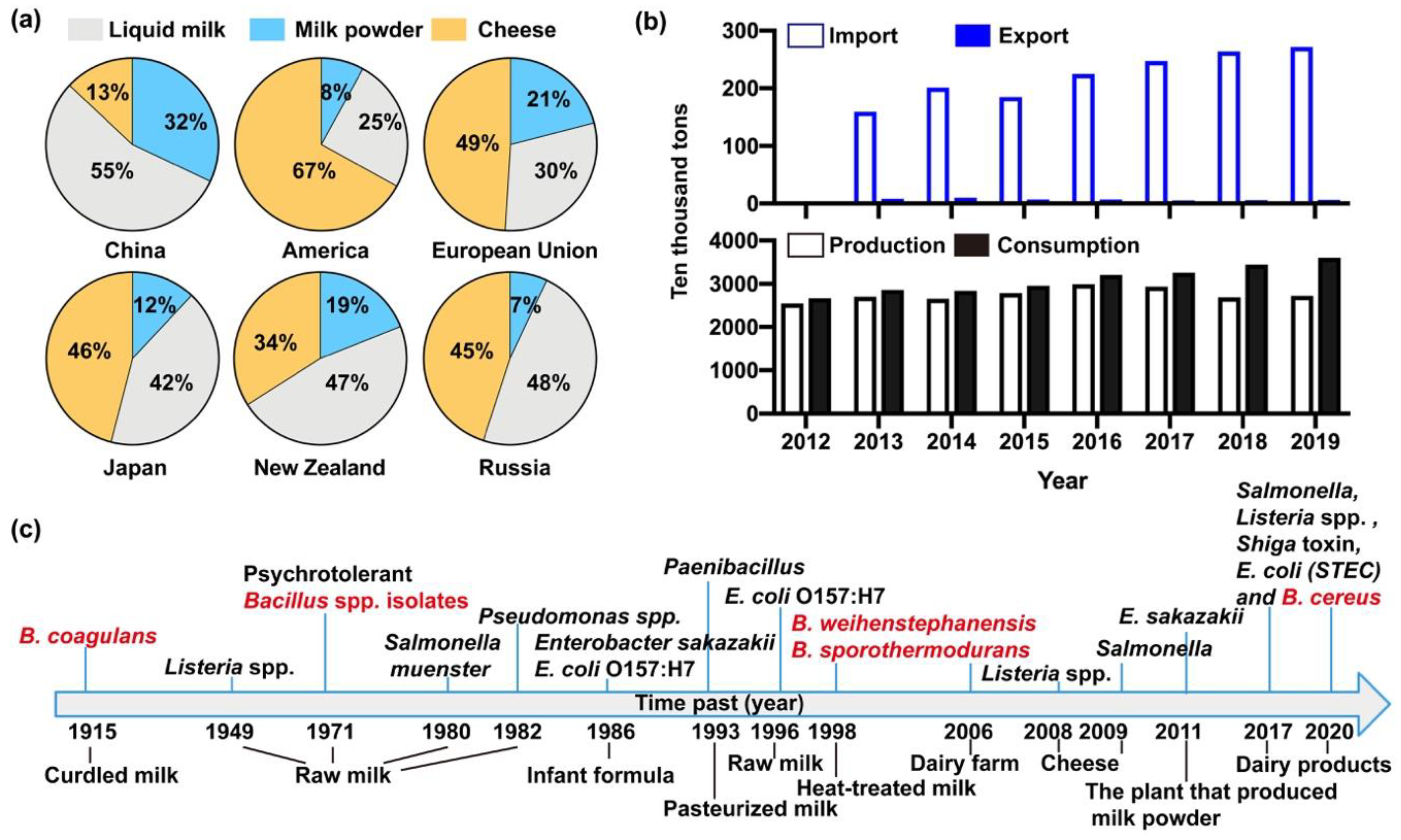
2. Dairy Products with Bacillus cereus
3. Prevalence of Bacillus cereus from Dairy Products in China
3.1. Contamination of Bacillus cereus Isolates
3.2. Distribution of Bacillus cereus in Milk and Milk Products in China
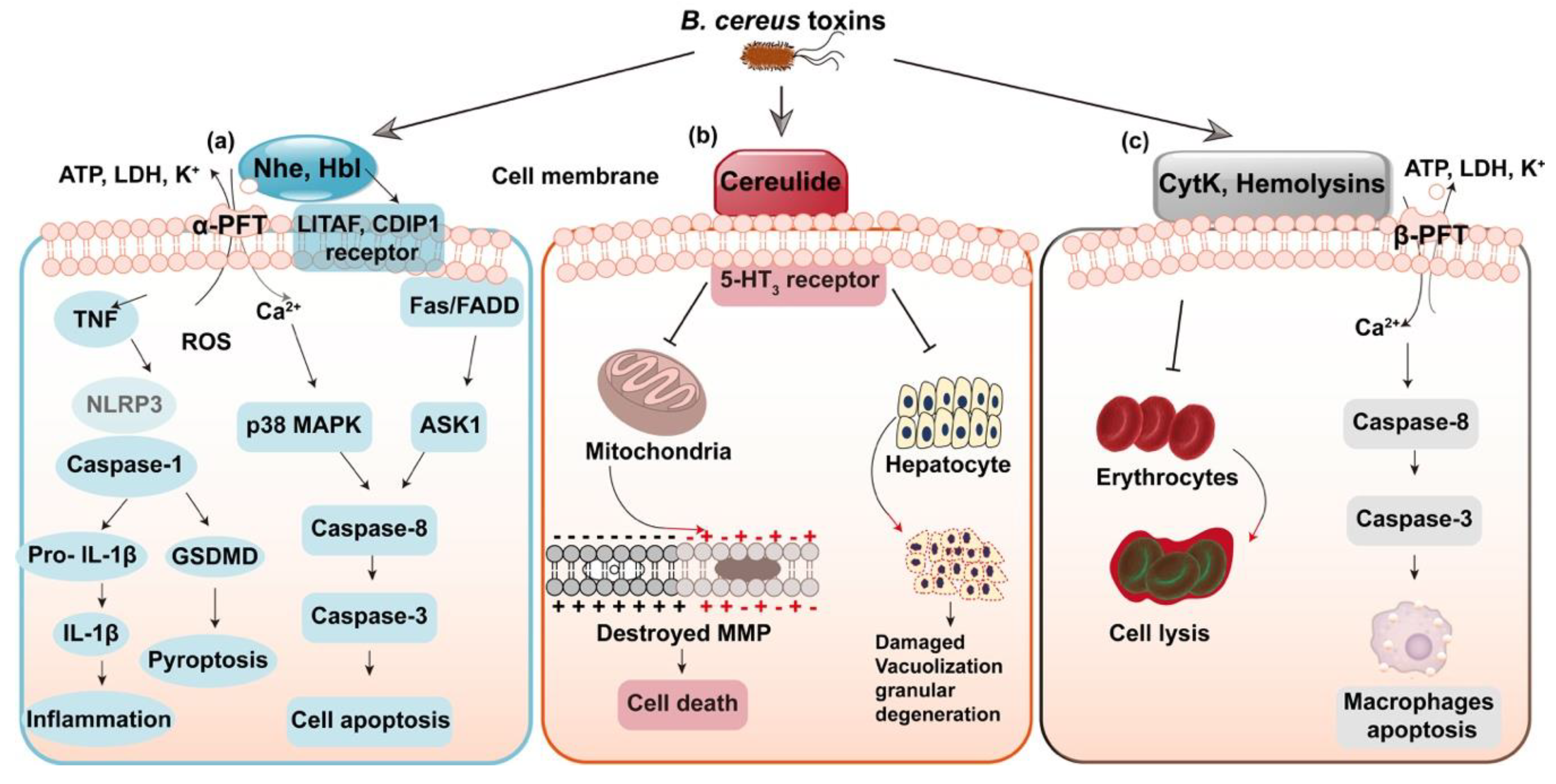
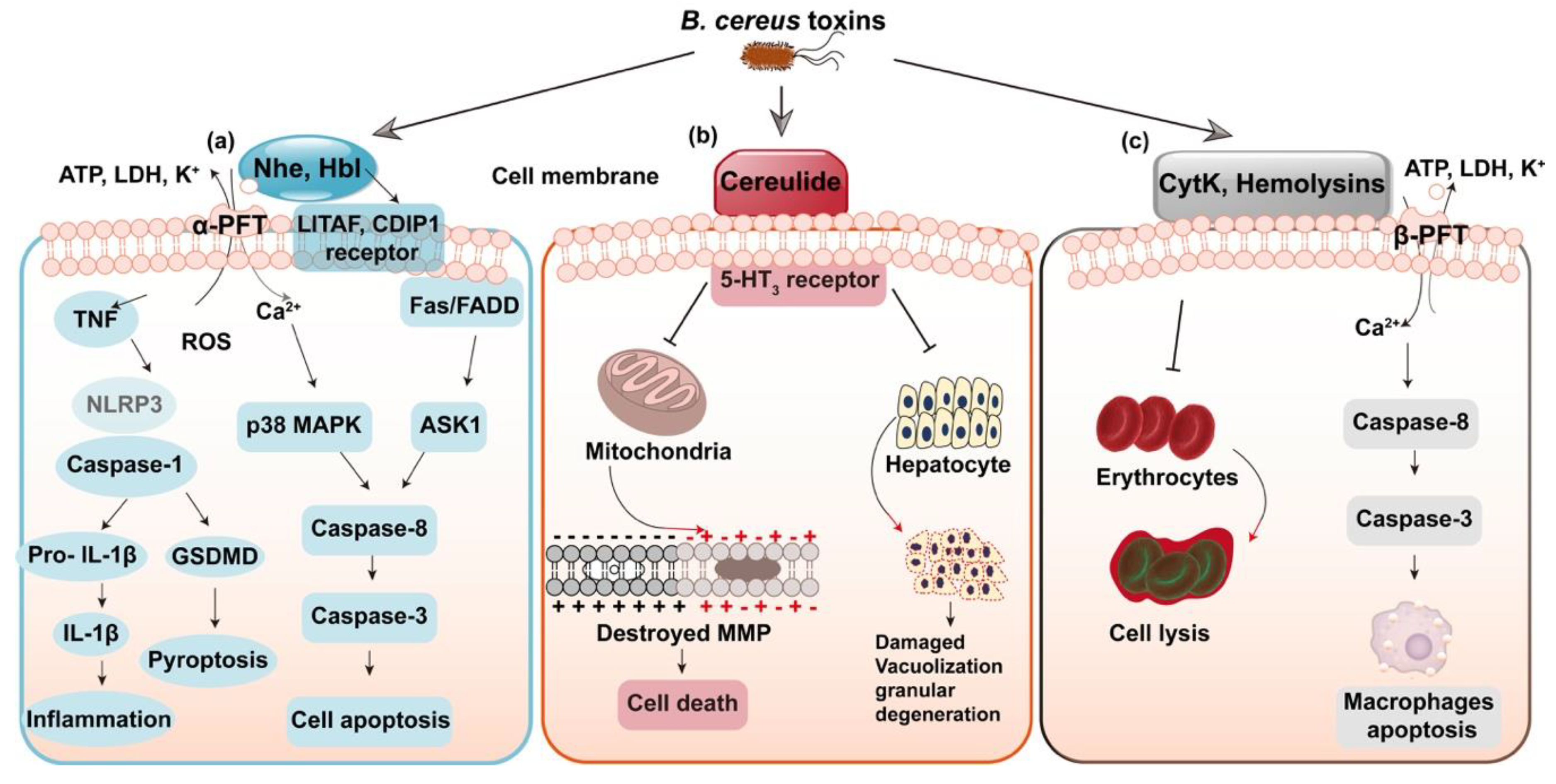
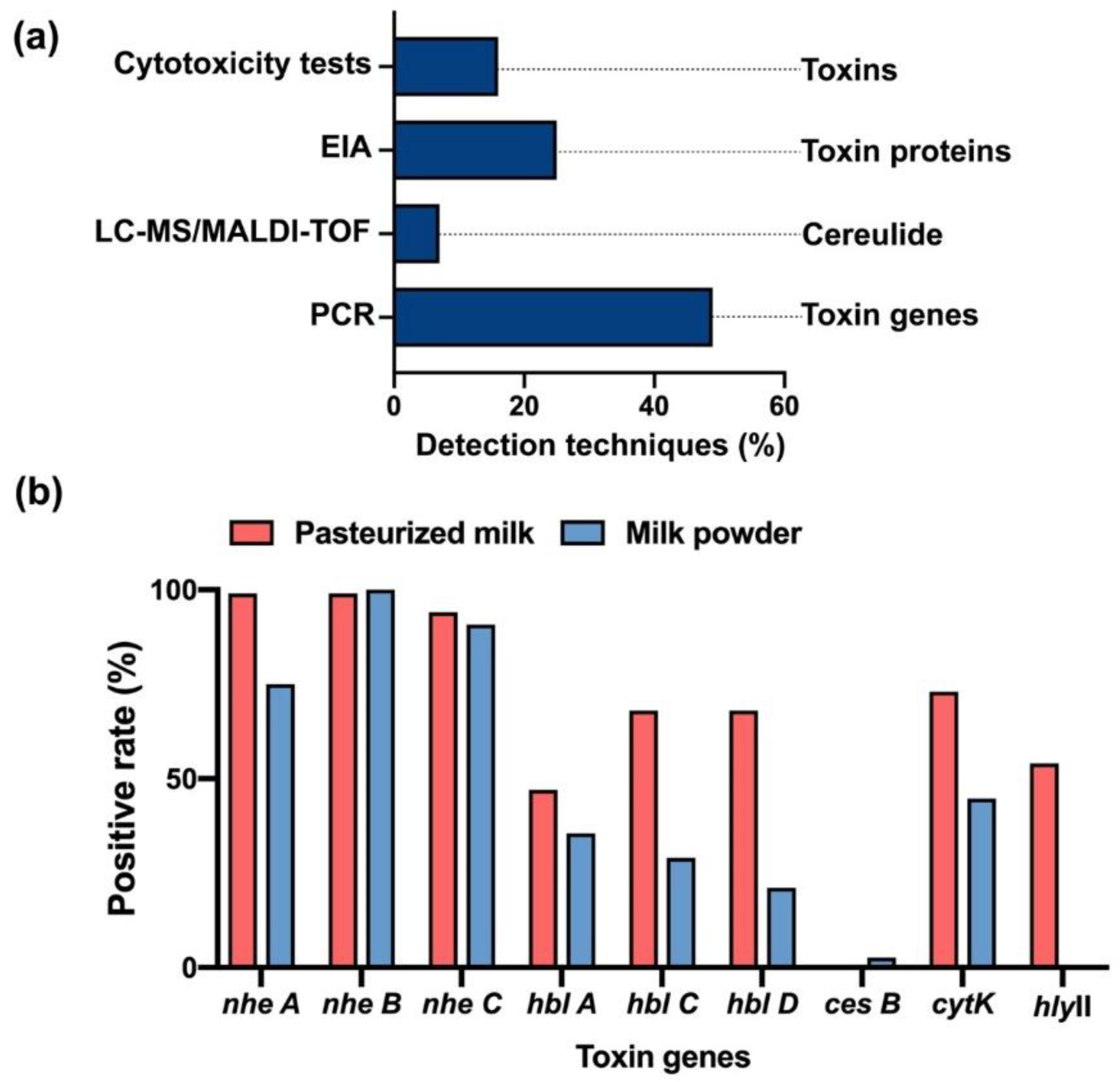
4. Virulence Factors of Bacillus cereus and Detection Techniques
4.1. Virulence Factors of Bacillus cereus
4.2. Detection of Bacillus cereus Isolates
4.3. Detection of Toxins Secreted from Bacillus cereus
4.4. Analysis of Bacillus Toxin Detections in Dairy Products in China
5. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Liu, X.; Liu, F.; Ding, S.; Shen, J.; Zhu, K. Sublethal levels of antibiotics promote bacterial persistence in epithelial cells. Adv. Sci. 2020. [Google Scholar] [CrossRef]
- Stenfors Arnesen, L.P.; Fagerlund, A.; Granum, P.E. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 2008, 32, 579–606. [Google Scholar] [CrossRef] [Green Version]
- Rigourd, V.; Barnier, J.P.; Ferroni, A.; Nicloux, M.; Hachem, T.; Magny, J.F.; Lapillonne, A.; Frange, P.; Nassif, X.; Bille, E. Recent actuality about Bacillus cereus and human milk bank: A new sensitive method for microbiological analysis of pasteurized milk. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Abdeen, E.E.; Hussien, H.; Hadad, G.A.E.; Mousa, W.S. Prevalence of virulence determinants among Bacillus cereus isolated from milk products with potential public health concern. Pak. J. Biol. Sci. 2020, 23, 206–212. [Google Scholar] [PubMed] [Green Version]
- Vidic, J.; Chaix, C.; Manzano, M.; Heyndrickx, M. Food sensing: detection of Bacillus cereus spores in dairy products. Biosensors 2020, 10, 15. [Google Scholar] [CrossRef] [Green Version]
- Fiedler, G.; Schneider, C.; Igbinosa, E.O.; Kabisch, J.; Brinks, E.; Becker, B.; Stoll, D.A.; Cho, G.S.; Huch, M.; Franz, C. Antibiotics resistance and toxin profiles of Bacillus cereus-group isolates from fresh vegetables from German retail markets. BMC Microbiol. 2019, 19, 250. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, J.; Feng, C.; Zhan, L.; Zhang, J.; Li, Y.; Yang, Y.; Chen, H.; Zhang, Z.; Zhang, Y.; et al. Quantitative prevalence, phenotypic and genotypic characteristics of Bacillus cereus isolated from retail infant foods in China. Foodborne Pathog. Dis. 2017, 14, 564–572. [Google Scholar] [CrossRef]
- Tunick, M.H.; Van Hekken, D.L. Dairy products and health: recent insights. J. Agric. Food Chem. 2015, 63, 9381–9388. [Google Scholar] [CrossRef]
- Godos, J.; Tieri, M.; Ghelfi, F.; Titta, L.; Marventano, S.; Lafranconi, A.; Gambera, A.; Alonzo, E.; Sciacca, S.; Buscemi, S.; et al. Dairy foods and health: An umbrella review of observational studies. Int. J. Food Sci. Nutr. 2020, 71, 138–151. [Google Scholar] [CrossRef]
- Romero-Alvarez, D.; Peterson, A.T.; Salzer, J.S.; Pittiglio, C.; Shadomy, S.; Traxler, R.; Vieira, A.R.; Bower, W.A.; Walke, H.; Campbell, L.P. Potential distributions of Bacillus anthracis and Bacillus cereus biovar anthracis causing anthrax in Africa. PLoS Negl. Trop. Dis. 2020, 14, e0008131. [Google Scholar] [CrossRef]
- Guo, R.; Tian, Y.; Zhang, H.; Guo, D.; Pei, X.; Wen, H.; Li, P.; Mehmood, K.; Yang, K.; Chang, Y.F.; et al. Biological characteristics and genetic evolutionary analysis of emerging pathogenic Bacillus cereus isolated from Pere David’s Deer (Elaphurus davidianus). Microb. Pathog. 2020. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Chen, J.; Fei, P.; Feng, H.; Wang, Y.; Ali, M.A.; Li, S.; Jing, H.; Yang, W. Prevalence, molecular characterization, and antibiotic susceptibility of Bacillus cereus isolated from dairy products in China. J. Dairy Sci. 2020, 103, 3994–4001. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Liu, X.; Dietrich, R.; Martlbauer, E.; Cao, J.; Ding, S.; Zhu, K. Characterization of Bacillus cereus isolates from local dairy farms in China. FEMS Microbiol. Lett. 2016, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, T.; Ding, Y.; Wu, Q.; Wang, J.; Zhang, J.; Yu, S.; Yu, P.; Liu, C.; Kong, L.; Feng, Z.; et al. Prevalence, virulence Genes, antimicrobial susceptibility, and genetic diversity of Bacillus cereus isolated from pasteurized milk in China. Front Microbiol 2018, 9, 533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.; Yu, P.; Wang, J.; Li, C.; Guo, H.; Liu, C.; Kong, L.; Yu, L.; Wu, S.; Lei, T.; et al. A study on prevalence and characterization of Bacillus cereus in ready-to-eat foods in China. Front. Microbiol. 2019, 10, 3043. [Google Scholar] [CrossRef] [Green Version]
- Granum, P.E. Spotlight on Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef] [Green Version]
- Dal Peraro, M.; van der Goot, F.G. Pore-forming toxins: Ancient, but never really out of fashion. Nat Rev. Microbiol. 2016, 14, 77–92. [Google Scholar] [CrossRef]
- Zhu, K.; Didier, A.; Dietrich, R.; Heilkenbrinker, U.; Waltenberger, E.; Jessberger, N.; Martlbauer, E.; Benz, R. Formation of small transmembrane pores: An intermediate stage on the way to Bacillus cereus non-hemolytic enterotoxin (Nhe) full pores in the absence of NheA. Biochem. Biophys. Res. Commun. 2016, 469, 613–618. [Google Scholar] [CrossRef]
- Fagerlund, A.; Lindback, T.; Storset, A.K.; Granum, P.E.; Hardy, S.P. Bacillus cereus Nhe is a pore-forming toxin with structural and functional properties similar to the ClyA (HlyE, SheA) family of haemolysins, able to induce osmotic lysis in epithelia. Microbiology 2008, 154, 693–704. [Google Scholar] [CrossRef] [Green Version]
- Jessberger, N.; Dietrich, R.; Schwemmer, S.; Tausch, F.; Schwenk, V.; Didier, A.; Martlbauer, E. Binding to the target cell surface is the crucial step in pore formation of hemolysin BL from Bacillus cereus. Toxins 2019, 11, 281. [Google Scholar] [CrossRef] [Green Version]
- Fagerlund, A.; Lindback, T.; Granum, P.E. Bacillus cereus cytotoxins Hbl, Nhe and CytK are secreted via the Sec translocation pathway. BMC Microbiol. 2010, 10, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehling-Schulz, M.; Svensson, B.; Guinebretiere, M.H.; Lindback, T.; Andersson, M.; Schulz, A.; Fricker, M.; Christiansson, A.; Granum, P.E.; Martlbauer, E.; et al. Emetic toxin formation of Bacillus cereus is restricted to a single evolutionary lineage of closely related strains. Microbiology 2005, 151, 183–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucking, G.; Frenzel, E.; Rutschle, A.; Marxen, S.; Stark, T.D.; Hofmann, T.; Scherer, S.; Ehling-Schulz, M. Ces locus embedded proteins control the non-ribosomal synthesis of the cereulide toxin in emetic Bacillus cereus on multiple levels. Front. Microbiol. 2015, 6, 1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramarao, N.; Sanchis, V. The pore-forming haemolysins of Bacillus cereus: A review. Toxins 2013, 5, 1119–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madegowda, M.; Eswaramoorthy, S.; Burley, S.K.; Swaminathan, S. X-ray crystal structure of the B component of hemolysin BL from Bacillus cereus. Proteins 2008, 71, 534–540. [Google Scholar] [CrossRef] [Green Version]
- Los, F.C.; Randis, T.M.; Aroian, R.V.; Ratner, A.J. Role of pore-forming toxins in bacterial infectious diseases. Microbiol. Mol. Biol. Rev. 2013, 77, 173–207. [Google Scholar] [CrossRef] [Green Version]
- Bischofberger, M.; Iacovache, I.; van der Goot, F.G. Pathogenic pore-forming proteins: function and host response. Cell Host Microbe 2012, 12, 266–275. [Google Scholar] [CrossRef] [Green Version]
- Alves, G.G.; Machado de Avila, R.A.; Chavez-Olortegui, C.D.; Lobato, F.C. Clostridium perfringens epsilon toxin: the third most potent bacterial toxin known. Anaerobe 2014, 30, 102–107. [Google Scholar] [CrossRef]
- Lindback, T.; Fagerlund, A.; Rodland, M.S.; Granum, P.E. Characterization of the Bacillus cereus Nhe enterotoxin. Microbiology 2004, 150, 3959–3967. [Google Scholar] [CrossRef] [Green Version]
- Heilkenbrinker, U.; Dietrich, R.; Didier, A.; Zhu, K.; Lindback, T.; Granum, P.E.; Martlbauer, E. Complex formation between NheB and NheC is necessary to induce cytotoxic activity by the three-component Bacillus cereus Nhe enterotoxin. PLoS ONE 2013, 8, e63104. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zuo, Z.; Sastalla, I.; Liu, C.; Jang, J.Y.; Sekine, Y.; Li, Y.; Pirooznia, M.; Leppla, S.H.; Finkel, T.; et al. Sequential CRISPR-Based Screens Identify LITAF and CDIP1 as the Bacillus cereus Hemolysin BL Toxin Host Receptors. Cell Host Microbe 2020. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ding, S.; Shi, P.; Dietrich, R.; Martlbauer, E.; Zhu, K. Non-hemolytic enterotoxin of Bacillus cereus induces apoptosis in Vero cells. Cell. Microbiol. 2017, 19. [Google Scholar] [CrossRef] [PubMed]
- Fox, D.; Mathur, A.; Xue, Y.; Liu, Y.; Tan, W.H.; Feng, S.; Pandey, A.; Ngo, C.; Hayward, J.A.; Atmosukarto, I.I.; et al. Bacillus cereus non-haemolytic enterotoxin activates the NLRP3 inflammasome. Nat. Commun. 2020, 11, 760. [Google Scholar] [PubMed]
- Doll, V.M.; Ehling-Schulz, M.; Vogelmann, R. Concerted action of sphingomyelinase and non-hemolytic enterotoxin in pathogenic Bacillus cereus. PLoS ONE 2013, 8, e61404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, M.A.; Hakulinen, P.; Honkalampi-Hamalainen, U.; Hoornstra, D.; Lhuguenot, J.C.; Maki-Paakkanen, J.; Savolainen, M.; Severin, I.; Stammati, A.L.; Turco, L.; et al. Toxicological profile of cereulide, the Bacillus cereus emetic toxin, in functional assays with human, animal and bacterial cells. Toxicon 2007, 49, 351–367. [Google Scholar] [CrossRef]
- Paananen, A.; Jarvinen, K.; Sareneva, T.; Salkinoja-Salonen, M.S.; Timonen, T.; Holtta, E. Valinomycin-induced apoptosis of human NK cells is predominantly caspase independent. Toxicology 2005, 212, 37–45. [Google Scholar] [CrossRef]
- Vangoitsenhoven, R.; Rondas, D.; Crevecoeur, I.; D’Hertog, W.; Baatsen, P.; Masini, M.; Andjelkovic, M.; Van Loco, J.; Matthys, C.; Mathieu, C.; et al. Foodborne cereulide causes beta-cell dysfunction and apoptosis. PLoS ONE 2014, 9, e104866. [Google Scholar] [CrossRef]
- Bauer, T.; Sipos, W.; Stark, T.D.; Kaser, T.; Knecht, C.; Brunthaler, R.; Saalmuller, A.; Hofmann, T.; Ehling-Schulz, M. First insights into within host translocation of the Bacillus cereus toxin cereulide using a porcine model. Front. Microbiol. 2018, 9, 2652. [Google Scholar] [CrossRef] [Green Version]
- Tran, S.-L.; Guillemet, E.; Ngo-Camus, M.; Clybouw, C.; Puhar, A.; Moris, A.; Gohar, M.; Lereclus, D.; Ramarao, N. Haemolysin II is a Bacillus cereus virulence factor that induces apoptosis of macrophages. Cell. Microbiol. 2011, 13, 92–108. [Google Scholar] [CrossRef]
- Jessberger, N.; Kranzler, M.; Da Riol, C.; Schwenk, V.; Buchacher, T.; Dietrich, R.; Ehling-Schulz, M.; Martlbauer, E. Assessing the toxic potential of enteropathogenic Bacillus cereus. Food Microbiol. 2019, 84, 103276. [Google Scholar] [CrossRef]
- Berthold-Pluta, A.; Pluta, A.; Garbowska, M. The effect of selected factors on the survival of Bacillus cereus in the human gastrointestinal tract. Microb. Pathog. 2015, 82, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, Y.; Okuno, T.; Minamiguchi, H.; Hodohara, K.; Mikamo, H.; Andoh, A. Survival of a case of Bacillus cereus meningitis with brain abscess presenting as immune reconstitution syndrome after febrile neutropenia—A case report and literature review. BMC Infect. Dis. 2020, 20, 15. [Google Scholar] [CrossRef] [PubMed]
- Meredith, F.T.; Fowler, V.G.; Gautier, M.; Corey, G.R.; Reller, L.B. Bacillus cereus necrotizing cellulitis mimicking clostridial myonecrosis: case report and review of the literature. Scand. J. Infect. Dis. 1997, 29, 528–529. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Liu, Y.; Wang, R.; Chen, T.; Yang, Z.; Deng, Y.; Zhao, Z.; Hu, X.; Chen, X.; Wei, W.; et al. Treatment of Bacillus cereus endophthalmitis with endoscopy-assisted vitrectomy. Medicine 2017, 96, e8701. [Google Scholar] [CrossRef] [PubMed]
- David, D.B.; Kirkby, G.R.; Noble, B.A. Bacillus cereus endophthalmitis. Br. J. Ophthalmol. 1994, 78, 577–580. [Google Scholar] [CrossRef] [Green Version]
- Ishida, R.; Ueda, K.; Kitano, T.; Yamamoto, T.; Mizutani, Y.; Tsutsumi, Y.; Imoto, K.; Yamamori, Y. Fatal community-acquired Bacillus cereus pneumonia in an immunocompetent adult man: A case report. BMC Infect. Dis. 2019, 19, 197. [Google Scholar] [CrossRef]
- Soudet, S.; Becquart, C.; Dezoteux, F.; Faure, K.; Staumont-Salle, D.; Delaporte, E. Bacillus cereus endocarditis and a probable cutaneous gateway. Ann. Dermatol. Venereol. 2017, 144, 45–48. [Google Scholar] [CrossRef]
- Avcu, G.; Sahbudak Bal, Z.; Cavusoglu, C.; Vardar, F. Osteomyelitis caused by Bacillus calmette-guerin vaccination in a healthy toddler. J. Trop. Pediatr. 2020, 66, 103–105. [Google Scholar] [CrossRef]
- Schaefer, G.; Campbell, W.; Jenks, J.; Beesley, C.; Katsivas, T.; Hoffmaster, A.; Mehta, S.R.; Reed, S. Persistent Bacillus cereus bacteremia in 3 persons who inject drugs, San Diego, California, USA. Emerg. Infect. Dis. 2016, 22, 1621–1623. [Google Scholar] [CrossRef] [Green Version]
- Ballard, O.; Morrow, A.L. Human milk composition: nutrients and bioactive factors. Pediatr. Clin. North Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [Green Version]
- Schroeter, H.; Holt, R.R.; Orozco, T.J.; Schmitz, H.H.; Keen, C.L. Nutrition: Milk and absorption of dietary flavanols. Nature 2003, 426, 787–788. [Google Scholar] [CrossRef] [PubMed]
- Rosa, D.D.; Dias, M.M.S.; Grzeskowiak, L.M.; Reis, S.A.; Conceicao, L.L.; Peluzio, M. Milk kefir: nutritional, microbiological and health benefits. Nutr. Res. Rev. 2017, 30, 82–96. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Yang, X.; Xia, J.; Zhao, L.; Yang, Y. Consumption of meat and dairy products in China: A review. Proc. Nutr. Soc. 2016, 75, 385–391. [Google Scholar] [CrossRef] [Green Version]
- Lagrange, V.; Whitsett, D.; Burris, C. Global market for dairy proteins. J. Food. Sci. 2015, 80, A16–A22. [Google Scholar] [CrossRef] [PubMed]
- Bian, S.; Hu, J.; Zhang, K.; Wang, Y.; Yu, M.; Ma, J. Dairy product consumption and risk of hip fracture: A systematic review and meta-analysis. BMC Public Health 2018, 18, 165. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Sun, Y.; Chen, J.; Tian, J. The quality of overviews on milk and dairy product consumption and cardiovascular diseases and milk and dairy product consumption and risk of mortality can be improved. Adv. Nutr. 2020, 11, 464. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Li, Y.; Lv, J.; Liu, X.; Gao, P.; Zhen, G.; Zhang, W.; Wu, D.; Jing, H.; Li, Y.; et al. A foodborne outbreak of gastroenteritis caused by Norovirus and Bacillus cereus at a university in the Shunyi District of Beijing, China 2018: A retrospective cohort study. BMC Infect. Dis. 2019, 19, 910. [Google Scholar] [CrossRef] [Green Version]
- Thirkell, C.E.; Sloan-Gardner, T.S.; Kacmarek, M.C.; Polkinghorne, B. An outbreak of Bacillus cereus toxin-mediated emetic and diarrhoeal syndromes at a restaurant in Canberra, Australia 2018. Commun. Dis. Intell. (2018) 2019, 43. [Google Scholar] [CrossRef]
- Chai, S.J.; Gu, W.; O’Connor, K.A.; Richardson, L.C.; Tauxe, R.V. Incubation periods of enteric illnesses in foodborne outbreaks, United States, 1998–2013. Epidemiol. Infect. 2019, 147, e285. [Google Scholar] [CrossRef] [Green Version]
- Ceuppens, S.; Rajkovic, A.; Heyndrickx, M.; Tsilia, V.; Van De Wiele, T.; Boon, N.; Uyttendaele, M. Regulation of toxin production by Bacillus cereus and its food safety implications. Crit. Rev. Microbiol. 2011, 37, 188–213. [Google Scholar] [CrossRef]
- Ombui, J.N.; Nduhiu, J.G. Prevalence of enterotoxigenic Bacillus cereus and its enterotoxins in milk and milk products in and around Nairobi. East Afr. Med. J. 2005, 82, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.A.M.; Silva, H.O.; Aguilar, C.E.G.; Rochetti, A.L.; Pascoe, B.; Méric, G.; Mourkas, E.; Hitchings, M.D.; Mathias, L.A.; de Azevedo Ruiz, V.L.; et al. Comparative genomic survey of Bacillus cereus sensu stricto isolates from the dairy production chain in Brazil. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef] [PubMed]
- Fasolato, L.; Cardazzo, B.; Carraro, L.; Fontana, F.; Novelli, E.; Balzan, S. Edible processed insects from e-commerce: Food safety with a focus on the Bacillus cereus group. Food Microbiol. 2018, 76, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Boor, K.J.; Wiedmann, M.; Murphy, S.; Alcaine, S. A 100-year review: microbiology and safety of milk handling. J. Dairy Sci. 2017, 100, 9933–9951. [Google Scholar] [CrossRef] [Green Version]
- Becker, H.; Schaller, G.; von Wiese, W.; Terplan, G. Bacillus cereus in infant foods and dried milk products. Int. J. Food Microbiol. 1994, 23, 1–15. [Google Scholar] [CrossRef]
- Zhou, G.; Liu, H.; He, J.; Yuan, Y.; Yuan, Z. The occurrence of Bacillus cereus, B. thuringiensis and B. mycoides in Chinese pasteurized full fat milk. Int. J. Food Microbiol. 2008, 121, 195–200. [Google Scholar] [CrossRef]
- Yusuf, U.; Kotwal, S.K.; Gupta, S.; Ahmed, T. Identification and antibiogram pattern of Bacillus cereus from the milk and milk products in and around Jammu region. Vet. World 2018, 11, 186–191. [Google Scholar] [CrossRef]
- Pedonese, F.; Verani, G.; Torracca, B.; Turchi, B.; Felicioli, A.; Nuvoloni, R. Effect of an Italian propolis on the growth of Listeria monocytogenes, Staphylococcus aureus and Bacillus cereus in milk and whey cheese. Ital. J. Food Saf. 2019, 8, 8036. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Flint, S.H.; Palmer, J.S. Bacillus cereus spores and toxins—The potential role of biofilms. Food Microbiol. 2020, 90, 103493. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, Y.; Liu, X.; Xia, X.; Ding, S.; Zhu, K. Evaluation of the toxicity and toxicokinetics of cereulide from an emetic Bacillus cereus strain of milk origin. Toxins 2016, 8, 156. [Google Scholar] [CrossRef] [Green Version]
- Sankararaman, S.; Velayuthan, S. Bacillus cereus. Pediatr. Rev. 2013, 34, 196–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christiansson, A.; Bertilsson, J.; Svensson, B. Bacillus cereus spores in raw milk: factors affecting the contamination of milk during the grazing period. J. Dairy Sci. 1999, 82, 305–314. [Google Scholar] [CrossRef]
- Owusu-Kwarteng, J.; Wuni, A.; Akabanda, F.; Tano-Debrah, K.; Jespersen, L. Prevalence, virulence factor genes and antibiotic resistance of Bacillus cereus sensu lato isolated from dairy farms and traditional dairy products. BMC Microbiol. 2017, 17, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanson, M.L.; Wendorff, W.L.; Houck, K.B. Effect of heat treatment of milk on activation of Bacillus spores. J. Food Prot. 2005, 68, 1484–1486. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Li, J.; Xi, X.; Xu, H.; Wuri, L.; Bian, Y.; Yu, Z.; Ren, M.; Duo, L.; Sun, Y.; et al. Evaluation of Bacterial contamination in goat milk powder using PacBio single molecule real-time sequencing and droplet digital PCR. J. Food Prot. 2018, 81, 1791–1799. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Wang, W.; Han, J.; Zhang, M. Investigation of Bacillus cereus contamination in infant formula food and cereals in Liaoning Province during 2016–2017. Chin. J. Health Lab. Tech. 2019, 29, 1525–1527. [Google Scholar]
- Fei, P.; Yuan, X.; Zhao, S.; Yang, T.; Xiang, J.; Chen, X.; Zhou, L.; Ji, M. Prevalence and genetic diversity of Bacillus cereus isolated from raw milk and cattle farm environments. Curr. Microbiol. 2019, 76, 1355–1360. [Google Scholar] [CrossRef]
- Zhuang, K.; Li, H.; Zhang, Z.; Wu, S.; Zhang, Y.; Fox, E.M.; Man, C.; Jiang, Y. Typing and evaluating heat resistance of Bacillus cereus sensu stricto isolated from the processing environment of powdered infant formula. J. Dairy Sci. 2019, 102, 7781–7793. [Google Scholar] [CrossRef]
- Jackson, E.E.; Erten, E.S.; Maddi, N.; Graham, T.E.; Larkin, J.W.; Blodgett, R.J.; Schlesser, J.E.; Reddy, R.M. Detection and enumeration of four foodborne pathogens in raw commingled silo milk in the United States. J. Food Prot. 2012, 75, 1382–1393. [Google Scholar] [CrossRef]
- Zhou, G.; Zheng, D.; Dou, L.; Cai, Q.; Yuan, Z. Occurrence of psychrotolerant Bacillus cereus group strains in ice creams. Int. J. Food Microbiol. 2010, 137, 143–146. [Google Scholar] [CrossRef]
- Xie, A.; Wu, K.; Li, Y.; Hong, C.; Hu, Y. Contamination survey and toxin genes distribution analysis of Bacillus cereus in milk powder in Wenzhou. Chin. J. Health Lab. Tech. 2018, 28, 2930–2932. [Google Scholar]
- Yang, Y.; Yu, X.; Zhan, L.; Chen, J.; Zhang, Y.; Zhang, J.; Chen, H.; Zhang, Z.; Zhang, Y.; Lu, Y. Multilocus sequence type profiles of Bacillus cereus isolates from infant formula in China. Food Microbiol. 2017, 62, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Yan, X.; Gan, X.; Li, Z.; Yu, D.; Zhao, X.; Li, F. Survey on contamination of Bacillus cereus and its virulence gene profiles isolated from retail infant formula in China. Chin. J. Food Hyg. 2015, 27, 286–291. [Google Scholar]
- Sun, T.; Wang, W.; Han, J.; Li, X.; Zhang, M.; Liu, H.; Wang, Y.; Zhang, M. Detection of virulence genes of Bacillus cereus in infant formula foods and cereal-based complementary foods in Liaoning in 2016. Chin. J. Microecol. 2018, 30, 791–794. [Google Scholar]
- Tang, X.; Yang, Z.; Fan, L.; Guo, Z. Investigation and assessment of Bacillus cereus contamination in infants and young children foods and ready-to-use foods in Yunnan from 2012 to 2016. J. Food Saf. Qual. 2017, 8, 3785–3789. [Google Scholar]
- Xie, C.; Wei, M.; Wang, H.; Jiang, Y.; Yang, X.; Hou, M. Investigation on hygiene and safety of school periphery food and infants food in Kunming in 2016. J. Food Saf. Qual. 2017, 8, 3903–3906. [Google Scholar]
- Sun, J.; Hu, X.; Lan, G.; Dong, K. Investigation on microbial contamination of infant formula powder during production process. Chin. J. Food Hyg. 2017, 29, 474–477. [Google Scholar]
- Li, Y.; Pei, X.; Yang, D.; Li, N. Occurrence of Bacillus cereus in infants and young children foods in 8 provinces in China. J. Food Res. 2014, 43, 435–438. [Google Scholar]
- Song, X.; Huang, X.; Xu, H.; Zhang, C.; Chen, S.; Liu, F.; Guan, S.; Zhang, S.; Zhu, K.; Wu, C. The prevalence of pathogens causing bovine mastitis and their associated risk factors in 15 large dairy farms in China: An observational study. Vet. Microbiol. 2020, 247. [Google Scholar] [CrossRef]
- Cui, Y.; Martlbauer, E.; Dietrich, R.; Luo, H.; Ding, S.; Zhu, K. Multifaceted toxin profile, an approach toward a better understanding of probiotic Bacillus cereus. Crit. Rev. Toxicol. 2019, 49, 342–356. [Google Scholar] [CrossRef]
- Lyu, Y.; Ye, L.; Xu, J.; Yang, X.; Chen, W.; Yu, H. Recent research progress with phospholipase C from Bacillus cereus. Biotechnol. Lett. 2016, 38, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Gohar, M.; Faegri, K.; Perchat, S.; Ravnum, S.; Okstad, O.A.; Gominet, M.; Kolsto, A.B.; Lereclus, D. The PlcR virulence regulon of Bacillus cereus. PLoS ONE 2008, 3, e2793. [Google Scholar] [CrossRef]
- Lucking, G.; Dommel, M.K.; Scherer, S.; Fouet, A.; Ehling-Schulz, M. Cereulide synthesis in emetic Bacillus cereus is controlled by the transition state regulator AbrB, but not by the virulence regulator PlcR. Microbiology 2009, 155, 922–931. [Google Scholar] [CrossRef] [Green Version]
- Naka, T.; Takaki, Y.; Hattori, Y.; Takenaka, H.; Ohta, Y.; Kirihata, M.; Tanimori, S. Chemical structure of hydrolysates of cereulide and their time course profile. Bioorg. Med. Chem. Lett. 2020, 30. [Google Scholar] [CrossRef] [PubMed]
- Ehling-Schulz, M.; Vukov, N.; Schulz, A.; Shaheen, R.; Andersson, M.; Martlbauer, E.; Scherer, S. Identification and partial characterization of the nonribosomal peptide synthetase gene responsible for cereulide production in emetic Bacillus cereus. Appl. Environ. Microbiol. 2005, 71, 105–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ISO 7932:2004. Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Presumptive Bacillus Cereus—Colony-Count Technique at 30 Degrees C; ISO: Geneva, Switzerland, 2004. [Google Scholar]
- Yossa, N.; Arce, G.; Huang, M.J.; Yin, L.; Brown, E.; Hammack, T. Factors of detection of Bacillus cereus strains in eye cream. Int. J. Cosmet. Sci. 2017, 39, 179–187. [Google Scholar] [CrossRef]
- Magnusson, M.; Christiansson, A.; Svensson, B. Bacillus cereus spores during housing of dairy cows: factors affecting contamination of raw milk. J. Dairy Sci. 2007, 90, 2745–2754. [Google Scholar] [CrossRef]
- Shaheen, R.; Svensson, B.; Andersson, M.A.; Christiansson, A.; Salkinoja-Salonen, M. Persistence strategies of Bacillus cereus spores isolated from dairy silo tanks. Food Microbiol. 2010, 27, 347–355. [Google Scholar] [CrossRef]
- Ouertani, A.; Chaabouni, I.; Mosbah, A.; Long, J.; Barakat, M.; Mansuelle, P.; Mghirbi, O.; Najjari, A.; Ouzari, H.I.; Masmoudi, A.S.; et al. Two new secreted proteases generate a casein-derived antimicrobial peptide in Bacillus cereus food born isolate leading to bacterial competition in milk. Front. Microbiol. 2018, 9, 1148. [Google Scholar] [CrossRef]
- Lakshmi, B.K.M.; Muni Kumar, D.; Hemalatha, K.P.J. Purification and characterization of alkaline protease with novel properties from Bacillus cereus strain S8. J. Genet. Eng. Biotechnol. 2018, 16, 295–304. [Google Scholar] [CrossRef]
- Hakovirta, J.R.; Prezioso, S.; Hodge, D.; Pillai, S.P.; Weigel, L.M. Identification and analysis of informative single nucleotide polymorphisms in 16S rRNA gene sequences of the Bacillus cereus group. J. Clin. Microbiol. 2016, 54, 2749–2756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Du, J.; Lai, Q.; Zeng, R.; Ye, D.; Xu, J.; Shao, Z. Proposal of nine novel species of the Bacillus cereus group. Int. J. Syst. Evol. Microbiol. 2017, 67, 2499–2508. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Chelliah, R.; Park, B.J.; Park, J.H.; Forghani, F.; Park, Y.S.; Cho, M.S.; Park, D.S.; Oh, D.H. Molecular discrimination of Bacillus cereus group species in foods (lettuce, spinach, and kimbap) using quantitative real-time PCR targeting groEL and gyrB. Microb. Pathog. 2018, 115, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Heini, N.; Stephan, R.; Ehling-Schulz, M.; Johler, S. Characterization of Bacillus cereus group isolates from powdered food products. Int. J. Food Microbiol. 2018, 283, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kindle, P.; Etter, D.; Stephan, R.; Johler, S. Population structure and toxin gene profiles of Bacillus cereus sensu lato isolated from flour products. FEMS Microbiol. Lett. 2019, 366. [Google Scholar] [CrossRef] [PubMed]
- Antolinos, V.; Fernández, P.S.; Ros-Chumillas, M.; Periago, P.M.; Weiss, J. Development of a high-resolution melting-based approach for efficient differentiation among Bacillus cereus group isolates. Foodborne Pathog. Dis. 2012, 9, 777–785. [Google Scholar] [CrossRef]
- Zhang, J.; Di, B.; Shan, H.; Liu, J.; Zhou, Y.; Chen, H.; Hu, L.; Wu, X.; Bai, Z. Rapid detection of Bacillus cereus using cross-priming amplification. J. Food Prot. 2019, 82, 1744–1750. [Google Scholar] [CrossRef]
- Wehrle, E.; Moravek, M.; Dietrich, R.; Burk, C.; Didier, A.; Martlbauer, E. Comparison of multiplex PCR, enzyme immunoassay and cell culture methods for the detection of enterotoxinogenic Bacillus cereus. J. Microbiol. Methods 2009, 78, 265–270. [Google Scholar] [CrossRef]
- Oltuszak-Walczak, E.; Walczak, P. PCR detection of cytK gene in Bacillus cereus group strains isolated from food samples. J. Microbiol. Methods 2013, 95, 295–301. [Google Scholar] [CrossRef]
- Fagerlund, A.; Ween, O.; Lund, T.; Hardy, S.P.; Granum, P.E. Genetic and functional analysis of the cytK family of genes in Bacillus cereus. Microbiology 2004, 150, 2689–2697. [Google Scholar] [CrossRef] [Green Version]
- Yu, P.; Yu, S.; Wang, J.; Guo, H.; Zhang, Y.; Liao, X.; Zhang, J.; Wu, S.; Gu, Q.; Xue, L.; et al. Bacillus cereus isolated from vegetables in China: incidence, genetic diversity, virulence genes, and antimicrobial resistance. Front. Microbiol. 2019, 10, 948. [Google Scholar]
- Yu, S.; Yan, L.; Wu, X.; Li, F.; Wang, D.; Xu, H. Multiplex PCR coupled with propidium monoazide for the detection of viable Cronobacter sakazakii, Bacillus cereus, and Salmonella spp. in milk and milk products. J. Dairy Sci. 2017, 100, 7874–7882. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Xie, G.; Liang, T.; Yu, B.; Aguilar, Z.; Xu, H. Rapid and quantitative detection of viable emetic Bacillus cereus by PMA-qPCR assay in milk. Mol. Cell. Probes 2019, 47, 101437. [Google Scholar] [CrossRef] [PubMed]
- Baida, G.; Budarina, Z.I.; Kuzmin, N.P.; Solonin, A.S. Complete nucleotide sequence and molecular characterization of hemolysin II gene from Bacillus cereus. FEMS Microbiol. Lett. 1999, 180, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Ueda, S.; Yamaguchi, M.; Eguchi, K.; Iwase, M. Identification of cereulide-producing Bacillus cereus by nucleic acid chromatography and reverse transcription real-time PCR. Biocontrol. Sci. 2016, 21, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, R.; Fella, C.; Strich, S.; Martlbauer, E. Production and characterization of monoclonal antibodies against the hemolysin BL enterotoxin complex produced by Bacillus cereus. Appl. Environ. Microbiol. 1999, 65, 4470–4474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietrich, R.; Moravek, M.; Burk, C.; Granum, P.E.; Martlbauer, E. Production and characterization of antibodies against each of the three subunits of the Bacillus cereus nonhemolytic enterotoxin complex. Appl. Environ. Microbiol. 2005, 71, 8214–8220. [Google Scholar] [CrossRef] [Green Version]
- Didier, A.; Dietrich, R.; Gruber, S.; Bock, S.; Moravek, M.; Nakamura, T.; Lindback, T.; Granum, P.E.; Martlbauer, E. Monoclonal antibodies neutralize Bacillus cereus Nhe enterotoxin by inhibiting ordered binding of its three exoprotein components. Infect. Immun. 2012, 80, 832–838. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, N.; Nagai, S.; Fujita, A.; Ido, Y.; Kato, K.; Saito, A.; Moriya, Y.; Tomimatsu, Y.; Kaneta, N.; Tsujimoto, Y.; et al. Discrimination of psychrotolerant Bacillus cereus group based on MALDI-TOF MS analysis of ribosomal subunit proteins. Food Microbiol. 2020, 91, 103542. [Google Scholar] [CrossRef]
- Ducrest, P.J.; Pfammatter, S.; Stephan, D.; Vogel, G.; Thibault, P.; Schnyder, B. Rapid detection of Bacillus ionophore cereulide in food products. Sci. Rep. 2019, 9, 5814. [Google Scholar] [CrossRef] [Green Version]
- Ueda, S.; Nakajima, H.; Iwase, M.; Shinagawa, K.; Kuwabara, Y. LC-MS analysis of the emetic toxin, cereulide, produced by Bacillus cereus. Biocontrol. Sci. 2012, 17, 191–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, M.; Kawai, T.; Kitagawa, M.; Kumeda, Y. A new method for rapid and quantitative detection of the Bacillus cereus emetic toxin cereulide in food products by liquid chromatography-tandem mass spectrometry analysis. Food Microbiol. 2013, 34, 29–37. [Google Scholar] [CrossRef]
- Saxena, R.; Singh, R. MALDI-TOF MS and CD spectral analysis for identification and structure prediction of a purified, novel, organic solvent stable, fibrinolytic metalloprotease from Bacillus cereus B80. Biomed. Res. Int. 2015, 2015, 527015. [Google Scholar] [CrossRef] [PubMed]
- Stark, T.; Marxen, S.; Rutschle, A.; Lucking, G.; Scherer, S.; Ehling-Schulz, M.; Hofmann, T. Mass spectrometric profiling of Bacillus cereus strains and quantitation of the emetic toxin cereulide by means of stable isotope dilution analysis and HEp-2 bioassay. Anal. Bioanal. Chem. 2013, 405, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Shinagawa, K.; Ueno, Y.; Hu, D.; Ueda, S.; Sugii, S. Mouse lethal activity of a HEp-2 vacuolation factor, cereulide, produced by Bacillus cereus isolated from vomiting-type food poisoning. J. Vet. Med. Sci. 1996, 58, 1027–1029. [Google Scholar] [CrossRef] [Green Version]
- Jeßberger, N.; Dietrich, R.; Bock, S.; Didier, A.; Märtlbauer, E. Bacillus cereus enterotoxins act as major virulence factors and exhibit distinct cytotoxicity to different human cell lines. Toxicon 2014, 77, 49–57. [Google Scholar] [CrossRef]
- Tran, S.L.; Ramarao, N. Bacillus cereus immune escape: A journey within macrophages. FEMS Microbiol. Lett. 2013, 347, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, E.; He, J.; Li, W.; Wei, J. Distribution of virulence genes of Bacillus cereus in several specimens. Chin. J. Zoonoses 2019, 35, 683–687. [Google Scholar]
- Lin, Y.; Ren, F.; Zhao, L.; Guo, H. Genotypes and the persistence survival phenotypes of Bacillus cereus isolated from UHT milk processing lines. Food Control. 2017, 82, 48–56. [Google Scholar] [CrossRef]
- Sánchez Chica, J.; Correa, M.M.; Aceves-Diez, A.E.; Rasschaert, G.; Heyndrickx, M.; Castañeda-Sandoval, L.M. Genomic and toxigenic heterogeneity of Bacillus cereus sensu lato isolated from ready-to-eat foods and powdered milk in day care centers in Colombia. Foodborne Pathog. Dis. 2020, 17, 340–347. [Google Scholar] [CrossRef] [Green Version]
- Saleh-Lakha, S.; Leon-Velarde, C.G.; Chen, S.; Lee, S.; Shannon, K.; Fabri, M.; Downing, G.; Keown, B. A study to assess the numbers and prevalence of Bacillus cereus and its toxins in pasteurized fluid milk. J. Food Prot. 2017, 80, 1085–1089. [Google Scholar] [CrossRef] [PubMed]
- Glasset, B.; Herbin, S.; Guillier, L.; Cadel-Six, S.; Vignaud, M.L.; Grout, J.; Pairaud, S.; Michel, V.; Hennekinne, J.A.; Ramarao, N.; et al. Bacillus cereus-induced food-borne outbreaks in France, 2007 to 2014: epidemiology and genetic characterisation. Euro Surveill. 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Yibar, A.; Çetinkaya, F.; Soyutemiz, E.; Yaman, G. Prevalence, enterotoxin production and antibiotic resistance of Bacillus cereus isolated from milk and cheese Kafkas. Univ. Vet. Fak. Derg. 2017, 23, 635–642. [Google Scholar]
- Ramarao, N.; Tran, S.L.; Marin, M.; Vidic, J. Advanced methods for detection of Bacillus cereus and its pathogenic factors. Sensors 2020, 20, 2667. [Google Scholar] [CrossRef]
- Gao, S.; Ni, C.; Huang, W.; Hao, H.; Jiang, H.; Lv, Q.; Zheng, Y.; Liu, P.; Kong, D.; Jiang, Y. The interaction between flagellin and the glycosphingolipid Gb3 on host cells contributes to Bacillus cereus acute infection. Virulence 2020, 11, 769–780. [Google Scholar] [CrossRef] [PubMed]
- Ramm, F.; Dondapati, S.K.; Thoring, L.; Zemella, A.; Wustenhagen, D.A.; Frentzel, H.; Stech, M.; Kubick, S. Mammalian cell-free protein expression promotes the functional characterization of the tripartite non-hemolytic enterotoxin from Bacillus cereus. Sci. Rep. 2020, 10, 2887. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.K.; Kim, W.S.; Paik, H.D. Bacillus strains as human probiotics: characterization, safety, microbiome, and probiotic carrier. Food Sci. Biotechnol. 2019, 28, 1297–1305. [Google Scholar] [CrossRef]
- Duc le, H.; Hong, H.A.; Barbosa, T.M.; Henriques, A.O.; Cutting, S.M. Characterization of Bacillus probiotics available for human use. Appl. Environ. Microbiol. 2004, 70, 2161–2171. [Google Scholar] [CrossRef] [Green Version]
- Green, D.H.; Wakeley, P.R.; Page, A.; Barnes, A.; Baccigalupi, L.; Ricca, E.; Cutting, S.M. Characterization of two Bacillus probiotics. Appl. Environ. Microbiol. 1999, 65, 4288–4291. [Google Scholar] [CrossRef] [Green Version]
- Hong, H.A.; Duc le, H.; Cutting, S.M. The use of bacterial spore formers as probiotics. FEMS Microbiol. Rev. 2005, 29, 813–835. [Google Scholar] [CrossRef] [Green Version]
- Colombo, M.; Todorov, S.D.; Eller, M.; Nero, L.A. The potential use of probiotic and beneficial bacteria in the Brazilian dairy industry. J. Dairy Res. 2018, 85, 487–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harms, A.; Maisonneuve, E.; Gerdes, K. Mechanisms of bacterial persistence during stress and antibiotic exposure. Science 2016, 354. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cui, Y.; Peng, W.; Huang, X.; Ma, L.; Zheng, M.; Ding, S.; Zhu, K. Prevalence of pathogens harboring mobile antimicrobial resistance genes and virulence factors in retail beef and mutton. FEMS Microbiol. Lett. 2020. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Region | Year | No. of B. cereus Isolates/ No. of Samples | Detection of Toxin Genes (%) | Reference | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| nheA | nheB | nheC | hblA | hblC | hblD | cesB | cytK | HlyⅡ | |||||
| Raw milk | Beijing | 2013–2014 | 92/306 | 100 | 100 | 100 | 79 | 79 | 79 | ND | ND | ND | [13] |
| Raw milk | Northeast China | 2017–2018 | 56/350 | ND | ND | ND | ND | ND | ND | ND | ND | ND | [77] |
| Pasteurized milk | Major cities in China, including Beijing, Nanchang, Chengdu, Hefei, Wuhan, Shanghai, et al. | 2011–2016 | 70/258 | 99 | 99 | 94 | 47 | 68 | 68 | 5 | 73 | 54 | [14] |
| Pasteurized milk | Wuhan | 2006 | 26/54 | 71.7 | 62 | 71.7 | 37 | 66.3 | 71.7 | ND | ND | ND | [80] |
| Ice cream | Wuhan | 2006 | 24/40 | ND | ND | ND | ND | ND | ND | ND | ND | ND | [66] |
| Milk powder | Wenzhou | 2015–2016 | 76/400 | 75 | 100 | 90.8 | 35.5 | 29.0 | 21.1 | ND | 44.7 | ND | [81] |
| Infant formula | Chinese markets | 2012–2013 | 74/513 | ND | ND | ND | ND | ND | ND | ND | ND | ND | [82] |
| Infant formula | Chinese markets | 2015 | 57/135 | 87.7 | 87.7 | 49.1 | 24.6 | 22.8 | 17.5 | 3.5 | 22.8 | ND | [83] |
| Infant formula | Liaoning | 2016 | 22/176 | 90.9 | 72.7 | 100 | 0 | 59.1 | 54.5 | ND | 68.2 | ND | [84] |
| Infant formula | Chinese markets | 2013–2015 | 33/401 | ND | ND | ND | ND | ND | ND | ND | ND | ND | [7] |
| Infant formula | Liaoning | 2016–2017 | 70/166 | ND | ND | ND | ND | ND | ND | ND | ND | ND | [76] |
| Infant formula | Yunnan | 2012–2016 | 71/605 | ND | ND | ND | ND | ND | ND | ND | ND | ND | [85] |
| Infant formula | Kunming | 2016 | 5/126 | ND | ND | ND | ND | ND | ND | ND | ND | ND | [86] |
| Infant formula and processing facility | Gansu | 2013–2014 | 31/183 | ND | ND | ND | ND | ND | ND | ND | ND | ND | [87] |
| Infant formula | Heilongjiang, Hebei, Henan, Hubei, Hunan, Jiangsu, Jiangxi, Guangdong | 2012 | 115/817 | ND | ND | ND | ND | ND | ND | ND | ND | ND | [88] |
| Dairy products | Heilongjiang, Jilin, Hebei, Henan, Guizhou | 2018–2019 | 54/500 | 94.4 | 94.4 | 100 | 57.4 | 68.5 | 16.7 | 11.1 | 75.9 | 53.7 | [12] |
| Toxin Genes (%) | Origin | Source | Year | Reference | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| nheA | nheB | nheC | hblA | hblC | hblD | cesB | cytK | hlyⅡ | ||||
| nhe 100 | hbl 78.3 | 1.1 | -- | -- | Beijing, China | Dairy farms | 2013–2014 | [13] | ||||
| 90.9 | 72.7 | 100 | 0 | 59.1 | 54.5 | -- | 68.2 | -- | Liaoning, China | Milk powder | 2016 | [84] |
| 87.2 | 81.6 | 86.4 | 36 | 38.4 | 38.4 | 3.2 | 36.8 | -- | Hebei, Hainan, Yunnan province, et al., China | Milk powder | 2019 | [129] |
| 74.1 | 88.9 | 100 | 55.6 | 77.8 | 0 | 48.2 | 33.3 | -- | China | UHT milk processing line | 2014–2015 | [130] |
| 84.1 | 89.9 | 84.1 | 59.4 | 44.9 | 53.6 | 2.9 | 53.6 | -- | Brazil | Dairy production chain | 2016 | [62] |
| nhe 100 | hbl 29.5 | 0 | 24.1 | -- | Colombia | Ready-to-eat food and milk | 2013 | [131] | ||||
| 76.5 | -- | -- | -- | 41.2 | -- | 0 | 5.9 | -- | Canada | Pasteurized milk | 2014–2015 | [132] |
| 96 | 99 | 100 | 44 | 40 | 44 | -- | 42 | 23 | France | FBO | 2007–2014 | [133] |
| 6.3 | 2.1 | 4.2 | 11.5 | 10.4 | 16.7 | 9.4 | 75 | -- | Ghana | Dairy farm | 2015 | [73] |
| 60 | 60 | 60 | 13 | 13 | 113 | -- | 75 | -- | Turkey | Milk and cheese | 2013 | [134] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.-Y.; Hu, Q.; Xu, F.; Ding, S.-Y.; Zhu, K. Characterization of Bacillus cereus in Dairy Products in China. Toxins 2020, 12, 454. https://doi.org/10.3390/toxins12070454
Liu X-Y, Hu Q, Xu F, Ding S-Y, Zhu K. Characterization of Bacillus cereus in Dairy Products in China. Toxins. 2020; 12(7):454. https://doi.org/10.3390/toxins12070454
Chicago/Turabian StyleLiu, Xiao-Ye, Qiao Hu, Fei Xu, Shuang-Yang Ding, and Kui Zhu. 2020. "Characterization of Bacillus cereus in Dairy Products in China" Toxins 12, no. 7: 454. https://doi.org/10.3390/toxins12070454
APA StyleLiu, X.-Y., Hu, Q., Xu, F., Ding, S.-Y., & Zhu, K. (2020). Characterization of Bacillus cereus in Dairy Products in China. Toxins, 12(7), 454. https://doi.org/10.3390/toxins12070454